
Time Evolution of Microbial Composition and Metabolic Profile for Biogenic Amines and Free Amino Acids in a Model Cucumber Fermentation System Brined with 0.5% to 5.0% Sodium Chloride

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. pH
2.2. Microbiota Profiles
2.2.1. LAB
2.2.2. Enterobacteriaceae
2.2.3. Fungi (Yeast and Molds)
2.2.4. Enterococcus spp.
2.2.5. Microbial Composition after Storage
2.3. BAs and FAAs
2.3.1. Time Evolution of BAs Content
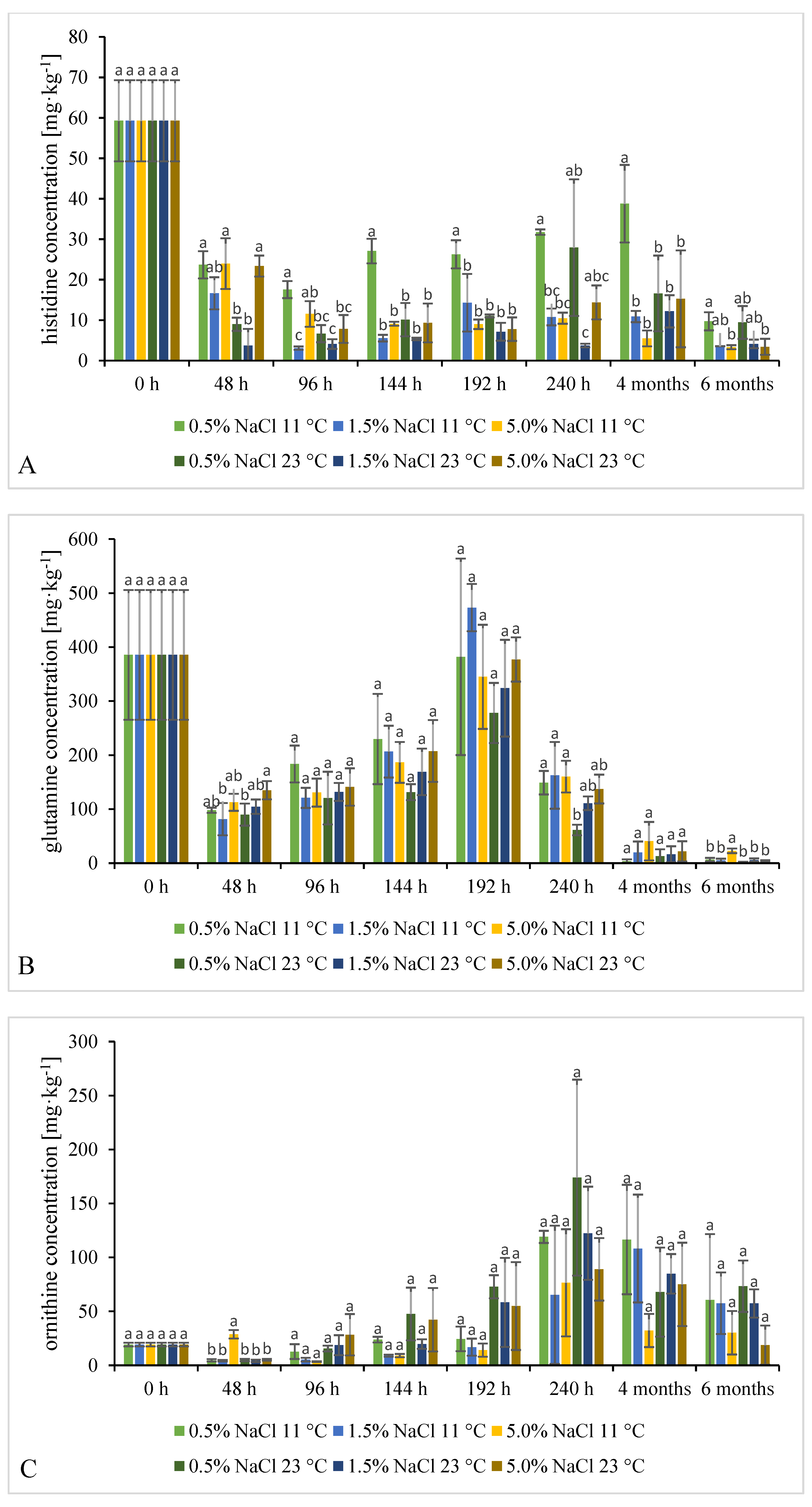
2.3.2. Time Evolution of FAAs Content
2.4. Intercorrelations (PCA)
2.5. Organic Acids Concentration
2.6. Metabolites
3. Materials and Methods
3.1. Chemicals/Reagents
3.2. Samples
3.3. Determination of Bacteria and Fungi
3.4. 16S rRNA Sequencing
3.5. Determination of BAs and FAAs
3.6. Determination of Organic Acids
3.7. Analysis of Bacterial Metabolites
3.8. Statistical Analysis of the Experimental Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Parveen Rani, R. Fermented Fruits and Vegetables of Asia: A Potential Source of Probiotics. Biotechnol. Res. Int. 2014, 2014, 250424. [Google Scholar] [CrossRef]
- Di Cagno, R.; Coda, R. Fermented Foods: Fermented Vegetable Products. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 875–883. [Google Scholar] [CrossRef]
- Lavefve, L.; Marasini, D.; Carbonero, F. Microbial Ecology of Fermented Vegetables and Non-Alcoholic Drinks and Current Knowledge on Their Impact on Human Health. Adv. Food Nutr. Res. 2019, 87, 147–185. [Google Scholar] [CrossRef]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, e00377. [Google Scholar] [CrossRef] [Green Version]
- Peñas, E.; Frias, J.; Sidro, B.; Vidal-Valverde, C. Impact of fermentation conditions and refrigerated storage on microbial quality and biogenic amine content of sauerkraut. Food Chem. 2010, 123, 143–150. [Google Scholar] [CrossRef]
- Wouters, D.; Bernaert, N.; Conjaerts, W.; Van Droogenbroeck, B.; De Loose, M.; De Vuyst, L. Species diversity, community dynamics, and metabolite kinetics of spontaneous leek fermentations. Food Microbiol. 2013, 33, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Li, J.; Liang, F.; Wang, Y.; Guan, Q.; Xie, M. Effects of salt concentration on Chinese sauerkraut fermentation. LWT—Food Sci. Technol. 2016, 69, 169–174. [Google Scholar] [CrossRef]
- Liu, X.; Kuda, T.; Takahashi, H.; Kimura, B. Bacterial and fungal microbiota of spontaneously fermented Chinese products, Rubing milk cake and Yan-cai vegetable pickles. Food Microbiol. 2018, 72, 106–111. [Google Scholar] [CrossRef]
- Nguyen, D.T.L.; Van Hoorde, K.; Cnockaert, M.; De Brandt, E.; Aerts, M.; Binh Thanh, L.; Vandamme, P. A description of the lactic acid bacteria microbiota associated with the production of traditional fermented vegetables in Vietnam. Int. J. Food Microbiol. 2013, 163, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Díaz, I.M.; Hayes, J.; Medina, E.; Anekella, K.; Daughtry, K.; Dieck, S.; Levi, M.; Price, R.; Butz, N.; Lu, Z.; et al. Reassessment of the succession of lactic acid bacteria in commercial cucumber fermentations and physiological and genomic features associated with their dominance. Food Microbiol. 2017, 63, 217–227. [Google Scholar] [CrossRef]
- Ladero, V.; Calles-Enriquez, M.; Fernandez, M.; Alvarez, M.A. Toxicological Effects of Dietary Biogenic Amines. Curr. Nutr. Food Sci. 2010, 6, 145–156. [Google Scholar] [CrossRef]
- Benkerroum, N. Biogenic Amines in Dairy Products: Origin, Incidence, and Control Means. Compr. Rev. Food Sci. Food Saf. 2016, 4, 801–826. [Google Scholar] [CrossRef]
- Guarcello, R.; Angelis, M.D.; Settanni, L.; Formiglio, S.; Gaglio, R.; Minervini, F.; Moschetti, G.; Gobbetti, M. Selection of Amine-Oxidizing Dairy Lactic Acid Bacteria and Identification of the Enzymes and Gene Involved in the Decrease of Biogenic Amines. Appl. Environ. Microbiol. 2016, 23, 6870–6880. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.A.; Morreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Tech. 2014, 39, 146–155. [Google Scholar] [CrossRef] [Green Version]
- Halász, A.; Baráth, Á.; Simon-Sarkadi, L.; Holzapfel, W. Biogenic amines and their production by microorganisms in food. Trends Food Sci. Tech. 1994, 5, 42–49. [Google Scholar] [CrossRef]
- Gardini, F.; Özogul, Y.; Suzzi, G.; Tabanelli, G.; Özogul, F. Technological Factors Affecting Biogenic Amine Content in Foods: A Review. Front. Microbiol. 2016, 7, 1218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özogul, F.; Özogul, Y. Formation of biogenic amines by Gram-negative rods isolated from fresh, spoiled, VP-packed and MAP-packed herring (Clupea harengus). Eur. Food Res. Technol. 2005, 221, 575–581. [Google Scholar] [CrossRef]
- Özogul, F.; Özogul, Y. The ability of biogenic amines and ammonia production by single bacterial cultures. Eur. Food Res. Technol. 2007, 225, 385–394. [Google Scholar] [CrossRef]
- Coton, M.; Delbés-Paus, C.; Irlinger, F.; Desmasures, N.; Le Fleche, A.; Stahl, V.; Montel, M.C.; Coton, E. Diversity and assessment of potential risk factors of Gram-negative isolates associated with French cheeses. Food Microbiol. 2012, 29, 88–98. [Google Scholar] [CrossRef]
- Mokhtar, S.; Mostafa, G.; Taha, R.; Eldeep, G.S.S. Effect of different starter cultures on the biogenic amines production as a critical control point in fresh fermented sausages. Eur. Food Res. Technol. 2012, 235, 527–535. [Google Scholar] [CrossRef]
- Pereira, C. Evidence for proteolytic activity and biogenic amines production in Lactobacillus curvatus and L. homohiochii. Int. J. Food Microbiol. 2001, 68, 211–216. [Google Scholar] [CrossRef]
- Gezginc, Y.; Akyol, I.; Kuley, E.; Özogul, F. Biogenic amines formation in Streptococcus thermophilus isolated from home-made natural yogurt. Food Chem. 2013, 138, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Wunderlichová, L.; Buňková, L.; Koutný, M.; Jančová, P.; Buňka, F. Formation, Degradation, and Detoxification of Putrescine by Foodborne Bacteria: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1012–1030. [Google Scholar] [CrossRef]
- Pachlová, V.; Buňková, L.; Flasarová, R.; Salek, R.-N.; Dlabajová, A.; Butor, I.; Buňka, F. Biogenic amine production by nonstarter strains of Lactobacillus curvatus and Lactobacillus paracasei in the model system of Dutch-type cheese. LWT—Food Sci. Technol. 2018, 97, 730–735. [Google Scholar] [CrossRef]
- Bargossi, E.; Tabanelli, G.; Montanari, C.; Lanciotti, R.; Gatto, V.; Gardini, F.; Torriani, S. Tyrosine decarboxylase activity of enterococci grown in media with different nutritional potential: Tyramine and 2-phenylethylamine accumulation and tyrDC gene expression. Front. Microbiol. 2015, 6, e00259. [Google Scholar] [CrossRef] [PubMed]
- Doeun, D.; Davaatseren, M.; Chung, M.S. Biogenic amines in foods. Food Sci. Biotechnol. 2017, 26, 1463–1474. [Google Scholar] [CrossRef]
- Tabanelli, G.; Montanari, C.; Gardini, F. Biogenic amines in food: A review of factors affecting their formation. In Encyclopedia of Food Chemistry; Melton, L., Shahidi, F., Varelis, P., Eds.; Academic Press: New York, NY, USA, 2019; pp. 337–343. [Google Scholar]
- Gardini, F.; Martuscelli, M.; Caruso, M.C.; Galgano, F.; Crudele, M.A.; Favati, F.; Guerzoni, M.E.; Suzzi, G. Effects of pH, temperature and NaCl concentration on the growth kinetics, proteolytic activity and biogenic amine production of Enterococcus faecalis. Int. J. Food Microbiol. 2001, 64, 105–117. [Google Scholar] [CrossRef]
- Pérez-Díaz, I.M.; Hayes, J.S.; Medina, E.; Weber, A.M.; Butz, N.; Dickey, A.N.; Lu, Z.; Azcarate-Peril, M.A. Assessment of the non-lactic acid bacteria microbiota in fresh cucumbers and commercially fermented cucumber pickles brined with 6% NaCl. Food Microbiol. 2019, 77, 10–20. [Google Scholar] [CrossRef]
- Taale, E.; Savadogo, A.; Zongo, C.; Ilboudo, A.J.; Traore, A.S. Bioactive molecules from bacteria strains: Case of bacteriocins producing bacteria isolated from foods. Curr. Res. Microbiol. Biotechnol. 2013, 1, 80–88. [Google Scholar]
- Franco, W.; Pérez-Díaz, I.M. Role of selected oxidative yeasts and bacteria in cucumber secondary fermentation associated with spoilage of the fermented fruit. Food Microbiol. 2012, 32, 338–344. [Google Scholar] [CrossRef]
- Baylis, C.L. Enterobacteriaceae. In Food Spoilage Microorganisms; de Blackburn, C.W., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2006; pp. 624–667. [Google Scholar]
- Paarup, T.; Nieto, J.C.; Peláez, C.; Reguera, J.I. Microbiological and physico-chemical characterisation of deep spoilage in Spanish dry-cured hams and characterisation of isolated Enterobacteriaceae with regard to salt and temperature tolerance. Eur. Food Res. Technol. 1999, 209, 366–371. [Google Scholar] [CrossRef]
- Paramithiotis, S.; Hondrodimou, O.L.; Drosinos, E.H. Development of the microbial community during spontaneous cauliflower fermentation. Food Res. Int. 2010, 43, 1098–1103. [Google Scholar] [CrossRef]
- Greif, G.; Greifová, M.; Karovičová, J. Effects of NaCl concentration and initial pH value on biogenic amine formation dynamics by Enterobacter spp. bacteria in model conditions. J. Food Nutr. Res. 2006, 45, 21–29. [Google Scholar]
- Marino, M.; Maifreni, M.; Moret, S.; Rondinini, G. The capacity of Enterobacteriaceae species to produce biogenic amines in cheese. Lett. Appl. Microbiol. 2000, 31, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Hu, W.; Jiang, A.; Xiu, Z.; Ji, Y.; Guan, Y.; Serengaowa, Y.X. Effect of salt concentration on quality of northeast sauerkraut fermented by Leuconostoc mesenteroides and Lactobacillus plantarum. Food Biosci. 2019, 30, e100421. [Google Scholar] [CrossRef]
- M’hir, S.; Minervini, F.; Di Cagno, R.; Chammem, N.; Hamdi, M. Technological, functional and safety aspects of enterococci in fermented vegetable products: A mini-review. Ann. Microbiol. 2012, 62, 469–481. [Google Scholar] [CrossRef]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The Genus Enterococcus: Between Probiotic Potential and Safety Concerns—An Update. Front. Microbiol. 2018, 9, e01791. [Google Scholar] [CrossRef] [PubMed]
- Etchells, J.L. A New Type of Gaseous Fermentation Occurring during the Salting of Cucumbers. Ph.D. Thesis, Michigan State University, Ann Arbor, MI, USA, 1941. [Google Scholar]
- Franco, W.; Pérez-Díaz, I.M.; Johanningsmeier, S.D.; McFeeters, R.F. Characteristics of spoilage-associated secondary cucumber fermentation. Appl. Environ. Microbiol. 2012, 78, 1273–1284. [Google Scholar] [CrossRef] [Green Version]
- Stoll, D.A.; Müller, A.; Meinhardt, A.K.; Dötsch, A.; Greiner, R.; Kulling, S.E.; Huch, M. Influence of salt concentration and iodized table salt on the microbiota of fermented cucumbers. Food Microbiol. 2020, 92, 103552. [Google Scholar] [CrossRef]
- Moret, S.; Smela, D.; Populin, T.; Conte, L.S. A survey on free biogenic amine content of fresh and preserved vegetables. Food Chem. 2005, 89, 355–361. [Google Scholar] [CrossRef]
- Simon Sarkadi, L. Biogenic amines in fermented foods and health implications. In Fermented Foods in Health and Disease Prevention; Frias, J., Martinez-Villaluenga, C., Peñas, E., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 625–648. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef] [Green Version]
- Galgano, F.; Caruso, M.; Condelli, N.; Favati, F. Focused Review: Agmatine in Fermented Foods. Front. Microbiol. 2012, 3, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halász, A.; Baráth, Á.; Holzapfel, W.H. The influence of starter culture selection on sauerkraut fermentation. Z. Für Lebensm. Und-Forsch. A 1999, 208, 434–438. [Google Scholar] [CrossRef]
- Świder, O.; Roszko, M.Ł.; Wójcicki, M.; Szymczyk, K. Biogenic amines and free amino acids in traditional fermented vegetables -dietary risk evaluation. J. Agric. Food Chem. 2020, 68, 856–868. [Google Scholar] [CrossRef]
- Rabie, M.A.; Siliha, H.; el-Saidy, S.; el-Badawy, A.A.; Malcata, F.X. Reduced biogenic amine content in sauerkraut via addition of selected lactic acid bacteria. Food Chem. 2011, 129, 1778–1782. [Google Scholar] [CrossRef]
- Morii, H.; Kasama, K. Activity of Two Histidine Decarboxylases from Photobacterium phosphoreum at Different Temperatures, pHs, and NaCl Concentrations. J. Food Protect. 2004, 67, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Pérez-Díaz, I.M.; Breidt, F.; Hayes, J.; Franco, W.; Butz, N.; Azcarate-Peril, M.A. Bacterial Ecology of Fermented Cucumber Rising pH Spoilage as Determined by Nonculture-Based Methods. J. Food Sci. 2016, 81, M121–M129. [Google Scholar] [CrossRef] [PubMed]
- McMurtrie, E.K.; Johanningsmeier, S.D.; Breidt, F., Jr.; Price, R.E. Effect of Brine Acidification on Fermentation Microbiota, Chemistry, and Texture Quality of Cucumbers Fermented in Calcium or Sodium Chloride Brines. J. Food Sci. 2019, 84, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Pickles. Available online: http://www.fao.org/3/a-au116e.pdf (accessed on 21 August 2021).
- Behera, S.S.; El Sheikha, A.F.; Hammami, R.; Kumar, A. Traditionally fermented pickles: How the microbial diversity associated with their nutritional and health benefits? J. Funct. Foods 2020, 70, 103971. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Świder, O.; Wójcicki, M.; Bujak, M.; Juszczuk-Kubiak, E.; Szczepańska, M.; Roszko, M.Ł. Time Evolution of Microbial Composition and Metabolic Profile for Biogenic Amines and Free Amino Acids in a Model Cucumber Fermentation System Brined with 0.5% to 5.0% Sodium Chloride. Molecules 2021, 26, 5796. https://doi.org/10.3390/molecules26195796
Świder O, Wójcicki M, Bujak M, Juszczuk-Kubiak E, Szczepańska M, Roszko MŁ. Time Evolution of Microbial Composition and Metabolic Profile for Biogenic Amines and Free Amino Acids in a Model Cucumber Fermentation System Brined with 0.5% to 5.0% Sodium Chloride. Molecules. 2021; 26(19):5796. https://doi.org/10.3390/molecules26195796
Chicago/Turabian StyleŚwider, Olga, Michał Wójcicki, Marzena Bujak, Edyta Juszczuk-Kubiak, Magdalena Szczepańska, and Marek Ł. Roszko. 2021. "Time Evolution of Microbial Composition and Metabolic Profile for Biogenic Amines and Free Amino Acids in a Model Cucumber Fermentation System Brined with 0.5% to 5.0% Sodium Chloride" Molecules 26, no. 19: 5796. https://doi.org/10.3390/molecules26195796