
Mucin 1 as a Molecular Target of a Novel Diisoquinoline Derivative Combined with Anti-MUC1 Antibody in AGS Gastric Cancer Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
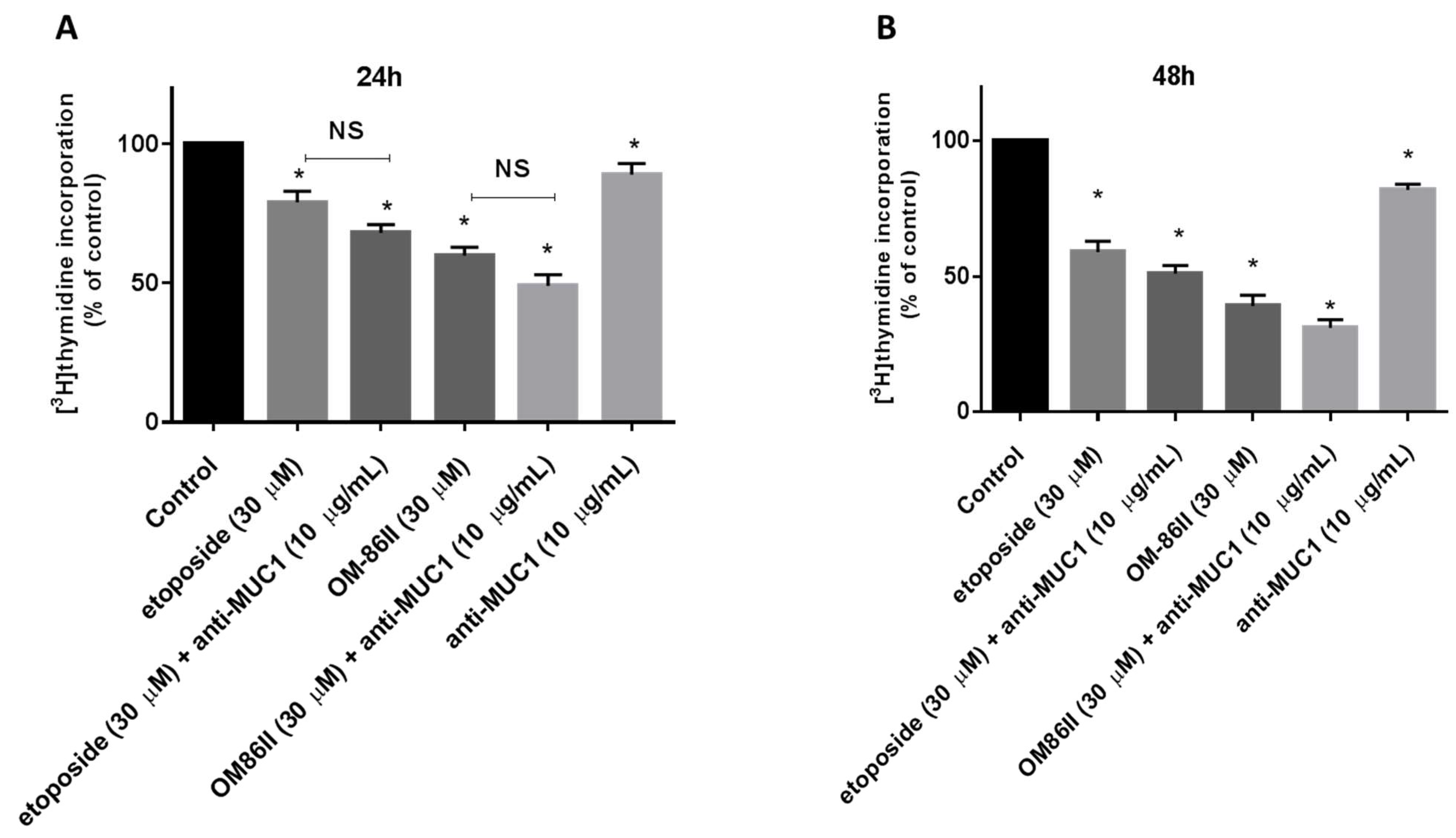
2.1. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Inhibits the Growth and Proliferation of Human Gastric Cancer Cells
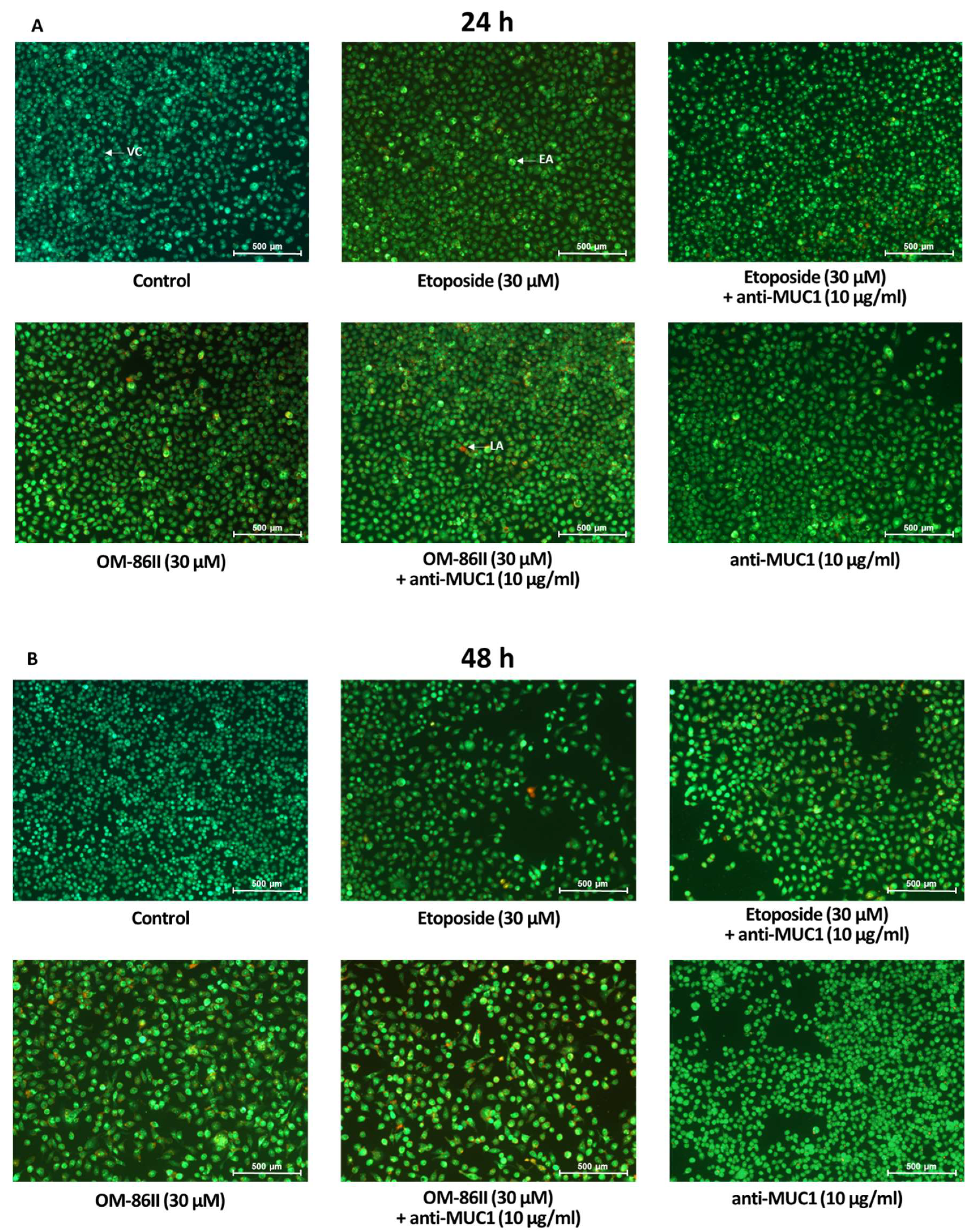
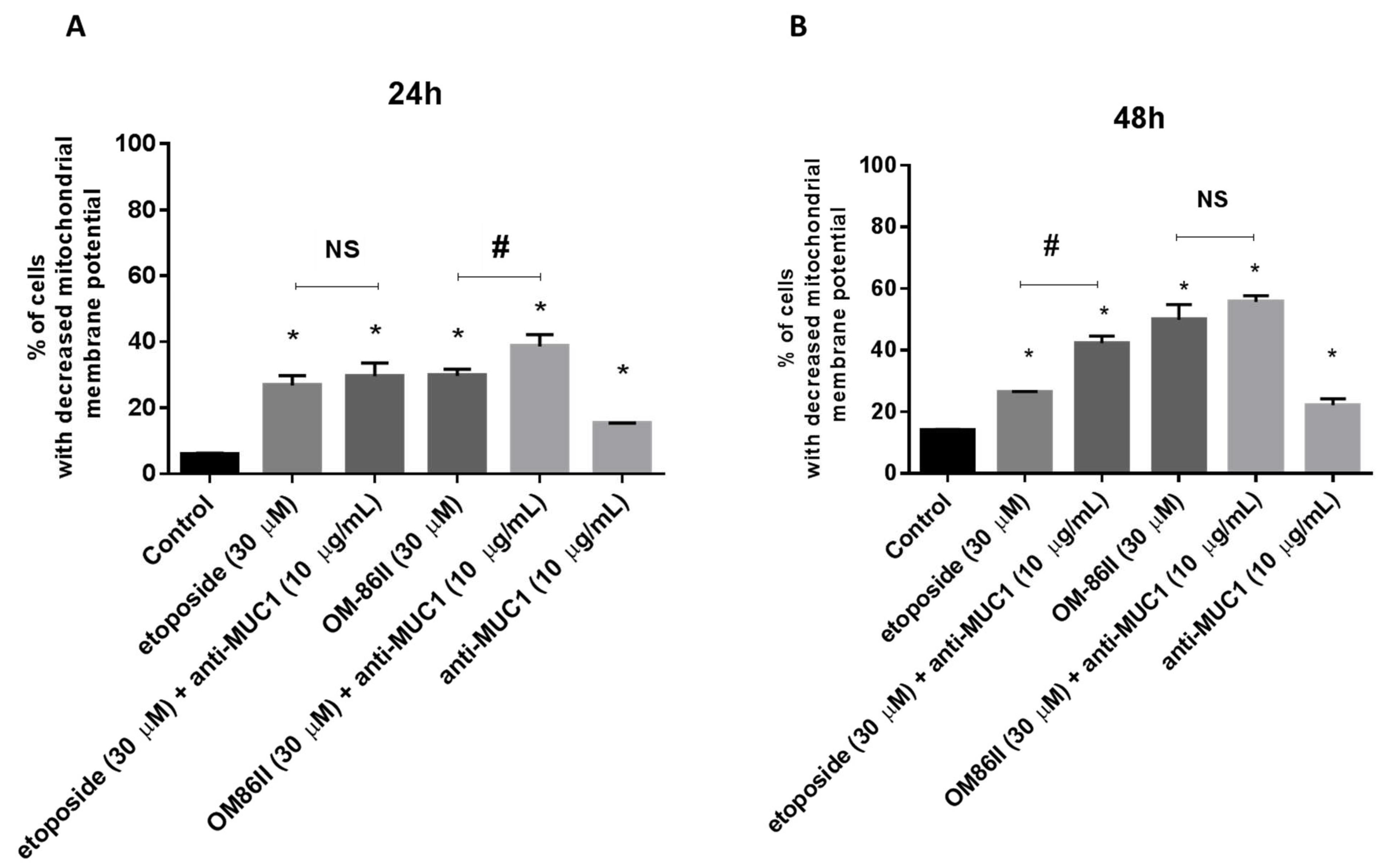
2.2. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Possesses Pro-Apoptotic Activity
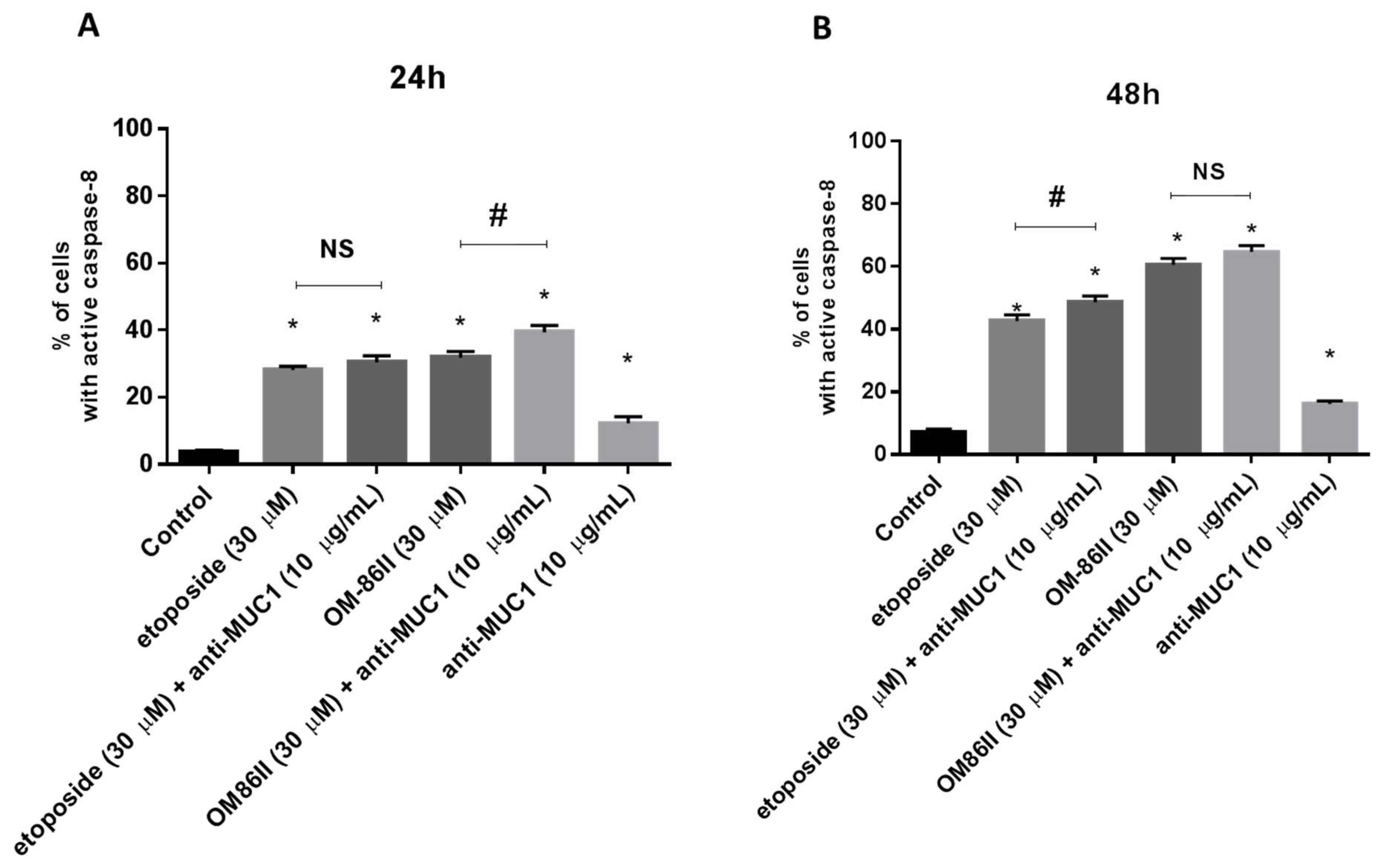
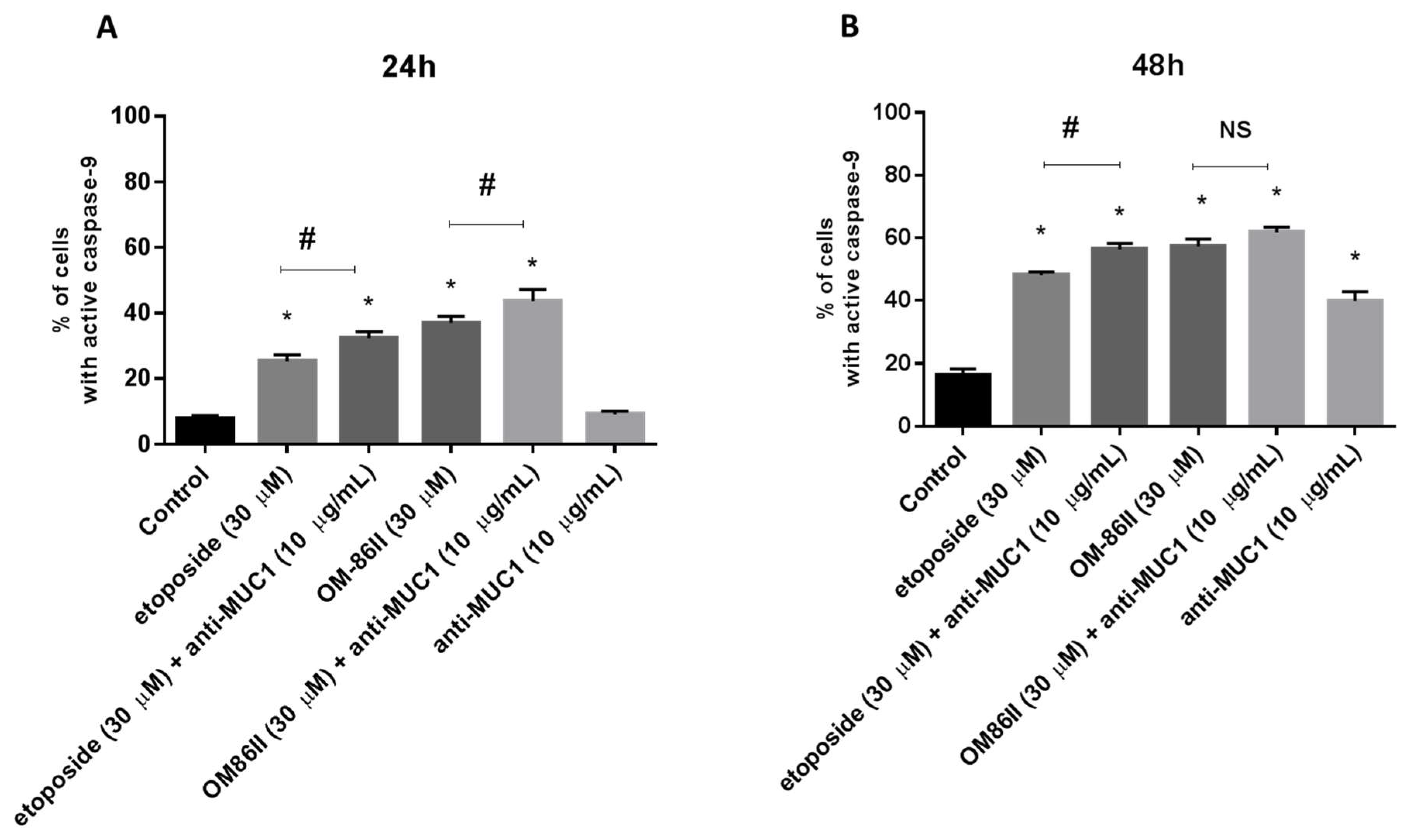
2.3. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Increases Caspase-8 and Caspase-9 Activity
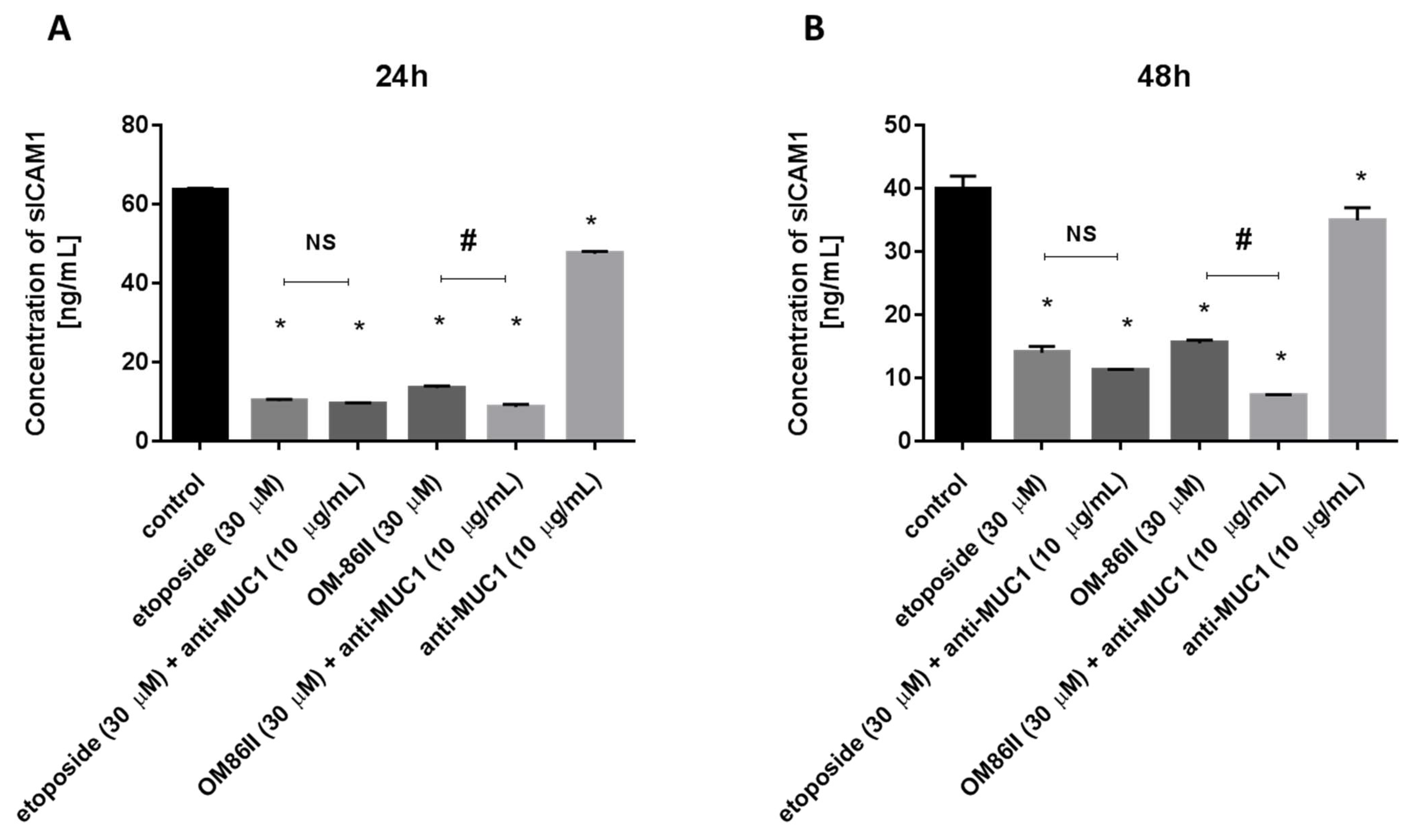
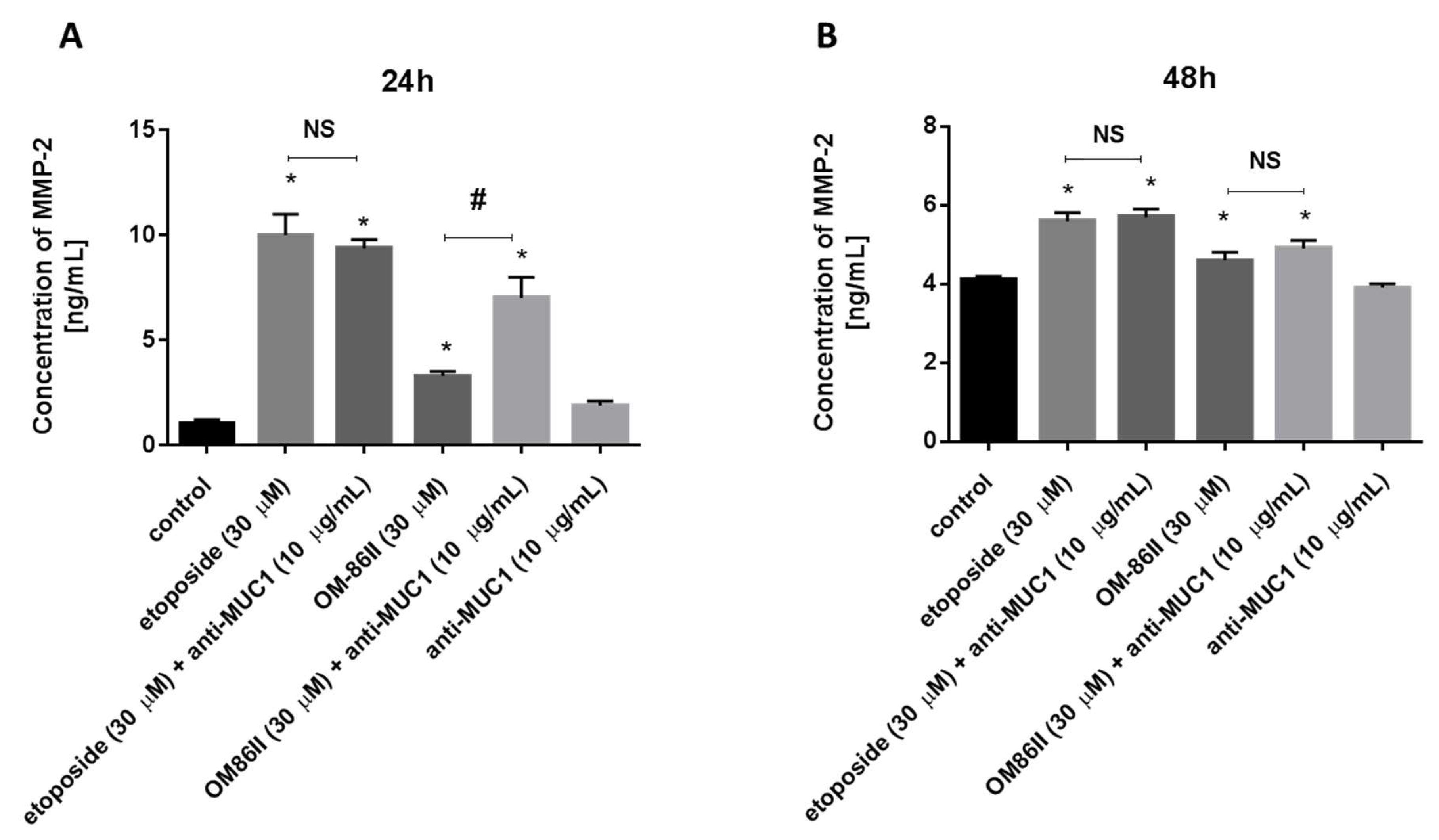
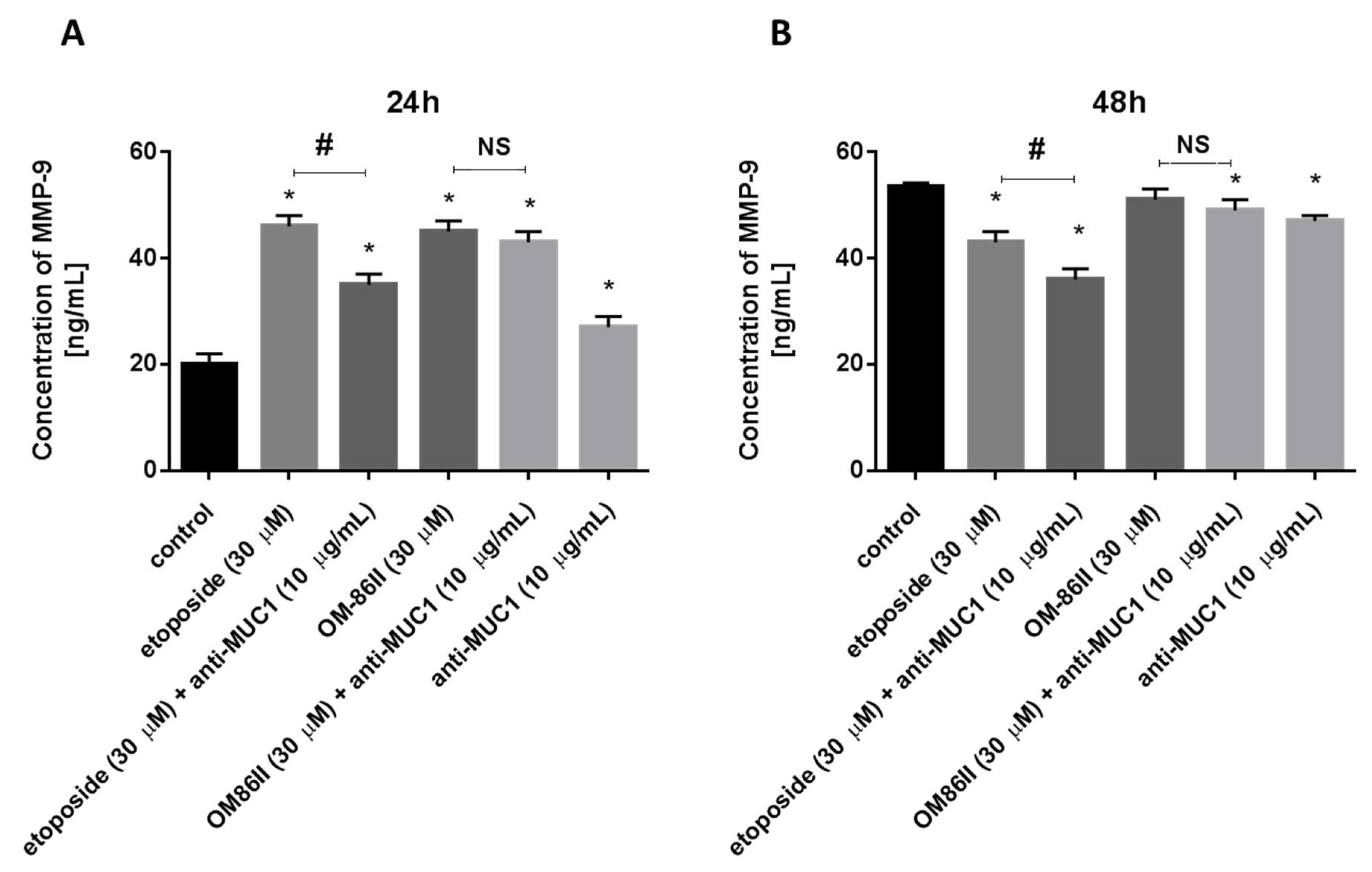
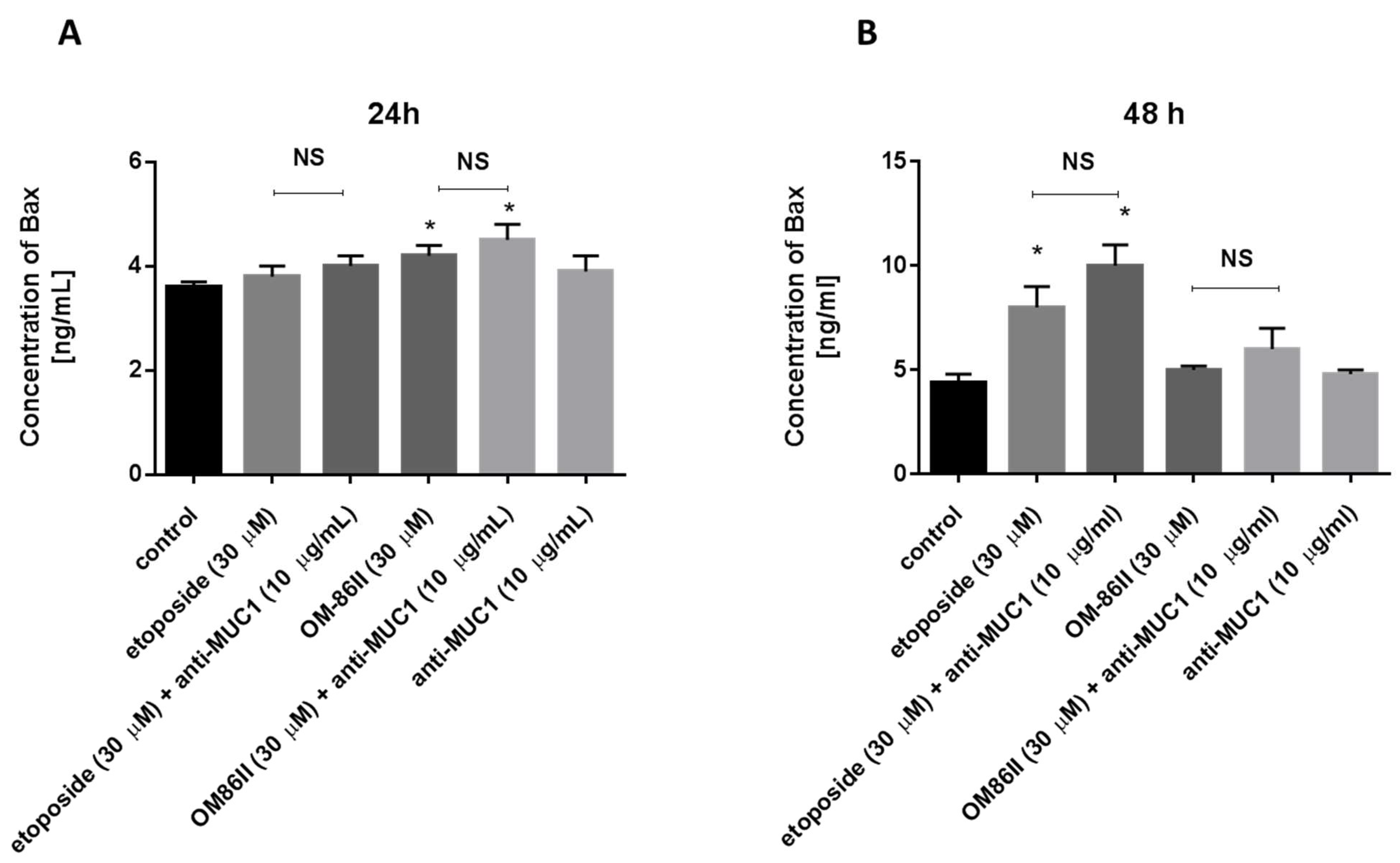
2.4. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Significantly Decreased the Concentrations of mTOR, sICAM1, MMP-9 and Bax after 48 h of Incubation
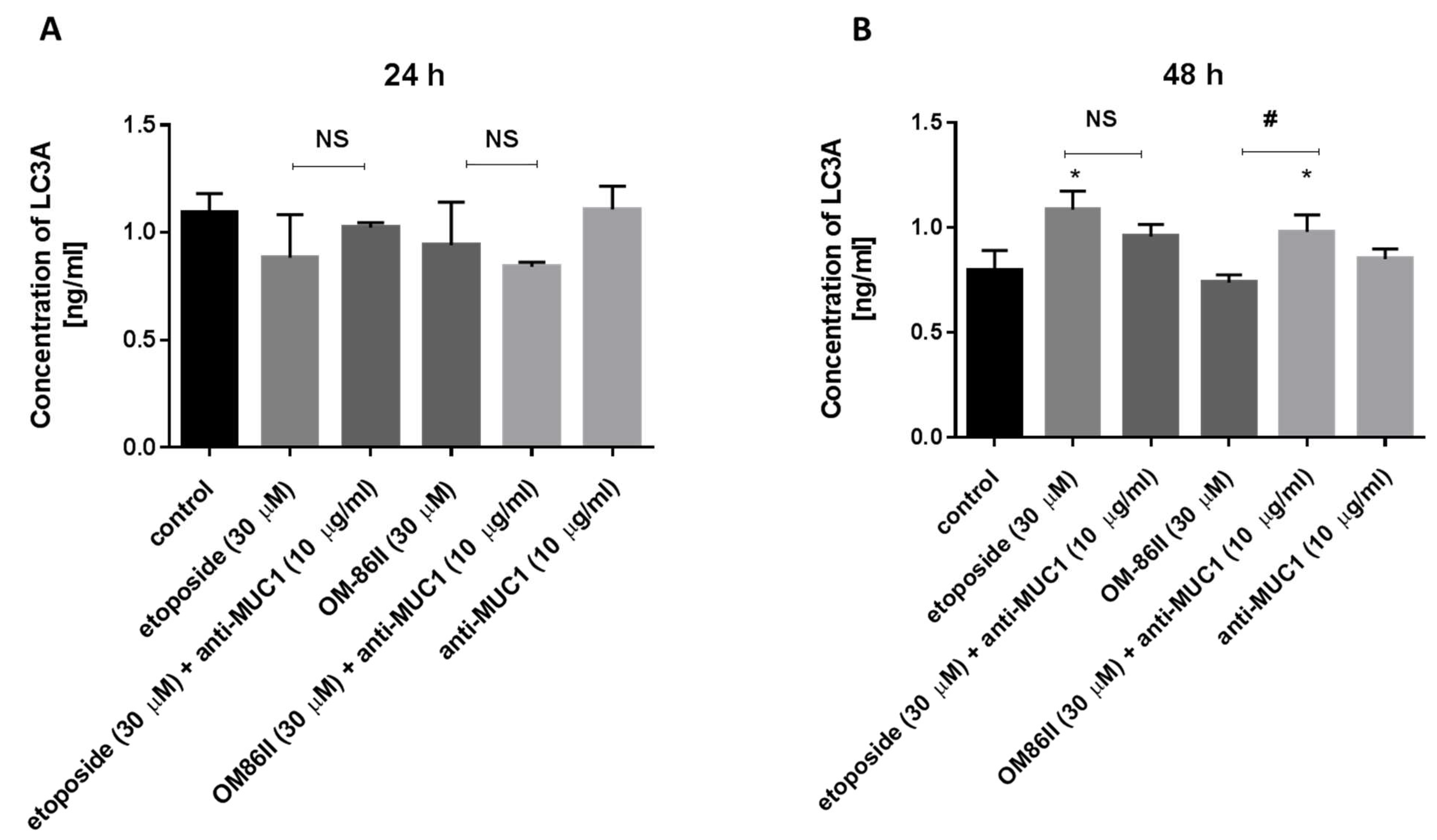
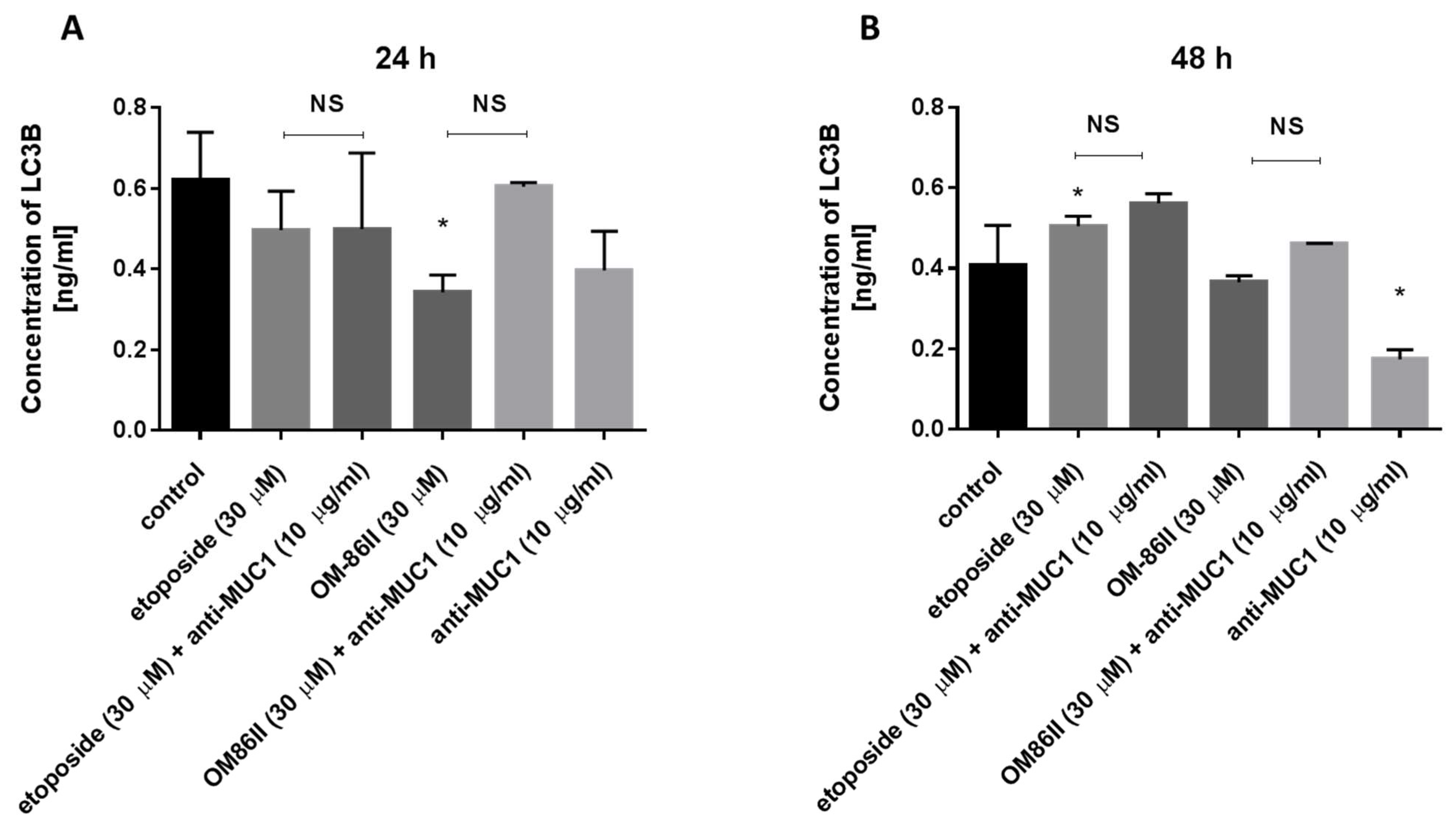
2.5. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Increased LC3A and LC3B Concentrations after 48 h of Incubation in Human Gastric Cancer Cells
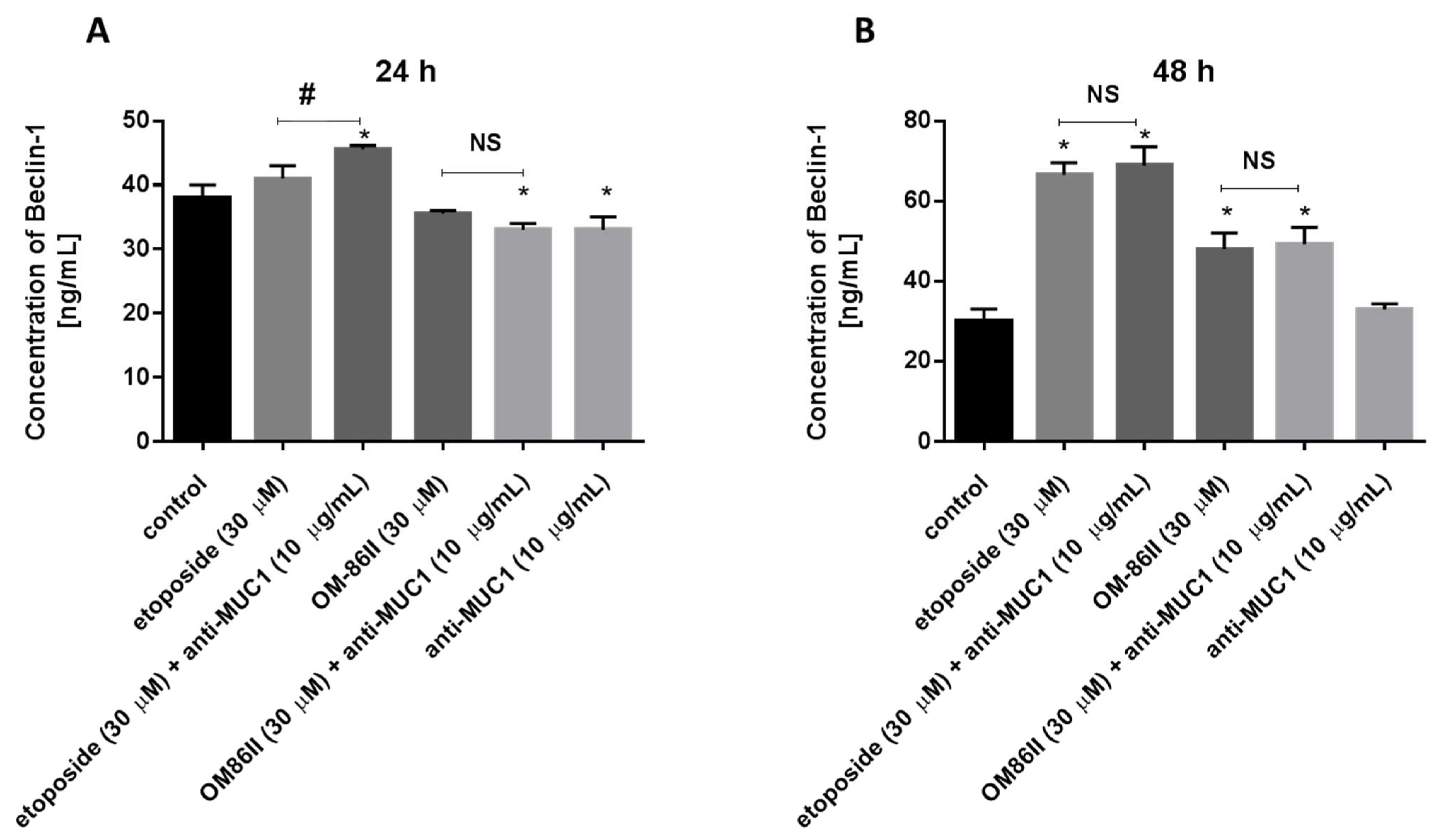
2.6. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Increased Beclin-1 Concentration after 48 h of Incubation in Human Gastric Cancer Cells
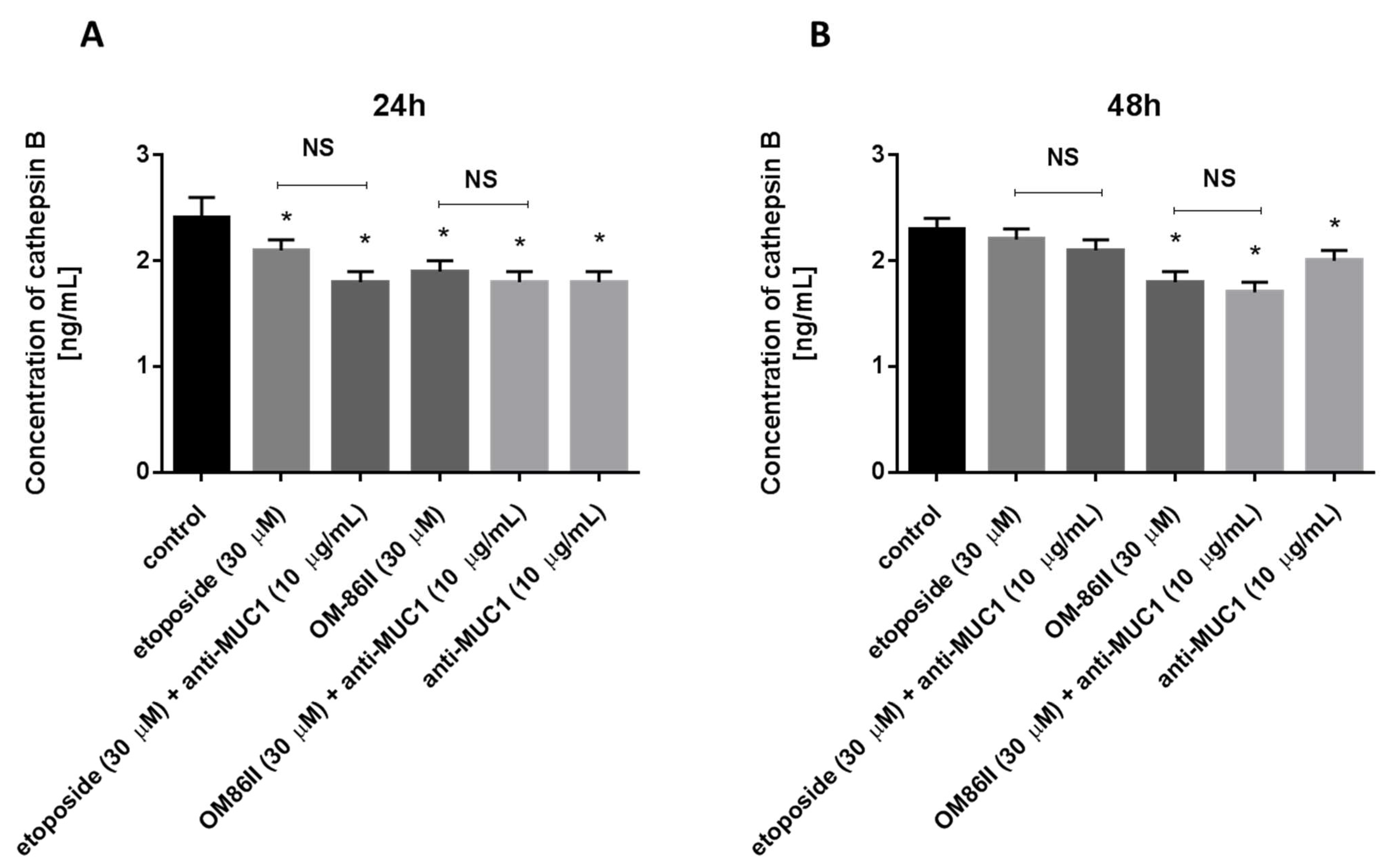
2.7. Novel Diisoquinoline Derivative with Anti-MUC1 Antibody Decreased Cathepsin B Concentration in Human Gastric Cancer Cells
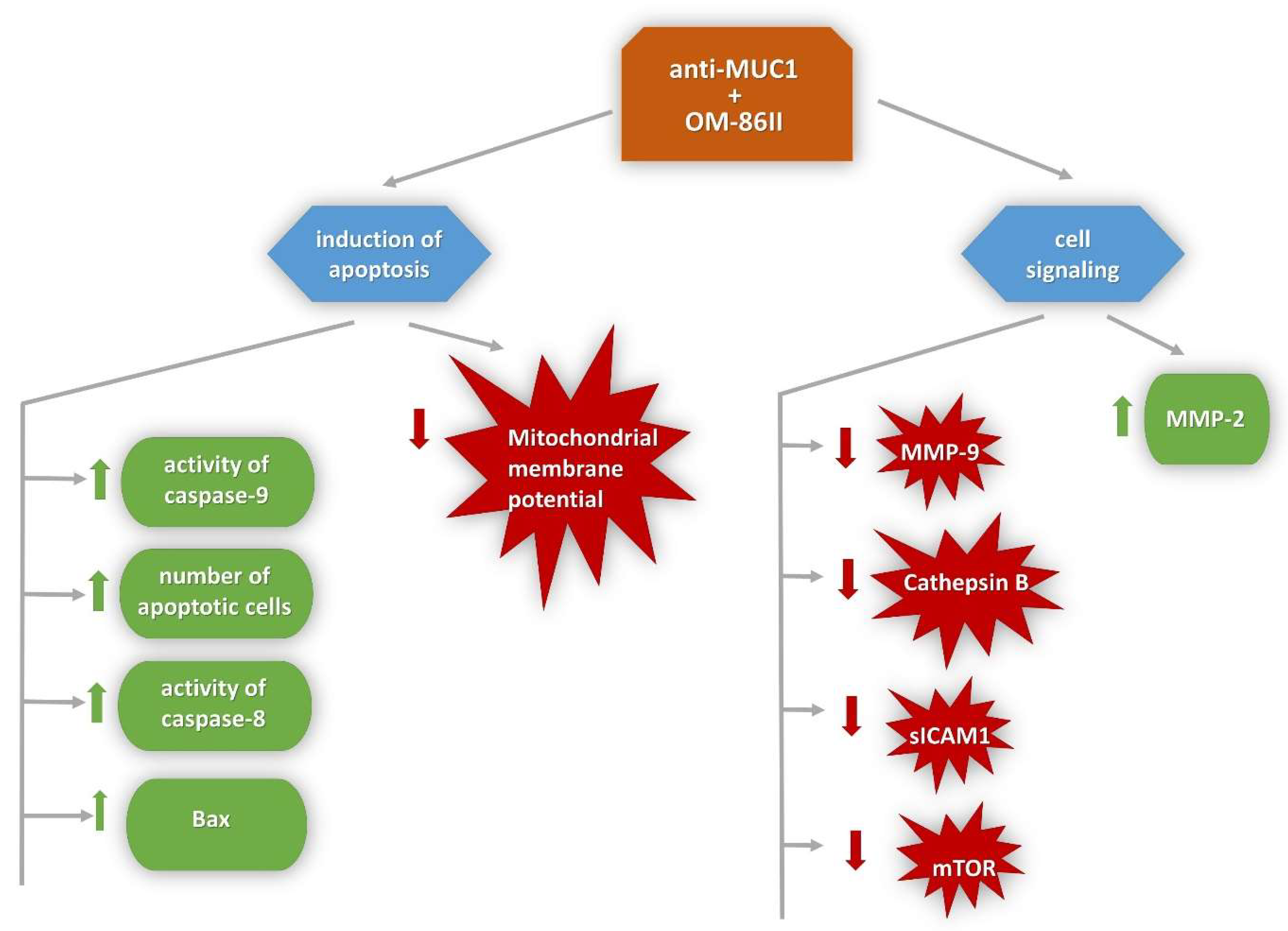
3. Discussion
4. Materials and Methods
4.1. Compounds
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. [3H]thymidine Incorporation Assay
4.5. Dual Acridine Orange/Ethidium Bromide Fluorescent Staining
4.6. Determination of Mitochondrial Membrane Potential
4.7. Caspase-8 Enzymatic Activity Assay
4.8. Caspase-9 Enzymatic Activity Assay
4.9. Determination of mTOR, sICAM1, MMP-2, MMP-9, Bax, LC3A, LC3B, Beclin-1 and Cathepsin B
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bafna, S.; Kaur, S.; Batra, S.K. Membrane-bound mucins: The mechanistic basis for alterations in the growth and survival of cancer cells. Oncogene 2010, 29, 2893–2904. [Google Scholar] [CrossRef] [Green Version]
- Kufe, D.W. Mucins in cancer: Function, prognosis and therapy. Nat. Rev. Cancer 2009, 9, 874–885. [Google Scholar] [CrossRef] [Green Version]
- Pinho, S.S.; Carvalho, S.; Marcos-Pinto, R.; Magalhães, A.; Oliveira, C.; Gu, J.; Dinis-Ribeiro, M.; Carneiro, F.; Seruca, R.; Reis, C.A. Gastric cancer: Adding glycosylation to the equation. Trends Mol. Med. 2013, 19, 664–676. [Google Scholar] [CrossRef]
- İlhan, Ö.; Han, Ü.; Önal, B.; Çelık, S.Y. Prognostic significance of MUC1, MUC2 and MUC5AC expressions in gastric carcinoma. Turk. J. Gastroenterol. 2010, 21, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.K.; Zhang, Q.M.; Zhao, T.H.; Li, Y.Y.; Yi, Y.F. Expression of mucins and E-cadherin in gastric carcinoma and their clinical significance. World J. Gastroenterol. 2004, 10, 3044–3047. [Google Scholar] [CrossRef] [PubMed]
- Raina, D.; Kharbanda, S.; Kufe, D. The MUC1 oncoprotein activates the anti-apoptotic phosphoinositide 3-kinase/Akt and Bcl-xL pathways in rat 3Y1 fibroblasts. J. Biol. Chem. 2004, 279, 20607–20612. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Xu, H.; Kufe, D. Human mucin 1 oncoprotein represses transcription of the p53 tumor suppressor gene. Cancer Res. 2007, 67, 1853–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahn, J.J.; Chow, J.W.; Horne, G.J.; Mah, B.K.; Emerman, J.T.; Hoffman, P.; Hugh, J.C. MUC1 mediates transendothelial migration in vitro by ligating endothelial cell ICAM-1. Clin. Exp. Metastasis 2005, 22, 475–483. [Google Scholar] [CrossRef]
- Karar, J.; Maity, A. PI3K/AKT/mTOR Pathway in Angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Mabuchi, S.; Kuroda, H.; Takahashi, R.; Sasano, T. The PI3K/AKT/mTOR pathway as a therapeutic target in ovarian cancer. Gynecol. Oncol. 2015, 137, 173–179. [Google Scholar] [CrossRef]
- Lheureux, S.; Denoyelle, C.; Ohashi, P.S.; De Bono, J.S.; Mottaghy, F.M. Molecularly targeted therapies in cancer: A guide for the nuclear medicine physician. Eur. J. Nucl. Med. Mol. Imaging 2017, 44, 41–54. [Google Scholar] [CrossRef]
- Martelli, A.M.; Buontempo, F.; McCubrey, J.A. Drug discovery targeting the mTOR pathway. Clin. Sci. 2018, 132, 543–568. [Google Scholar] [CrossRef]
- Malemud, C.J. Matrix metalloproteinases (MMPs) in health and disease: An overview. Front. Biosci. 2006, 11, 1696–1701. [Google Scholar] [CrossRef]
- Verma, S.; Kesh, K.; Ganguly, N.; Jana, S.; Swarnakar, S. Matrix metalloproteinases and gastrointestinal cancers: Impacts of dietary antioxidants. World J. Biol. Chem. 2014, 5, 355–376. [Google Scholar] [CrossRef] [Green Version]
- Fingleton, B. Matrix metalloproteinases: Roles in cancer and metastasis. Front. Biosci. 2006, 11, 479–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirano, T.; Yoshioka, H. Serum cathepsin B levels, urinary excretion of cathepsin B and tissue cathepsin B content in the patients with gastric cancer. Arch. Jpn. Chir. 1993, 62, 217–221. [Google Scholar]
- Herszényi, L.; Plebani, M.; Carraro, P.; De Paoli, M.; Roveroni, G.; Rugge, M.; Cardin, R.; Naccarato, R.; Farinati, F. Role and behavior of cathepsin B and cathepsin L in gastric cancer. Orv. Hetil. 1995, 136, 1315–1318. [Google Scholar]
- Czyzewska, J.; Guzińska-Ustymowicz, K.; Kemona, A.; Bandurski, R. The expression of matrix metalloproteinase 9 and cathepsin B in gastric carcinoma is associated with lymph node metastasis, but not with postoperative survival. Folia Histochem. Cytobiol. 2008, 46, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Baeken, M.W.; Weckmann, K.; Diefenthäler, P.; Schulte, J.; Yusifli, K.; Moosmann, B.; Behl, C.; Hajieva, P. Novel insights into the cellular localization and regulation of the autophagosomal proteins LC3A, LC3B and LC3C. Cells 2020, 18, 2315. [Google Scholar] [CrossRef]
- Wong, Y.C.; Holzbaur, E.L. Optineurin is an autophagy receptor for damaged mitochondria in parkin-mediated mitophagy that is disrupted by an ALS-linked mutation. Proc. Natl. Acad. Sci. USA 2014, 111, 4439–4448. [Google Scholar] [CrossRef] [Green Version]
- McLendon, P.M.; Ferguson, B.S.; Osinska, H.; Bhuiyan, M.S.; James, J.; McKinsey, T.A.; Robbins, J. Tubulin hyperacetylation is adaptive in cardiac proteotoxicity by promoting autophagy. Proc. Natl. Acad. Sci. USA 2014, 111, 5178–5186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Hong, M.J.; Sun, H.; Wang, L.; Shi, X.; Gilbert, B.E.; Corry, D.B.; Kheradmand, F.; Wang, J. Essential role for autophagy in the maintenance of immunological memory against influenza infection. Nat. Med. 2014, 20, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Sarparanta, J.; Garcia-Macia, M.; Singh, R. Autophagy and Mitochondria in obesity and type 2 diabetes. Curr. Diabetes Rev. 2017, 13, 352–369. [Google Scholar] [CrossRef]
- Amaravadi, R.K.; Lippincott-Schwartz, J.; Yin, X.M.; Weiss, W.A.; Takebe, N.; Timmer, W.; DiPaola, R.S.; Lotze, M.T.; White, E. Principles and current strategies for targeting autophagy for cancer treatment. Clin Cancer Res. 2011, 17, 654–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.R.; Yang, Y. Functional role of autophagy in gastric cancer. Oncotarget 2016, 5, 17641–17651. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Luo, Y.; Zou, J.; Ouyang, J.; Cai, Z.; Zeng, X.; Ling, H.; Zeng, T. Autophagy and its role in gastric cancer. Clin. Chim. Acta 2019, 489, 10–20. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, C.; Hu, J.; Su, R.; Zhang, J.; Han, Z.; Chen, H.; Li, Y. Molecular mechanism of Helicobacter pylori-induced autophagy in gastric cancer. Oncol. Lett. 2019, 18, 6221–6227. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Li, N.; Gu, J.; Zhuang, Y.; Li, Q.; Wang, H.G.; Fang, Y.; Yu, B.; Zhang, J.Y.; Xie, Q.H.; et al. Compromised autophagy by MIR30B benefits the intracellular survival of Helicobacter pylori. Autophagy 2012, 8, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Song, J.; Zhou, Y.; Gong, Y.; Liu, H.; Tang, L. Rottlerin promotes autophagy and apoptosis in gastric cancer cell lines. Mol. Med. Rep. 2018, 18, 2905–2913. [Google Scholar] [CrossRef]
- Lazăr, D.C.; Tăban, S.; Cornianu, M.; Faur, A.; Goldiş, A. New advances in targeted gastric cancer treatment. World J. Gastroenterol. 2016, 22, 6776–6799. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, H.K.; Kim, H.S.; Yang, H.K.; Kim, Y.I.; Kim, W.H. MUC1, MUC2, MUC5AC and MUC6 expressions in gastric carcinomas: Their roles as prognostic indicators. Cancer 2001, 92, 1427–1434. [Google Scholar] [CrossRef]
- Pinto-de-Sousa, J.; David, L.; Reis, C.A.; Gomes, R.; Silva, L.; Pimenta, A. Mucins MUC1, MUC2, MUC5AC and MUC6 expression in the evaluation of differentiation and clinico-biological behaviour of gastric carcinoma. Virchows Arch. 2002, 440, 304–310. [Google Scholar] [CrossRef]
- Uen, Y.H.; David, L.; Reis, C.A.; Gomes, R.; Silva, L.; Pimenta, A. Clinical significance of MUC1 andc-Met RT-PCR detection of circulating tumor cells in patients with gastric carcinoma. Clin. Chim. Acta 2006, 367, 55–61. [Google Scholar] [CrossRef]
- Gornowicz, A.; Pawłowska, N.; Czajkowska, A.; Czarnomysy, R.; Bielawska, A.; Bielawski, K.; Michalak, O.; Staszewska-Krajewska, O.; Kałuża, Z. Biological evaluation of octahydropyrazin[2,1-a:5,4-a’]diisoquinoline derivatives as potent anticancer agents. Tumour. Biol. 2017, 39, 1010428317701641. [Google Scholar] [CrossRef] [Green Version]
- Pawłowska, N.; Gornowicz, A.; Bielawska, A.; Surażyński, A.; Szymanowska, A.; Czarnomysy, R.; Bielawski, K. The molecular mechanism of anticancer action of novel octahydropyrazino[2,1-a:5,4-a’]diisoquinoline derivatives in human gastric cancer cells. Investig. New Drugs 2018, 36, 970–984. [Google Scholar] [CrossRef] [Green Version]
- Lepiarczyk, M.; Kałuża, Z.; Bielawska, A.; Czarnomysy, R.; Gornowicz, A.; Bielawski, K. Cytotoxic activity of octahydropyrazin[2,1-a:5,4-a’]diisoquinoline derivatives in human breast cancer cells. Arch. Pharm. Res. 2015, 38, 628–641. [Google Scholar] [CrossRef]
- MacKenzie, S.H.; Schipper, J.L.; Clark, A.C. The potential for caspases in drug discovery. Curr. Opin. Drug Discov. Devel. 2010, 13, 568–576. [Google Scholar]
- Wolan, D.W.; Zorn, J.A.; Gray, D.C.; Wells, J.A. Small-molecule activators of a proenzyme. Science 2009, 326, 853–858. [Google Scholar] [CrossRef] [Green Version]
- Peterson, Q.; Goode, D.R.; West, D.C.; Ramsey, K.N.; Lee, J.J.; Hergenrother, P.J. PAC-1 activates procaspase-3 in vitro through relief of zinc-mediated inhibition. J. Mol. Biol. 2009, 388, 144–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putt, K.S.; Hen, G.W.; Pearson, J.M.; Sandhorst, J.S.; Hoagland, M.S.; Kwon, J.T.; Hwang, S.K.; Jin, H.; Churchwell, M.I.; Cho, M.H.; et al. Small-molecule activation of procaspase-3 to caspase-3 as a personalized anticancer strategy. Nat. Chem. Biol. 2006, 2, 543–550. [Google Scholar] [CrossRef]
- Li, X. Inhibitory effect of recombinant adenowirus carrying immunocaspase-3 on hepatocellular carcinoma. Biochem. Biohys. Res. Commun. 2007, 358, 489–494. [Google Scholar] [CrossRef]
- Chen, K.; Shou, L.M.; Lin, F.; Duan, W.M.; Wu, M.Y.; Xie, X.; Xie, Y.F.; Li, W.; Tao, M. Artesunate induces G2/M cell cycle arrest through autophagy induction in breast cancer cells. Anticancer Drugs 2014, 25, 652–662. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Rucker, E.B.; Zhou, B.P. Autophagy regulation in the development and treatment of breast cancer. Acta Biochim. Biophys. Sin. 2016, 48, 60–74. [Google Scholar] [CrossRef] [Green Version]
- Shen, M.; Duan, W.M.; Wu, M.Y.; Wang, W.J.; Liu, L.; Xu, M.D.; Zhu, J.; Li, D.M.; Gui, Q.; Lian, L.; et al. Participation of autophagy in the cytotoxicity against breast cancer cells by cisplatin. Oncol. Rep. 2015, 34, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.C.; Lin, C.S.; Tai, W.T.; Liu, C.Y.; Shiau, C.W.; Chen, K.F. Nilotinib induces autophagy in hepatocellular carcinoma through AMPK activation. J. Biol. Chem. 2013, 288, 18249–18259. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.S.; Xing, C.G.; Lin, F.; Fan, X.Q.; Zhao, K.; Qin, Z.H. Blocking NF-κB nuclear translocation leads to p53-related autophagy activation and cell apoptosis. World J. Gastroenterol. 2011, 17, 478–487. [Google Scholar] [CrossRef]
- Onorati, A.V.; Dyczynski, M.; Ojha, R.; Amaravadi, R.K. Targeting autophagy in cancer. Cancer 2018, 124, 3307–3318. [Google Scholar] [CrossRef] [Green Version]
- Shoji-Kawata, S.; Sumpter, R.; Leveno, M.; Campbell, G.R.; Zou, Z.; Kinch, L.; Wilkins, A.D.; Sun, Q.; Pallauf, K.; MacDuff, D.; et al. Identification of a candidate therapeutic autophagy-inducing peptide. Nature 2013, 494, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.L.; Yuan, L.; Tang, Y.C.; Xu, Z.Y.; Xu, H.D.; Cheng, X.D.; Qin, J.J. The role of autophagy in gastric cancer chemoresistance: Friend or foe? Front. Cell Dev. Biol. 2020, 8, 621428. [Google Scholar] [CrossRef]
- Kufe, D.W. MUC1-C Oncoprotein as a Target in Breast Cancer; Activation of Signaling Pathways and Therapeutic Approaches. Oncogene 2013, 32, 1073–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahn, J.J.; Shen, Q.; Mah, B.K.; Hugh, J.C. MUC1 initiates a calcium signal after ligation by intercellular adhesion molecule-1. J. Biol. Chem. 2004, 279, 29386–29390. [Google Scholar] [CrossRef] [Green Version]
- Bramhall, S.R.; Hallissey, M.T.; Whiting, J.; Scholefield, J.; Tierney, G.; Stuart, R.C.; Hawkins, R.E.; McCulloch, P.; Maughan, T.; Brown, P.D.; et al. Marimastat as maintenance therapy for patients with advanced gastric cancer: A randomised trial. Br. J. Cancer 2002, 86, 1864–1870. [Google Scholar] [CrossRef] [PubMed]
- McKerrow, J.H.; Bhargava, V.; Hansell, E.; Huling, S.; Kuwahara, T.; Matley, M.; Coussens, L.; Warren, R. A functional proteomics screen of proteases in colorectal carcinoma. Mol. Med. 2000, 6, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Srisomsap, C.; Subhasitanont, P.; Otto, A.; Mueller, E.C.; Punyarit, P.; Wittmann-Liebold, B.; Svasti, J. Detection of cathepsin B up-regulation in neoplastic thyroid tissues by proteomic analysis. Proteomics 2002, 2, 706–712. [Google Scholar] [CrossRef]
- Wulfkuhle, J.D.; Sgroi, D.C.; Krutzsch, H.; McLean, K.; McGarvey, K.; Knowlton, M.; Chen, S.; Shu, H.; Sahin, A.; Kurek, R.; et al. Proteomics of human breast ductal carcinoma in situ. Cancer Res. 2002, 62, 6740–6749. [Google Scholar]
- Ebert, M.P.; Krüger, S.; Fogeron, M.L.; Lamer, S.; Chen, J.; Pross, M.; Schulz, H.U.; Lage, H.; Heim, S.; Roessner, A.; et al. Overexpression of cathepsin B in gastric cancer identified by proteome analysis. Proteomics 2005, 5, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Tummalapalli, P.; Spomar, D.; Gondi, C.S.; Olivero, W.C.; Gujrati, M.; Dinh, D.H.; Rao, J.S. RNAi-mediated abrogation of cathepsin B and MMP-9 gene expression in a malignant meningioma cell line leads to decreased tumor growth, invasion and angiogenesis. Int. J. Oncol. 2007, 31, 1039–1050. [Google Scholar]
- Kałuża, Z.; Mostowicz, D.; Dołęga, G.; Wójcik, R. A new route to optically pure highly functionalized tetrahydroisoquinolines with a quaternary carbon stereocenter. Tetrahedron 2008, 64, 2321–2328. [Google Scholar] [CrossRef]
- Gornowicz, A.; Szymanowski, W.; Bielawska, A.; Szymanowska, A.; Czarnomysy, R.; Kałuża, Z.; Bielawski, K. Monoclonal anti-MUC1 antibody with novel octahydropyrazino[2,1-a:5,4-a’]diisoquinoline derivative as a potential multi-targeted strategy in MCF-7 breast cancer cells. Oncol. Rep. 2019, 42, 1391–1403. [Google Scholar] [CrossRef]
- Carmichael, J.; Degraff, W.; Gazdar, A.; Minna, J.; Mitchell, J. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of radiosensitivity. Cancer Res. 1987, 47, 943–946. [Google Scholar] [PubMed]
- Singh, S.K.; Moretta, D.; Almaguel, F.; Wall, N.R.; De León, M.; De León, D. Differential effect of proIGF-II and IGF-II on resveratrol induced cell death by regulating surviving cellular localization and mitochondrial depolarization in breast cancer cells. Growth Factors 2007, 25, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Czarnomysy, R.; Bielawski, K.; Muszynska, A.; Bielawska, A.; Gornowicz, A. Biological evaluation of dimethylpyridine-platinum complexes with potent antiproliferative activity. J. Enzym. Inhib. Med. Chem. 2016, 31, 150–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gornowicz, A.; Szymanowski, W.; Bielawski, K.; Kałuża, Z.; Michalak, O.; Bielawska, A. Mucin 1 as a Molecular Target of a Novel Diisoquinoline Derivative Combined with Anti-MUC1 Antibody in AGS Gastric Cancer Cells. Molecules 2021, 26, 6504. https://doi.org/10.3390/molecules26216504
Gornowicz A, Szymanowski W, Bielawski K, Kałuża Z, Michalak O, Bielawska A. Mucin 1 as a Molecular Target of a Novel Diisoquinoline Derivative Combined with Anti-MUC1 Antibody in AGS Gastric Cancer Cells. Molecules. 2021; 26(21):6504. https://doi.org/10.3390/molecules26216504
Chicago/Turabian StyleGornowicz, Agnieszka, Wojciech Szymanowski, Krzysztof Bielawski, Zbigniew Kałuża, Olga Michalak, and Anna Bielawska. 2021. "Mucin 1 as a Molecular Target of a Novel Diisoquinoline Derivative Combined with Anti-MUC1 Antibody in AGS Gastric Cancer Cells" Molecules 26, no. 21: 6504. https://doi.org/10.3390/molecules26216504