
Comparative Study of Bioactivity and Safety Evaluation of Ethanolic Extracts of Zanthoxylum schinifolium Fruit and Pericarp



Abstract
:1. Introduction
2. Results and Discussion
2.1. ZS Extract Yield
2.2. ZS Extract Safety
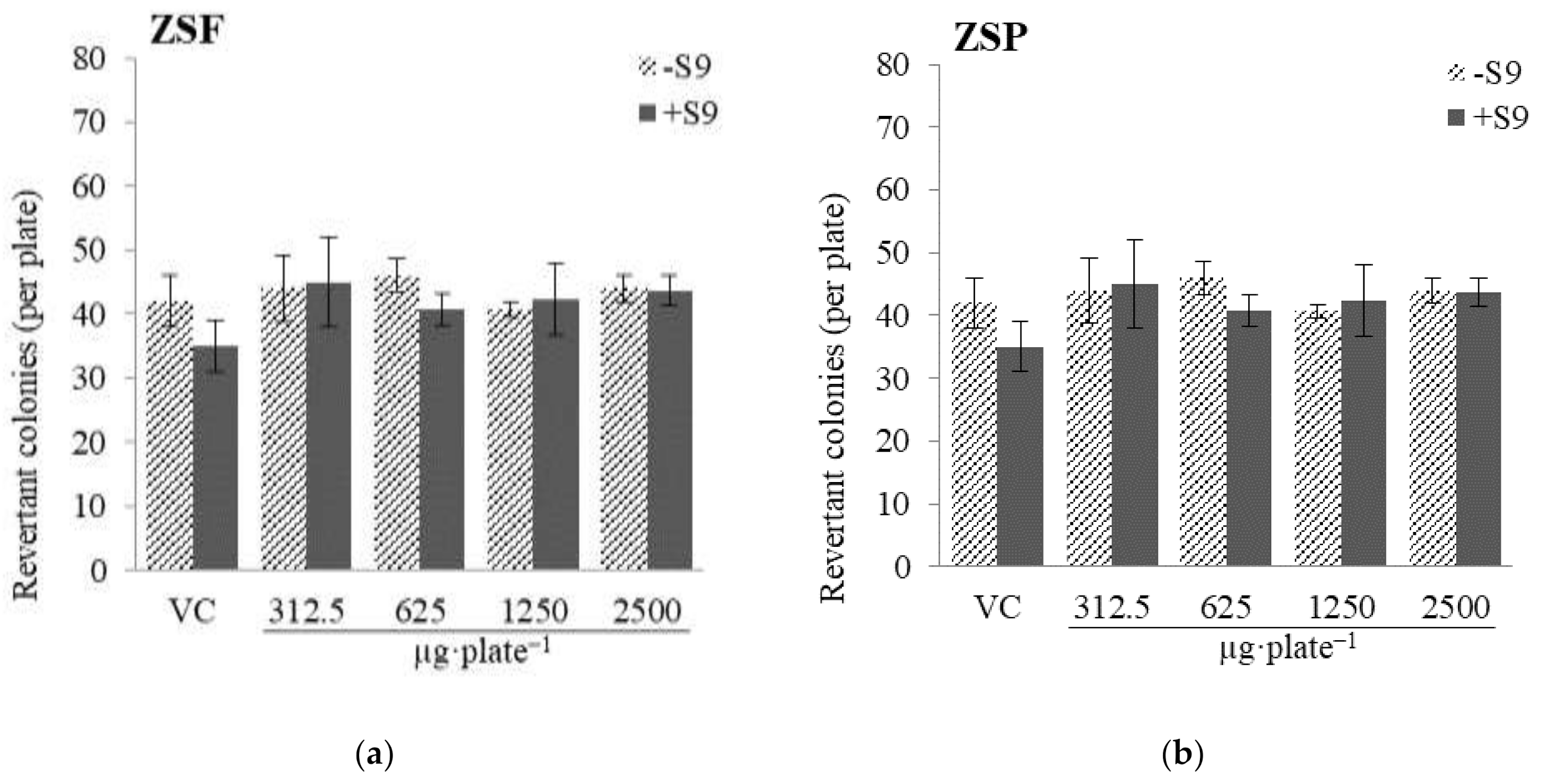
2.2.1. Mutagenicity
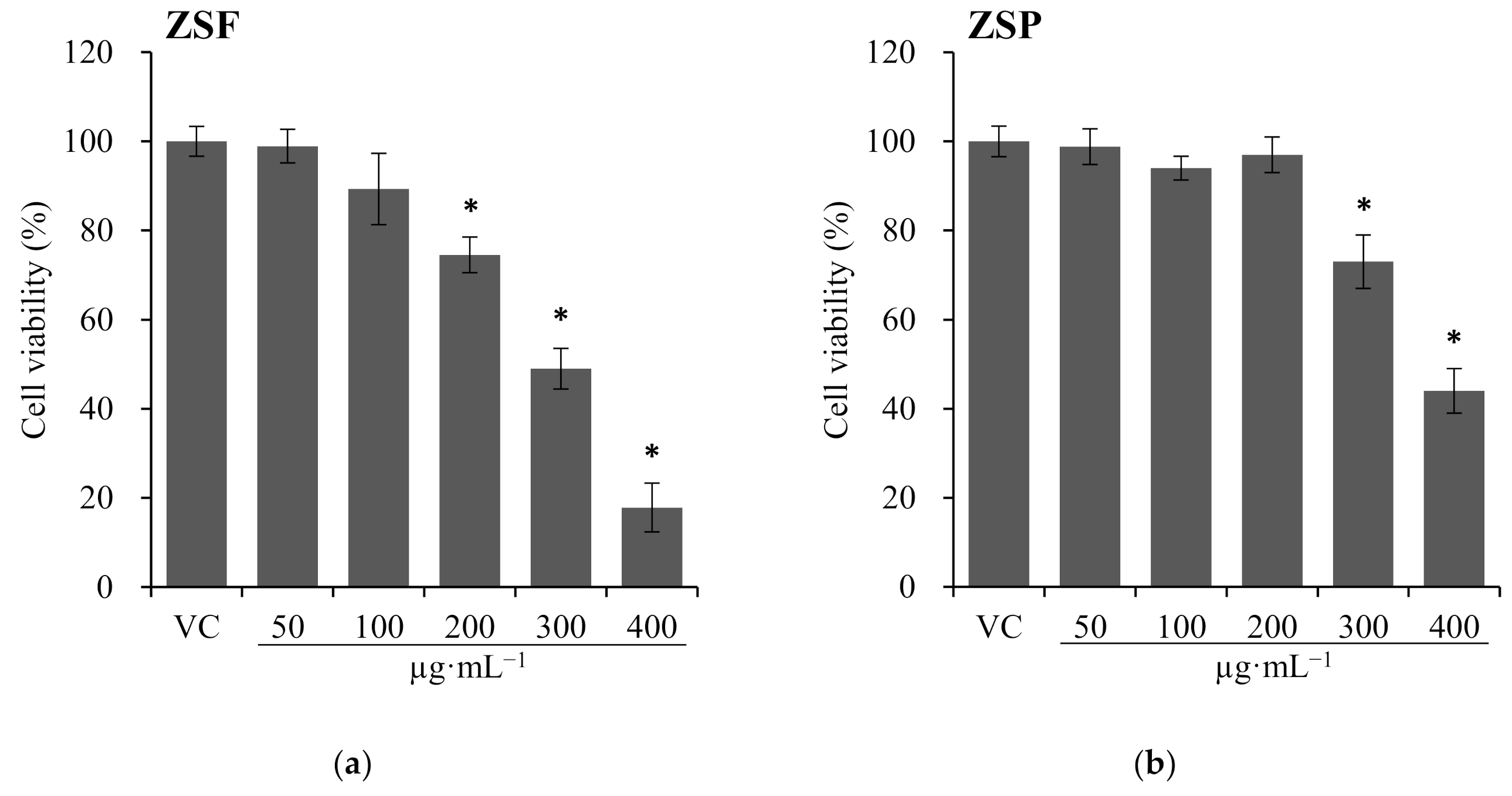
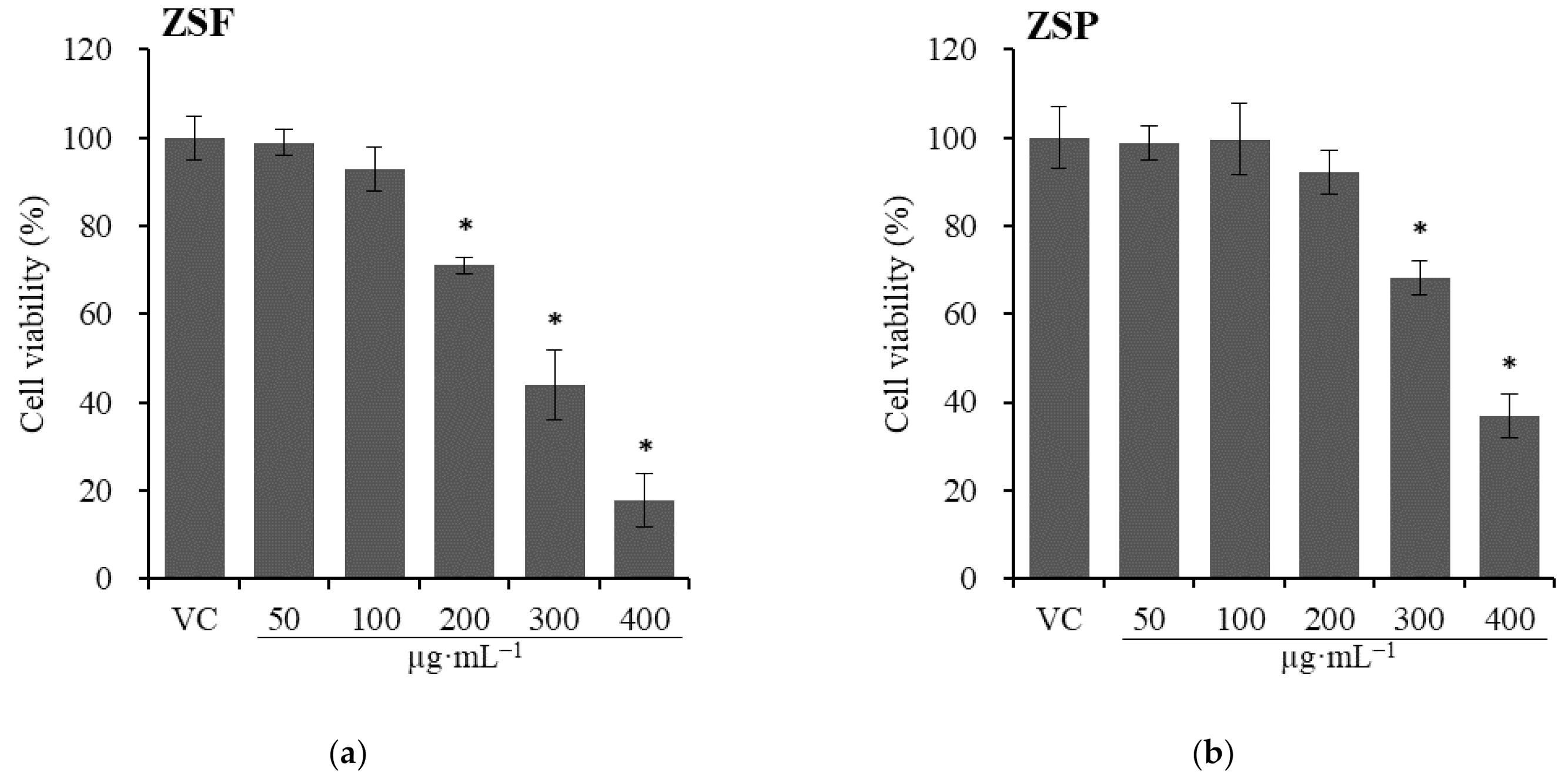
2.2.2. Cytotoxicity
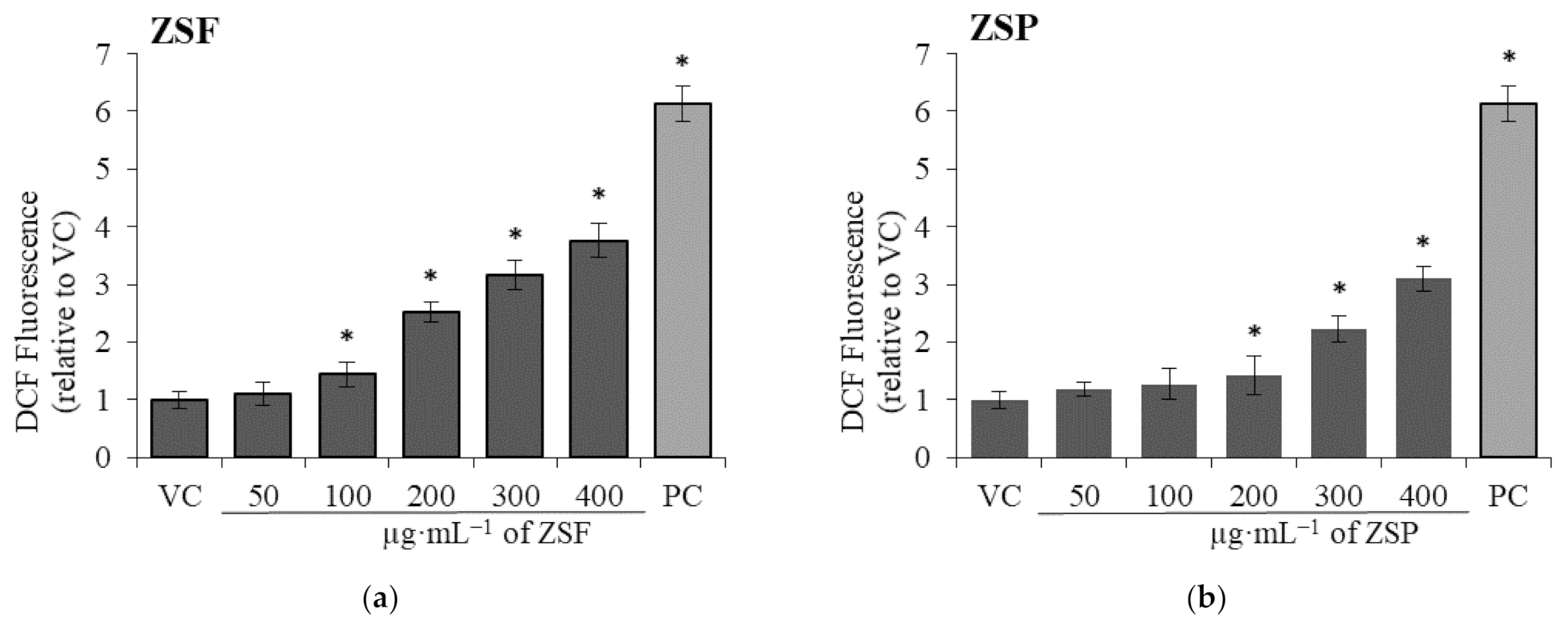
2.2.3. Intracellular ROS Generation
2.3. ZS Extract Bioactivity
2.3.1. Antimutagenicity
2.3.2. Antioxidant Activity
Total Polyphenol Content (TPC) and Total Flavonoid Content (TFC)
Free Radical Scavenging Activity
2.3.3. Anti-Inflammatory Activity
3. Materials and Methods
3.1. Plant Materials
3.2. Chemicals/Reagents
3.3. Strain and Cell/Medium
3.4. Preparation of ZSF and ZSP Ethanol Extracts
3.5. Safety Evaluations
3.5.1. Strain Culture
3.5.2. Mutagenicity Assay
3.5.3. Cytotoxicity Assay
Cell Culture
Neutral Red Uptake (NRU) Assay
WST Assay
3.5.4. Measurement of Intracellular ROS Generation
3.6. Evaluation of Bioactivity of ZS Extracts
3.6.1. Antimutagenicity of ZS Extracts
3.6.2. Antioxidant Activity of ZS Extracts
Total Polyphenol Content
Total Flavonoid Content
DPPH Radical Scavenging Ability
ABTS Radical Scavenging Ability
3.6.3. Anti-Inflammatory Activity of ZS Extract through ELISA Kit
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Clark, N.M. Management of chronic disease by patients. Annu. Rev. Public Health 2003, 24, 289–313. [Google Scholar] [CrossRef] [PubMed]
- Hueda, M.C. Functional Food: Improve Health through Adequate Food; IntechOpen: Rijeka, Croatia, 2017; pp. 143–144. [Google Scholar]
- Manikandan, D.; Jeyachandran, V.; Manikandan, A. Biological Activity Sources from Traditionally Usedtribe and Herbal Plant Material. Int. J. Pure Appl. Math. 2018, 119, 4509–4531. [Google Scholar]
- Słoczyńska, K.; Powroźnik, B.; Pękala, E.; Waszkielewicz, A.M. Antimutagenic compounds and their possible mechanisms of action. J. Appl. Genet. 2014, 55, 273–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertram, J.S. The molecular biology of cancer. Mol. Asp. Med. 2000, 21, 167–223. [Google Scholar] [CrossRef]
- Bhattacharya, S. Natural antimutagens: A review. Res. J. Med. Plants 2011, 5, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Bayr, H. Reactive oxygen species. Crit. Care Med. 2005, 33, S498–S501. [Google Scholar] [CrossRef]
- Jena, N. DNA damage by reactive species: Mechanisms, mutation and repair. J. Biosci. 2012, 37, 503–517. [Google Scholar] [CrossRef]
- Schumacker, P.T. Reactive oxygen species in cancer cells: Live by the sword, die by the sword. Cancer Cell 2006, 10, 175–176. [Google Scholar] [CrossRef] [Green Version]
- Liou, G.-Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Hileman, E.O.; Liu, J.; Albitar, M.; Keating, M.J.; Huang, P. Intrinsic oxidative stress in cancer cells: A biochemical basis for therapeutic selectivity. Cancer Chemother. Pharmacol. 2004, 53, 209–219. [Google Scholar] [CrossRef]
- Murphy, K.; Weaver, C. Janeway’s Immunobiology; Garland Science: New York, NY, USA, 2016; pp. 10–11. [Google Scholar]
- Delves, P.J.; Roitt, I.M. The immune system. N. Engl. J. Med. 2000, 343, 37–49. [Google Scholar] [CrossRef]
- Reid, A.H.; Taubenberger, J.K.; Fanning, T.G. The 1918 Spanish influenza: Integrating history and biology. Microbes Infect. 2001, 3, 81–87. [Google Scholar] [CrossRef]
- Wong, S.S.; Yuen, K.-Y. Avian influenza virus infections in humans. Chest 2006, 129, 156–168. [Google Scholar] [CrossRef]
- Erdmann, N.; Heath, S.L. Cytokine storm syndrome as a manifestation of primary HIV infection. In Cytokine Storm Syndrome; Springer: Berlin/Heidelberg, Germany, 2019; pp. 299–306. [Google Scholar]
- Fajgenbaum, D.C.; June, C.H. Cytokine storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Mann, J. Natural products as immunosuppressive agents. Nat. Prod. Rep. 2001, 18, 417–430. [Google Scholar] [CrossRef] [PubMed]
- Pagare, S.; Bhatia, M.; Tripathi, N.; Pagare, S.; Bansal, Y. Secondary metabolites of plants and their role: Overview. Curr. Trends Biotechnol. Pharm. 2015, 9, 293–304. [Google Scholar]
- Baldino, L.; Scognamiglio, M.; Reverchon, E. Supercritical fluid technologies applied to the extraction of compounds of industrial interest from Cannabis sativa L. and to their pharmaceutical formulations: A review. J. Supercrit. Fluids 2020, 165, 104960. [Google Scholar] [CrossRef]
- Li, W.; Sun, Y.N.; Yan, X.T.; Yang, S.Y.; Kim, E.-J.; Kang, H.K.; Kim, Y.H. Coumarins and lignans from Zanthoxylum schinifolium and their anticancer activities. J. Agric. Food Chem. 2013, 61, 10730–10740. [Google Scholar] [CrossRef]
- Hu, L.; Wang, K.; Wang, Z.; Liu, J.; Wang, K.; Zhang, J.; Luo, Z.; Xue, Y.; Zhang, Y.; Zhang, Y. A new megastigmane sesquiterpenoid from Zanthoxylum schinifolium Sieb. et Zucc. Molecules 2016, 21, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Yang, S.Y.; Yan, X.T.; Sun, Y.N.; Song, S.B.; Kang, H.K.; Kim, Y.H. NF-κB inhibitory activities of glycosides and alkaloids from Zanthoxylum schinifolium stems. Chem. Pharm. Bull. 2014, 62, 196–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-F.; Fan, L.; Tian, M.; Qi, X.-S.; Liu, J.-X.; Feng, J.-B.; Du, S.-S.; Su, X.; Wang, Y.-Y. Radiosensitizing effect of schinifoline from Zanthoxylum schinifolium Sieb et Zucc on human non-small cell lung cancer A549 cells: A preliminary in vitro investigation. Molecules 2014, 19, 20128–20138. [Google Scholar] [CrossRef] [Green Version]
- Min, B.K.; Hyun, D.G.; Jeong, S.Y.; Kim, Y.H.; Ma, E.S.; Woo, M.H. A new cytotoxic coumarin, 7-[(E)-3′, 7′-dimethyl-6′-oxo-2′, 7′-octadienyl] oxy coumarin, from the leaves of Zanthoxylum schinifolium. Arch. Pharmacal Res. 2011, 34, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Jun, D.Y.; Kim, J.S.; Park, H.S.; Han, C.R.; Fang, Z.; Woo, M.H.; Rhee, I.K.; Kim, Y.H. Apoptogenic activity of auraptene of Zanthoxylum schinifolium toward human acute leukemia Jurkat T cells is associated with ER stress-mediated caspase-8 activation that stimulates mitochondria-dependent or-independent caspase cascade. Carcinogenesis 2007, 28, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.S.; Huong, D.T.L.; Bae, K.H.; Lee, M.K.; Kim, Y.H. Monoamine oxidase inhibitory coumarin from Zanthoxylum schinifolium. Planta Med. 2002, 68, 84–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-S.; Lin, Y.-C.; Tsai, I.-L.; Teng, C.-M.; Ko, F.-N.; Ishikawa, T.; Ishii, H. Coumarins and anti-platelet aggregation constituents from Zanthoxylum schinifolium. Phytochemistry 1995, 39, 1091–1097. [Google Scholar] [CrossRef]
- Lee, Y.J.; Yoon, J.J.; Lee, S.M.; Kim, J.S.; Kang, D.G.; Lee, H.S. Inhibitory effect of Zanthoxylum schinifolium on vascular smooth muscle proliferation. Immunopharmacol. Immunotoxicol. 2012, 34, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Chon, S.-U.; Heo, B.-G.; Park, Y.-S.; Kim, D.-K.; Gorinstein, S. Total phenolics level, antioxidant activities and cytotoxicity of young sprouts of some traditional Korean salad plants. Plant Foods Hum. Nutr. 2009, 64, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Diao, W.-R.; Hu, Q.-P.; Feng, S.-S.; Li, W.-Q.; Xu, J.-G. Chemical composition and antibacterial activity of the essential oil from green huajiao (Zanthoxylum schinifolium) against selected foodborne pathogens. J. Agric. Food Chem. 2013, 61, 6044–6049. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.-S.; Oh, Y.N.; Park, J.; Lee, J.Y.; Jin, S.; Hyun, S.K.; Hwang, H.J.; Kwon, H.J.; Kim, B.W. Anti-oxidant, anti-melanogenic, and anti-inflammatory activities of Zanthoxylum schinifolium extract and its solvent fractions. Microbiol. Biotechnol. Lett. 2012, 40, 371–379. [Google Scholar] [CrossRef]
- Wang, C.F.; Yang, K.; Zhang, H.M.; Cao, J.; Fang, R.; Liu, Z.L.; Du, S.S.; Wang, Y.Y.; Deng, Z.W.; Zhou, L. Components and insecticidal activity against the maize weevils of Zanthoxylum schinifolium fruits and leaves. Molecules 2011, 16, 3077–3088. [Google Scholar] [CrossRef]
- Kwon, H.-J.; Hwang, J.; Lee, J.; Chae, S.-K.; Lee, J.-H.; Kim, J.-H.; Hwang, K.-S.; Kim, E.-C.; Park, Y.-D. Analysis and investigation of chemical stability on phenolic compounds in Zanthoxylum schinifolium-containing dentifrices. J. Liq. Chromatogr. Relat. Technol. 2014, 37, 1685–1701. [Google Scholar] [CrossRef]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.-H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Zhang, Z.; Lei, Z.; Yang, Y.; Sugiura, N. Comparative study of antioxidant activity and antiproliferative effect of hot water and ethanol extracts from the mushroom Inonotus obliquus. J. Biosci. Bioeng. 2009, 107, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Custer, L.; Sweder, K. The role of genetic toxicology in drug discovery and optimization. Curr. Drug Metab. 2008, 9, 978–985. [Google Scholar] [CrossRef]
- Martincorena, I.; Campbell, P.J. Somatic mutation in cancer and normal cells. Science 2015, 349, 1483–1489. [Google Scholar] [CrossRef]
- Stratton, M.R.; Campbell, P.J.; Futreal, P.A. The cancer genome. Nature 2009, 458, 719–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, K.M.; Hudson, T.J.; McPherson, J.D. Unraveling the genetics of cancer: Genome sequencing and beyond. Annu. Rev. Genom. Hum. Genet. 2011, 12, 407–430. [Google Scholar] [CrossRef]
- Lee, S.-W.; Lim, J.-M.; Mohan, H.; Seralathan, K.-K.; Park, Y.-J.; Lee, J.-H.; Oh, B.-T. Enhanced bioactivity of Zanthoxylum schinifolium fermented extract: Anti-inflammatory, anti-bacterial, and anti-melanogenic activity. J. Biosci. Bioeng. 2020, 129, 638–645. [Google Scholar] [CrossRef]
- Lim, H.B.; Lee, H.R. Safety and biological activity evaluation of Uncaria rhynchophylla ethanolic extract. Drug Chem. Toxicol. 2020, 1–12. [Google Scholar] [CrossRef]
- Ates, G.; Vanhaecke, T.; Rogiers, V.; Rodrigues, R.M. Assaying cellular viability using the neutral red uptake assay. In Cell Viability Assays; Springer: Berlin/Heidelberg, Germany, 2017; pp. 19–26. [Google Scholar]
- Babich, H.; Schuck, A.G.; Weisburg, J.H.; Zuckerbraun, H.L. Research strategies in the study of the pro-oxidant nature of polyphenol nutraceuticals. J. Toxicol. 2011, 2011. [Google Scholar] [CrossRef]
- Yaffe, P.B.; Power Coombs, M.R.; Doucette, C.D.; Walsh, M.; Hoskin, D.W. Piperine, an alkaloid from black pepper, inhibits growth of human colon cancer cells via G1 arrest and apoptosis triggered by endoplasmic reticulum stress. Mol. Carcinog. 2015, 54, 1070–1085. [Google Scholar] [CrossRef]
- Bombaça, A.C.S.; Von Dossow, D.; Barbosa, J.M.C.; Paz, C.; Burgos, V.; Menna-Barreto, R.F.S. TrypanocidalActivity of Natural Sesquiterpenoids Involves Mitochondrial Dysfunction, ROS Production and Autophagic Phenotype in Trypanosoma cruzi. Molecules 2018, 23, 2800. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, A.; Dey, D.; Ghosh, D.; Lai, T.K.; Bhuvanesh, N.; Dolui, S.; Velayutham, R.; Acharya, K. Astrakurkurone, a sesquiterpenoid from wild edible mushroom, targets liver cancer cells by modulating bcl-2 family proteins. IUBMB Life 2019, 71, 992–1002. [Google Scholar] [CrossRef]
- Zeouk, I.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Bethencourt-Estrella, C.J.; Bazzocchi, I.L.; Bekhti, K.; Lorenzo-Morales, J.; Jiménez, I.A.; Piñero, J.E. Sesquiterpenoids and flavonoids from Inula viscosa induce programmed cell death in kinetoplastids. Biomed. Pharmacother. 2020, 130, 110518. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, S.; Mukherjee, A.; Ghosh, S.; Khuda-Bukhsh, A.R. Condurango glycoside-rich components stimulate DNA damage-induced cell cycle arrest and ROS-mediated caspase-3 dependent apoptosis through inhibition of cell-proliferation in lung cancer, in vitro and in vivo. Environ. Toxicol. Pharmacol. 2014, 37, 300–314. [Google Scholar] [CrossRef]
- Li, S.; Dong, P.; Wang, J.; Zhang, J.; Gu, J.; Wu, X.; Wu, W.; Fei, X.; Zhang, Z.; Wang, Y. Icariin, a natural flavonol glycoside, induces apoptosis in human hepatoma SMMC-7721 cells via a ROS/JNK-dependent mitochondrial pathway. Cancer Lett. 2010, 298, 222–230. [Google Scholar] [CrossRef] [PubMed]
- Slaninová, I.; Pěnčíková, K.; Urbanová, J.; Slanina, J.; Táborská, E. Antitumour activities of sanguinarine and related alkaloids. Phytochem. Rev. 2014, 13, 51–68. [Google Scholar] [CrossRef]
- Kaminskyy, V.; Kulachkovskyy, O.; Stoika, R. A decisive role of mitochondria in defining rate and intensity of apoptosis induction by different alkaloids. Toxicol. Lett. 2008, 177, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.-H.; Luo, S.-J.; Chen, C.-L.; Cheng, J.-H.; Hsieh, C.-Y.; Wang, C.-Y.; Huang, W.-C.; Su, W.-C.; Lin, C.-F. Vinca alkaloids cause aberrant ROS-mediated JNK activation, Mcl-1 downregulation, DNA damage, mitochondrial dysfunction, and apoptosis in lung adenocarcinoma cells. Biochem. Pharmacol. 2012, 83, 1159–1171. [Google Scholar] [CrossRef]
- Domracheva, I.; Kanepe-Lapsa, I.; Jackevica, L.; Vasiljeva, J.; Arsenyan, P. Selenopheno quinolinones and coumarins promote cancer cell apoptosis by ROS depletion and caspase-7 activation. Life Sci. 2017, 186, 92–101. [Google Scholar] [CrossRef]
- Lin, M.-H.; Cheng, C.-H.; Chen, K.-C.; Lee, W.-T.; Wang, Y.-F.; Xiao, C.-Q.; Lin, C.-W. Induction of ROS-independent JNK-activation-mediated apoptosis by a novel coumarin-derivative, DMAC, in human colon cancer cells. Chem.-Biol. Interact. 2014, 218, 42–49. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.; Kiene, R.; Huntsman, S. An antioxidant function for DMSP and DMS in marine algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef]
- Sanmartín-Suárez, C.; Soto-Otero, R.; Sánchez-Sellero, I.; Méndez-Álvarez, E. Antioxidant properties of dimethyl sulfoxide and its viability as a solvent in the evaluation of neuroprotective antioxidants. J. Pharmacol. Toxicol. Methods 2011, 63, 209–215. [Google Scholar] [CrossRef]
- Paik, S.-Y.; Koh, K.-H.; Beak, S.-M.; Paek, S.-H.; Kim, J.-A. The essential oils from Zanthoxylum schinifolium pericarp induce apoptosis of HepG2 human hepatoma cells through increased production of reactive oxygen species. Biol. Pharm. Bull. 2005, 28, 802–807. [Google Scholar] [CrossRef] [Green Version]
- Zahin, M.; Ahmad, I.; Aqil, F. Antioxidant and antimutagenic potential of Psidium guajava leaf extracts. Drug Chem. Toxicol. 2017, 40, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Marnewick, J.L.; Gelderblom, W.C.; Joubert, E. An investigation on the antimutagenic properties of South African herbal teas. Mutat. Res. Genet. Toxicol. Environ. Mutagenes. 2000, 471, 157–166. [Google Scholar] [CrossRef]
- De Flora, S.; Izzotti, A.; D’Agostini, F.; Balansky, R.M.; Noonan, D.; Albini, A. Multiple points of intervention in the prevention of cancer and other mutation-related diseases. Mutat. Res. Fundam. Mol. Mech. Mutagenes. 2001, 480, 9–22. [Google Scholar] [CrossRef]
- Hour, T.-C.; Liang, Y.-C.; Chu, I.-S.; Lin, J.-K. Inhibition of eleven mutagens by various tea extracts, (−) epigallocatechin-3-gallate, gallic acid and caffeine. Food Chem. Toxicol. 1999, 37, 569–579. [Google Scholar] [CrossRef]
- Bhattacharya, U.; Mukhopadhyay, S.; Giri, A.K. Comparative antimutagenic and anticancer activity of three fractions of black tea polyphenols thearubigins. Nutr. Cancer 2011, 63, 1122–1132. [Google Scholar] [CrossRef] [PubMed]
- Di Sotto, A.; Durazzi, F.; Sarpietro, M.G.; Mazzanti, G. Antimutagenic and antioxidant activities of some bioflavours from wine. Food Chem. Toxicol. 2013, 60, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Cheng, S.J.; Zhou, Z.C.; Athar, M.; Khan, W.A.; Bickers, D.R.; Mukhtar, H. Antimutagenic activity of green tea polyphenols. Mutat. Res. Genet. Toxicol. 1989, 223, 273–285. [Google Scholar] [CrossRef]
- Wall, M.E.; Wani, M.C.; Manikumar, G.; Hughes, T.J.; Taylor, H.; McGivney, R.; Warner, J. Plant antimutagenic agents, 3. Coumarins. J. Nat. Prod. 1988, 51, 1148–1152. [Google Scholar] [CrossRef]
- Wall, M.E. Antimutagenic agents from natural products. J. Nat. Prod. 1992, 55, 1561–1568. [Google Scholar] [CrossRef]
- Pereira, D.M.; Valentão, P.; Pereira, J.A.; Andrade, P.B. Phenolics: From Chemistry to Biology. Molecules 2009, 14, 2202–2211. [Google Scholar] [CrossRef]
- RuQin, X.; Rui, W.; Ping, L. Content determination of polyphenol in Zhaotong Zanthoxylum schinifolium Sieb. et Zucc. by Folin-Ciocalteu colorimetry. China Condiment 2019, 44, 140–143. [Google Scholar]
- Kim, B. Anti-oxidant and anti-inflammatory activities of Zanthoxylum schinifolium essential oil. J. Korean Appl. Sci. Technol. 2014, 31, 440–445. [Google Scholar] [CrossRef]
- Oh, S.-M.; Han, W.; Wang, M.-H. Antioxidant and α-glucosidase inhibition activity from different extracts of Zanthoxylum schnifolium fruits. Korean J. Pharmacogn. 2010, 41, 130–135. [Google Scholar]
- Orhan Puskullu, M.; Tekiner, B.; Suzen, S. Recent studies of antioxidant quinoline derivatives. Mini Rev. Med. Chem. 2013, 13, 365–372. [Google Scholar] [PubMed]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Kimmel, D.W.; Meschievitz, M.E.; Hiatt, L.A.; Cliffel, D.E. Multianalyte microphysiometry of macrophage responses to phorbol myristate acetate, lipopolysaccharide, and lipoarabinomannan. Electroanalysis 2013, 25, 1706–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.-H.; Zhao, B.T.; Kim, O.; Lee, J.H.; Choi, J.S.; Min, B.S.; Woo, M.H. Anti-inflammatory terpenylated coumarins from the leaves of Zanthoxylum schinifolium with α-glucosidase inhibitory activity. J. Nat. Med. 2016, 70, 276–281. [Google Scholar] [CrossRef]
- Cao, L.H.; Lee, Y.J.; Kang, D.G.; Kim, J.S.; Lee, H.S. Effect of Zanthoxylum schinifolium on TNF-α-induced vascular inflammation in human umbilical vein endothelial cells. Vasc. Pharmacol. 2009, 50, 200–207. [Google Scholar] [CrossRef]
- Maron, D.M.; Ames, B.N. Revised methods for the Salmonella mutagenicity test. Mutat. Res. Environ. Mutagenes. Relat. Subj. 1983, 113, 173–215. [Google Scholar] [CrossRef]
- Repetto, G.; Del Peso, A.; Zurita, J.L. Neutral red uptake assay for the estimation of cell viability/cytotoxicity. Nat. Protoc. 2008, 3, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Pourmorad, F.; Hosseinimehr, S.; Shahabimajd, N. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afr. J. Biotechnol. 2006, 5. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Yield (%) 1 |
|---|---|
| ZSF | 10.96 |
| ZSP | 25.58 |
| Treatment | Concentration (µg·plate−1) | − S9 | + S9 | ||
|---|---|---|---|---|---|
| Reversion 2 | Inhibition (%) 3 | Reversion | Inhibition (%) | ||
| Control 1 | 0 | 27 ± 3 | - | 33 ± 6 | - |
| 1 | 476 ± 47 | - | 289 ± 7 | - | |
| ZSF | 312.5 | 449 ± 6 | 6.0 c | 255 ± 23 | 13.3 e |
| 625 | 433 ± 11 | 9.6 b,c | 228 ± 18 | 23.8 d | |
| 1250 | 412 ± 21 | 14.2 b | 107 ± 14 | 71.1 c | |
| 2500 | 338 ± 29 | 30.7 a | 37 ± 2 | 98.4 a | |
| ZSP | 312.5 | 476 ± 24 | 0.0 d | 57 ± 11 | 90.6 b |
| 625 | 449 ± 3 | 6.0 c | 39 ± 6 | 97.7 a | |
| 1250 | 392 ± 12 | 18.7 b | 32 ± 1 | 100.4 a | |
| 2500 | 322 ± 4 | 34.3 a | 32 ± 9 | 100.4 a | |
| Treatment | Concentration (µg·plate−1) | − S9 | + S9 | ||
|---|---|---|---|---|---|
| Reversion 2 | Inhibition (%) 3 | Reversion | Inhibition (%) | ||
| Control 1 | 0 | 124 ± 7 | - | 146 ± 8 | - |
| 1 | 412 ± 13 | - | 437 ± 42 | - | |
| ZSF | 312.5 | 336 ± 6 | 26.4 b,c | 413 ± 25 | 8.2 d |
| 625 | 322 ± 32 | 31.2 b | 401 ± 40 | 12.4 d | |
| 1250 | 263 ± 28 | 51.7 a | 370 ± 23 | 23.0 c | |
| 2500 | 253 ± 32 | 55.2 a | 191 ± 38 | 84.5 b | |
| ZSP | 312.5 | 386 ± 20 | 9.0 d | 188 ± 25 | 85.6 b |
| 625 | 375 ± 34 | 12.8 d | 157 ± 7 | 96.2 a | |
| 1250 | 354 ± 8 | 20.1 c | 148 ± 11 | 99.3 a | |
| 2500 | 254 ± 34 | 54.9 a | 147 ± 2 | 99.7 a | |
| Sample | Total Phenol Content (mg GAE·g−1) 1 | Total Flavonoid Content (mg QE·g−1) 2 |
|---|---|---|
| ZSF | 22.7 ± 3.1 a | 18.6 ± 2.5 b |
| ZSP | 62.3 ± 7.9 b | 10.8 ± 2.0 a |
| Sample | DPPH Radical Scavenging Activity | ABTS Radical Scavenging Activity |
|---|---|---|
| IC50 (µg·mL−1) 1 | IC50 (µg·mL−1) | |
| l-Ascorbic acid | 4.1 ± 1.7 a | 22.9 ± 4.3 a |
| ZSF | 281.1 ± 16.8 c | 101.7 ± 11.5 c |
| ZSP | 75.6 ± 6.1 b | 57.4 ± 6.4 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.G.; Lim, J.J.; You, J.S.; Kwon, H.J.; Lim, H.B. Comparative Study of Bioactivity and Safety Evaluation of Ethanolic Extracts of Zanthoxylum schinifolium Fruit and Pericarp. Molecules 2021, 26, 5919. https://doi.org/10.3390/molecules26195919
Kim JG, Lim JJ, You JS, Kwon HJ, Lim HB. Comparative Study of Bioactivity and Safety Evaluation of Ethanolic Extracts of Zanthoxylum schinifolium Fruit and Pericarp. Molecules. 2021; 26(19):5919. https://doi.org/10.3390/molecules26195919
Chicago/Turabian StyleKim, Jun Gu, Jae Jung Lim, Ji Sang You, Hyeok Jun Kwon, and Heung Bin Lim. 2021. "Comparative Study of Bioactivity and Safety Evaluation of Ethanolic Extracts of Zanthoxylum schinifolium Fruit and Pericarp" Molecules 26, no. 19: 5919. https://doi.org/10.3390/molecules26195919