
Antioxidant Effects of Anthocyanin-Rich Riceberry™ Rice Flour Prepared Using Dielectric Barrier Discharge Plasma Technology on Iron-Induced Oxidative Stress in Mice

 , and
, and 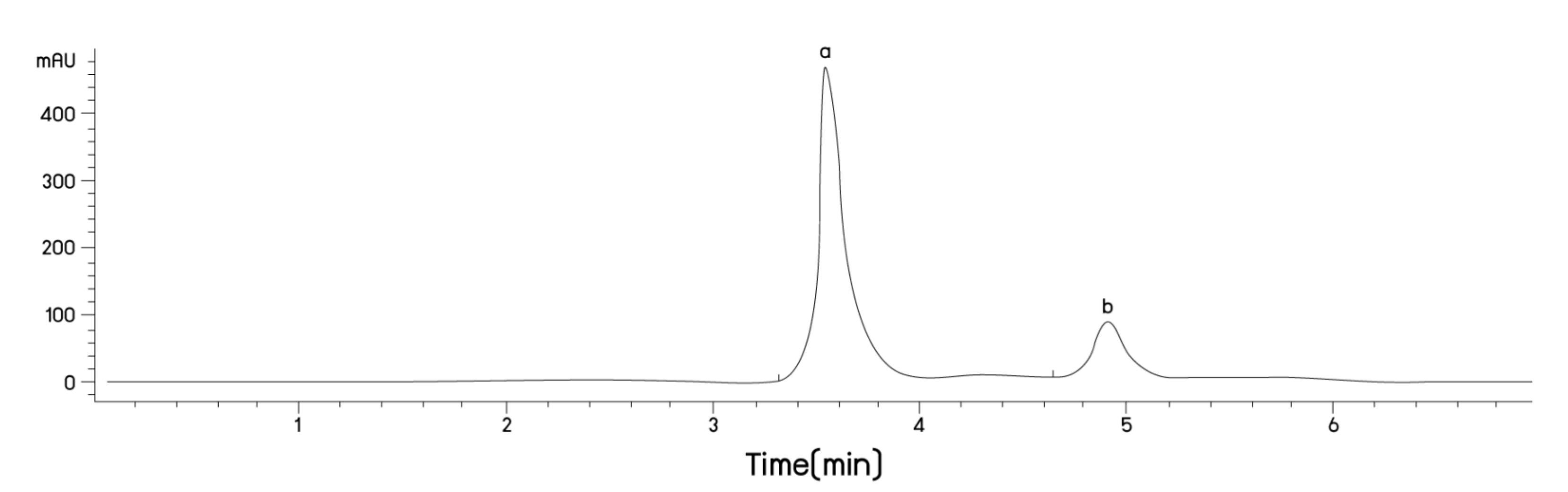{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Anthocyanin Content of Riceberry™ Rice Flour (PRBF) Using Plasma Technology
2.2. Effect of PRBF Treatment on BW and OWI
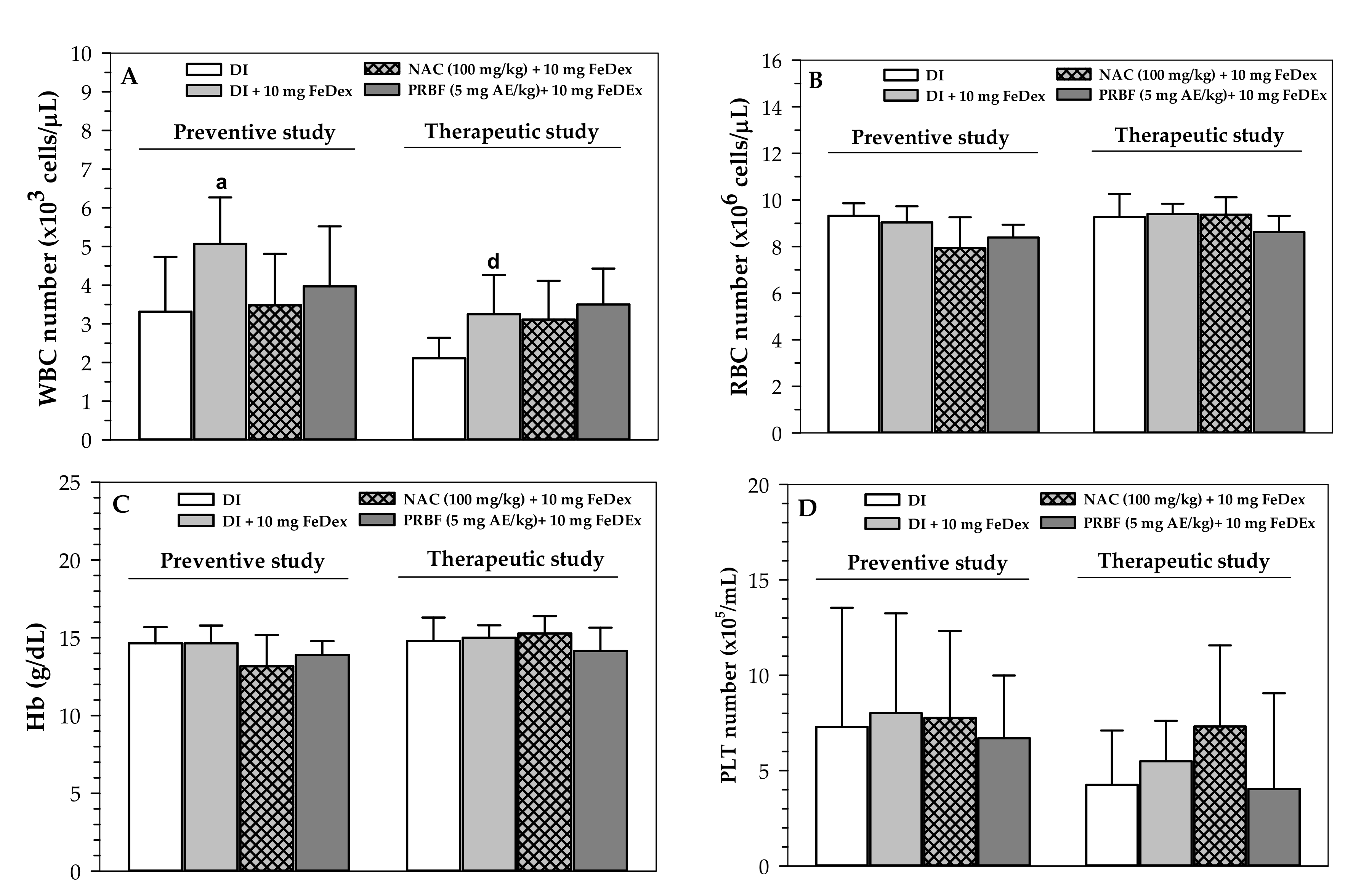
2.3. Effect of PRBF Treatment on Hematopoiesis
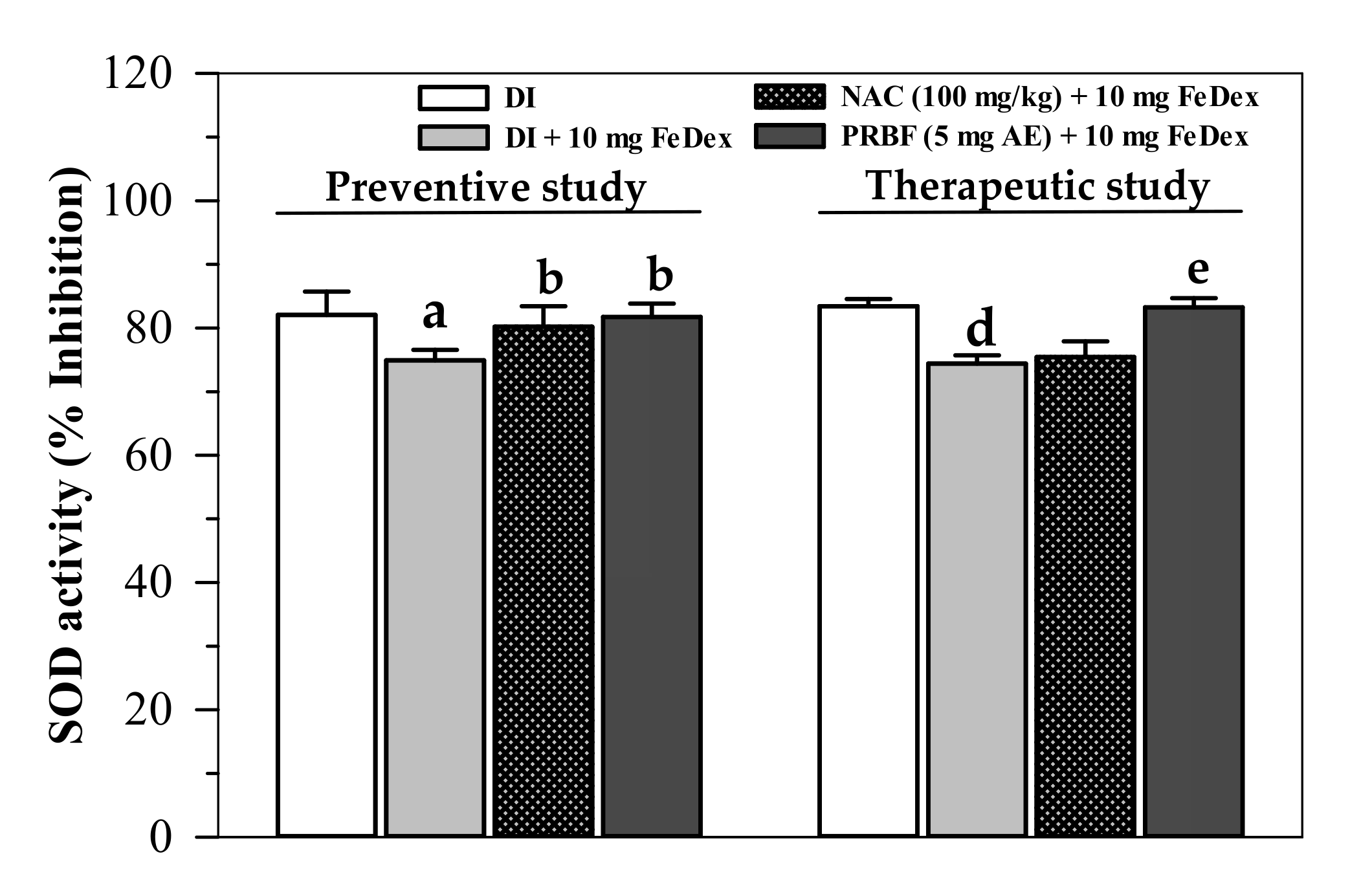
2.4. Effect of PRBF Treatment on Erythrocyte SOD Activity
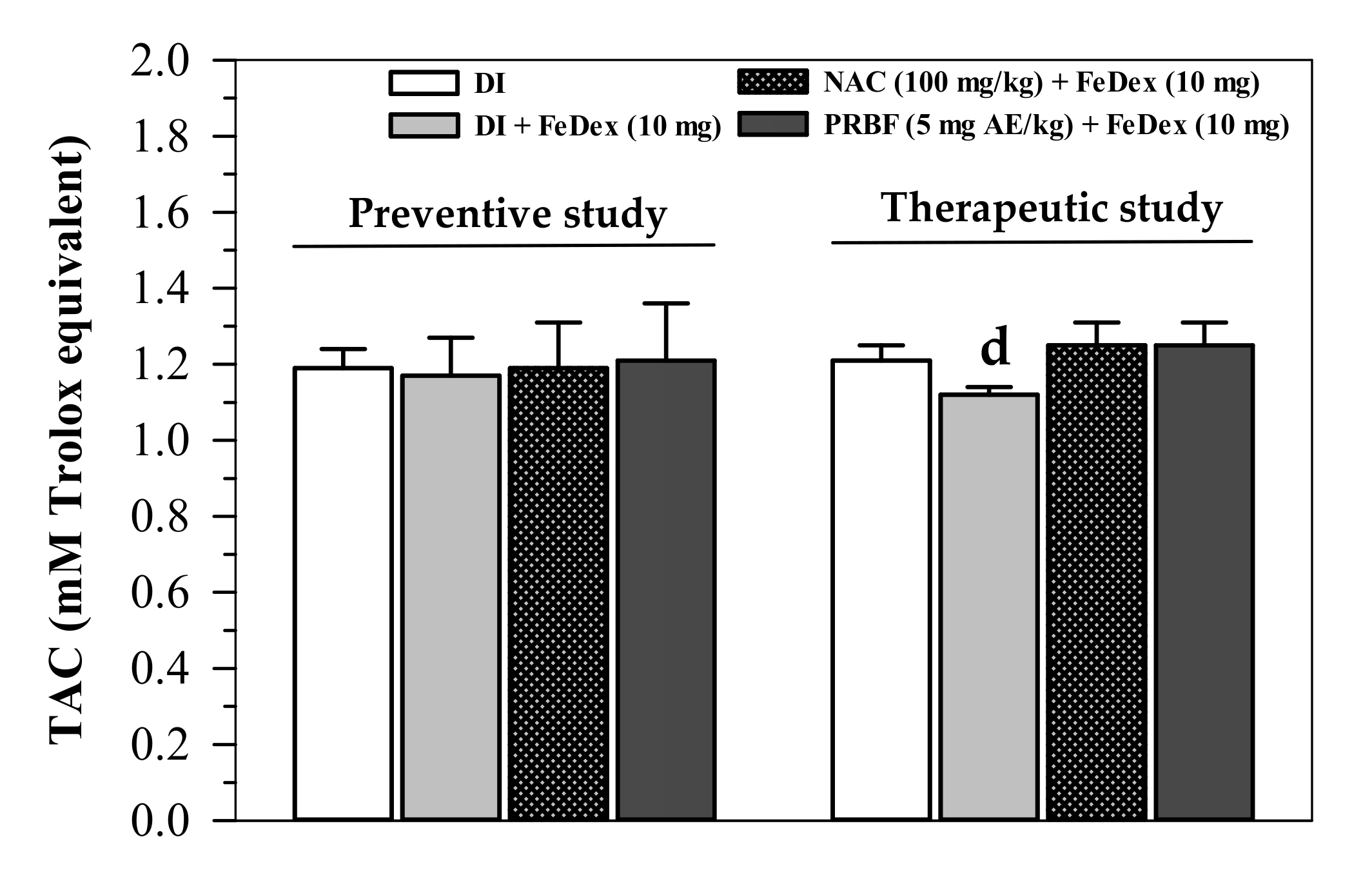
2.5. Effect of PRBF on Plasma TAC
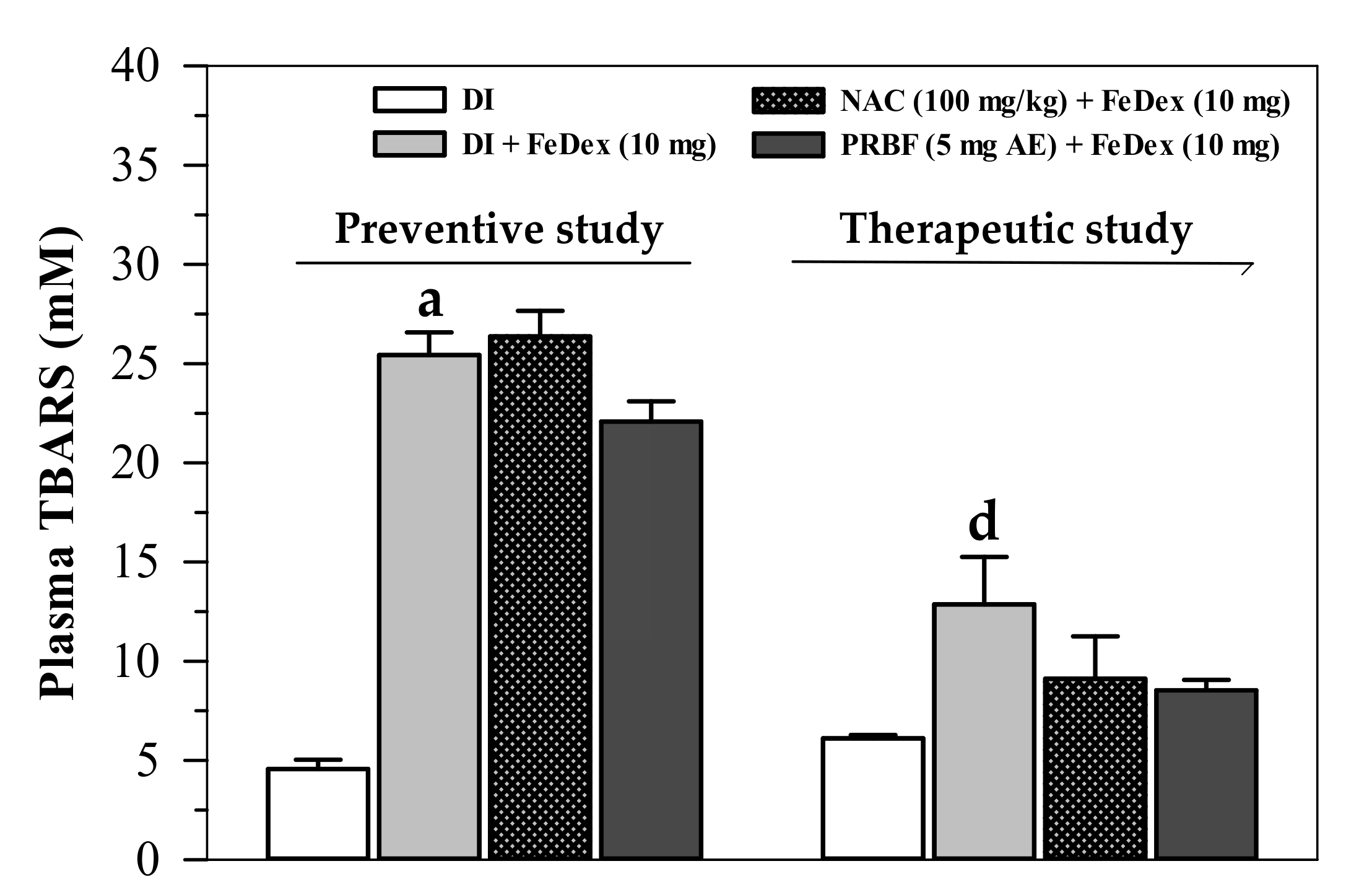
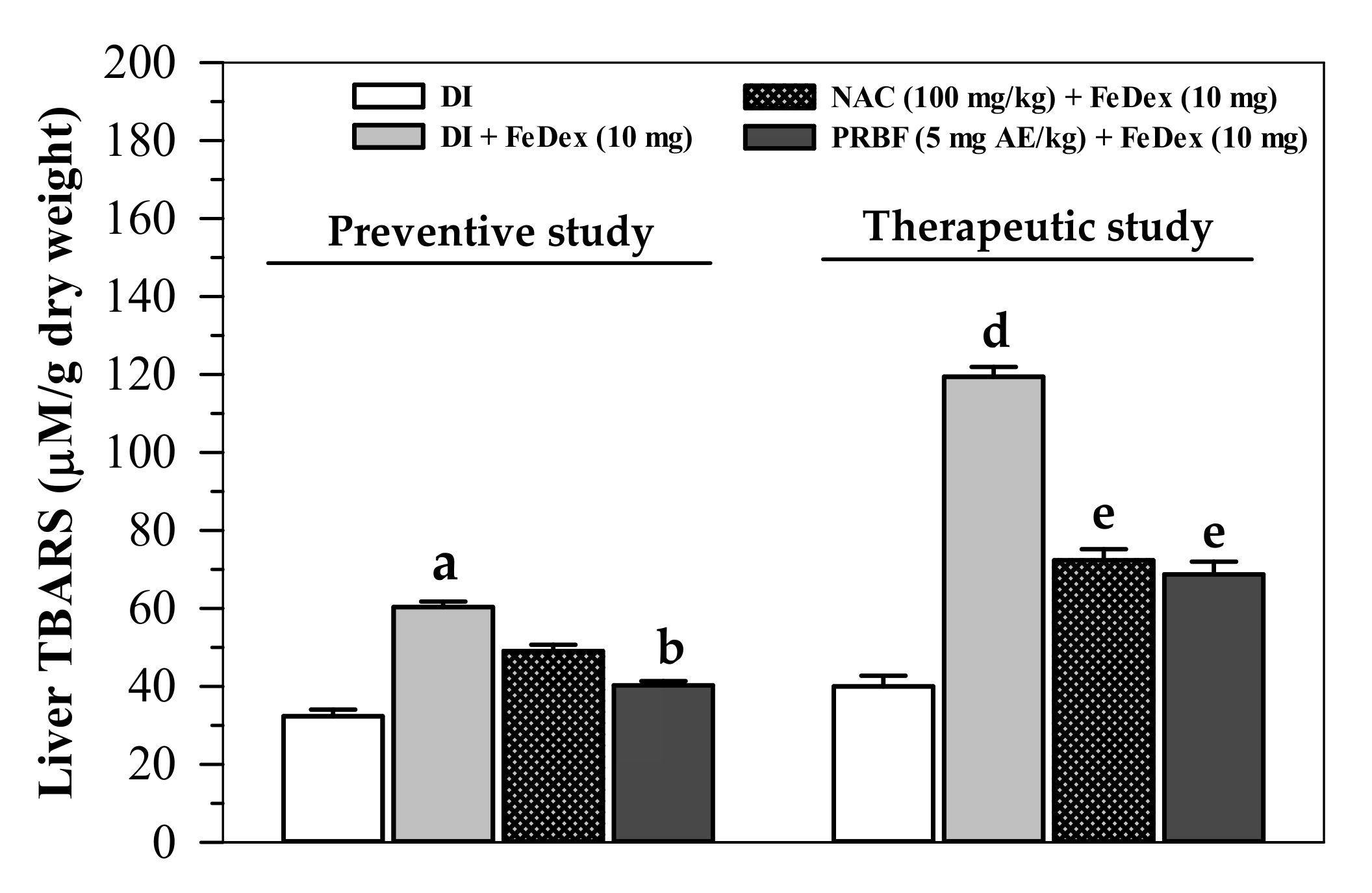
2.6. Effect of PRBF on Plasma and Liver TBARS
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Methods
4.2.1. Production of Riceberry Rice Flour Using Plasma Technology
4.2.2. HPLC Analysis of Anthocyanin Content
4.2.3. Animal Care
4.2.4. Iron Loading and PRBF Treatment in Mice
4.2.5. Hematological Parameter Analysis
4.2.6. SOD Activity Assay
4.2.7. Determination of Plasma Antioxidant Capacity
4.2.8. Quantification of Plasma and Liver TBARS
4.2.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burton, G.J.; Jauniaux, E. Oxidative stress. Best Pract. Res. Clin. Obstet. Gynaecol. 2011, 25, 287–299. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxidative Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef]
- Young, I.S.; Woodside, J.V. Antioxidants in health and disease. J. Clin. Pathol. 2001, 54, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lee, H.L.; Jung, W.T.; Sung, F.C.; Su, T.C. The association between urinary levels of 1,3-butadiene metabolites, cardiovascular risk factors, microparticles, and oxidative stress products in adolescents and young adults. J. Hazard. Mater. 2020, 396, 122745. [Google Scholar] [CrossRef]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Poljsak, B. Strategies for reducing or preventing the generation of oxidative stress. Oxidative Med. Cell. Longev. 2011, 2011, 194586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonyosying, P.; Tantiworawit, A.; Hantrakool, S.; Utama-Ang, N.; Cresswell, M.; Fucharoen, S.; Porter, J.B.; Srichairatanakool, S. Consumption of a green tea extract-curcumin drink decreases blood urea nitrogen and redox iron in beta-thalassemia patients. Food Funct. 2020, 11, 932–943. [Google Scholar] [CrossRef]
- Kalpravidh, R.W.; Siritanaratkul, N.; Insain, P.; Charoensakdi, R.; Panichkul, N.; Hatairaktham, S.; Srichairatanakool, S.; Phisalaphong, C.; Rachmilewitz, E.; Fucharoen, S. Improvement in oxidative stress and antioxidant parameters in beta-thalassemia/Hb E patients treated with curcuminoids. Clin. Biochem. 2009, 43, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Koonyosying, P.; Kongkarnka, S.; Uthaipibull, C.; Svasti, S.; Fucharoen, S.; Srichairatanakool, S. Green tea extract modulates oxidative tissue injury in beta-thalassemic mice by chelation of redox iron and inhibition of lipid peroxidation. Biomed. Pharm. 2018, 108, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Koonyosying, P.; Uthaipibull, C.; Fucharoen, S.; Koumoutsea, E.V.; Porter, J.B.; Srichairatanakool, S. Decrement in Cellular Iron and Reactive Oxygen Species, and Improvement of Insulin Secretion in a Pancreatic Cell Line Using Green Tea Extract. Pancreas 2019, 48, 636–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ounjaijean, S.; Thephinlap, C.; Khansuwan, U.; Phisalapong, C.; Fucharoen, S.; Porter, J.B.; Srichairatanakool, S. Effect of green tea on iron status and oxidative stress in iron-loaded rats. Med. Chem. 2008, 4, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Saewong, T.; Ounjaijean, S.; Mundee, Y.; Pattanapanyasat, K.; Fucharoen, S.; Porter, J.B.; Srichairatanakool, S. Effects of green tea on iron accumulation and oxidative stress in livers of iron-challenged thalassemic mice. Med. Chem. 2010, 6, 57–64. [Google Scholar] [CrossRef]
- Srichairatanakool, S.; Thephinlap, C.; Phisalaphong, C.; Porter, J.B.; Fucharoen, S. Curcumin contributes to in vitro removal of non-transferrin bound iron by deferiprone and desferrioxamine in thalassemic plasma. Med. Chem. 2007, 3, 469–474. [Google Scholar] [CrossRef]
- Thephinlap, C.; Phisalaphong, C.; Fucharoen, S.; Porter, J.B.; Srichairatanakool, S. Efficacy of curcuminoids in alleviation of iron overload and lipid peroxidation in thalassemic mice. Med. Chem. 2009, 5, 474–482. [Google Scholar] [CrossRef]
- Thephinlap, C.; Phisalaphong, C.; Lailerd, N.; Chattipakorn, N.; Winichagoon, P.; Vadolas, J.; Fucharoen, S.; Porter, J.B.; Srichairatanakool, S. Reversal of cardiac iron loading and dysfunction in thalassemic mice by curcuminoids. Med. Chem. 2011, 7, 62–69. [Google Scholar] [CrossRef]
- Yanpanitch, O.U.; Hatairaktham, S.; Charoensakdi, R.; Panichkul, N.; Fucharoen, S.; Srichairatanakool, S.; Siritanaratkul, N.; Kalpravidh, R.W. Treatment of beta-Thalassemia/Hemoglobin E with Antioxidant Cocktails Results in Decreased Oxidative Stress, Increased Hemoglobin Concentration, and Improvement of the Hypercoagulable State. Oxidative Med. Cell. Longev. 2015, 2015. [Google Scholar] [CrossRef] [Green Version]
- Chepelev, N.L.; Zhang, H.; Liu, H.; McBride, S.; Seal, A.J.; Morgan, T.E.; Finch, C.E.; Willmore, W.G.; Davies, K.J.A.; Forman, H.J. Competition of nuclear factor-erythroid 2 factors related transcription factor isoforms, Nrf1 and Nrf2, in antioxidant enzyme induction. Redox Biol. 2013, 1, 183–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannino, G.; Gentile, C.; Ertani, A.; Serio, G.; Bertea, C.M. Anthocyanins: Biosynthesis, Distribution, Ecological Role, and Use of Biostimulants to Increase Their Content in Plant Foods—A Review. Agriculture 2021, 11, 212. [Google Scholar] [CrossRef]
- Abdel-Aal, E.M.; Hucl, P.; Rabalski, I. Compositional and antioxidant properties of anthocyanin-rich products prepared from purple wheat. Food Chem. 2018, 254, 13–19. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.G.; Brownmiller, C.R.; Lee, S.O.; Kang, H.W. Anti-Inflammatory and Antioxidant Effects of Anthocyanins of Trifolium pratense (Red Clover) in Lipopolysaccharide-Stimulated RAW-267.4 Macrophages. Nutrients 2020, 12, 1089. [Google Scholar] [CrossRef] [Green Version]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and Human Health—A Focus on Oxidative Stress, Inflammation and Disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Van Hung, P. Phenolic Compounds of Cereals and Their Antioxidant Capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef]
- Xiang, J.; Apea-Bah, F.B.; Ndolo, V.U.; Katundu, M.C.; Beta, T. Profile of phenolic compounds and antioxidant activity of finger millet varieties. Food Chem. 2019, 275, 361–368. [Google Scholar] [CrossRef]
- Settapramote, N.; Laokuldilok, T.; Boonyawan, D.; Utama-ang, N. Physiochemical, Antioxidant Activities and Anthocyanin of Riceberry Rice from Different Locations in Thailand. Food Appl. Biosci. J. 2018, 6, 84–94. [Google Scholar]
- Thiranusornkij, L.; Thamnarathip, P.; Chandrachai, A.; Kuakpetoon, D.; Adisakwattana, S. Comparative studies on physicochemical properties, starch hydrolysis, predicted glycemic index of Hom Mali rice and Riceberry rice flour and their applications in bread. Food Chem. 2019, 283, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Rakkhumkaew, N.; Boonsri, Y.; Sukchum, A. Utilization of small broken riceberry flour in gluten-free bread. Food Sci. Technol. Int. 2019, 25, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Phuapaiboon, P. Gamma-aminobutyric acid, total anthocyanin content and antioxidant activity of vinegar brewed from germinated pigmented rice. Pak. J. Nutr. 2017, 16, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Sangngern, N.; Puangnark, T.; Nguansangiam, W.; Saithong, P.; Kitpreechavanich, V.; Lomthong, T. Production and development of vinegar fermentation from broken Riceberry rice using raw starch-degrading enzyme hydrolysis. Biotech 2020, 10, 515. [Google Scholar] [CrossRef]
- Suttiarporn, P.; Chumpolsri, W.; Mahatheeranont, S.; Luangkamin, S.; Teepsawang, S.; Leardkamolkarn, V. Structures of phytosterols and triterpenoids with potential anti-cancer activity in bran of black non-glutinous rice. Nutrients 2015, 7, 1672–1687. [Google Scholar] [CrossRef] [Green Version]
- Rodboon, T.; Okada, S.; Suwannalert, P. Germinated Riceberry Rice Enhanced Protocatechuic Acid and Vanillic Acid to Suppress Melanogenesis through Cellular Oxidant-Related Tyrosinase Activity in B16 Cells. Antioxidants 2020, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Settapramote, N.; Laokuldilok, T.; Boonyawan, D.; Utama-ang, N. Optimisation of the dielectric barrier discharge to produce Riceberry rice flour retained with high activities of bioactive compounds using plasma technology. Int. Food Res. J. 2021, 28, 386–392. [Google Scholar]
- Bernhardt, T.; Semmler, M.L.; Schäfer, M.; Bekeschus, S.; Emmert, S.; Boeckmann, L. Plasma Medicine: Applications of Cold Atmospheric Pressure Plasma in Dermatology. Oxidative Med. Cell. Longev. 2019, 2019, 3873928. [Google Scholar] [CrossRef] [Green Version]
- Jain, V.; Visani, A.; Srinivasan, R.; Agarwal, V. Design and development of a low cost, high current density power supply for streamer free atmospheric pressure DBD plasma generation in air. Rev. Sci. Instrum. 2018, 89, 033502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahrami, N.; Bayliss, D.; Chope, G.; Penson, S.; Perehinec, T.; Fisk, I.D. Cold plasma: A new technology to modify wheat flour functionality. Food Chem. 2016, 202, 247–253. [Google Scholar] [CrossRef]
- Pal, P.; Kaur, P.; Singh, N.; Kaur, A.; Misra, N.N.; Tiwari, B.K.; Cullen, P.J.; Virdi, A.S. Effect of nonthermal plasma on physico-chemical, amino acid composition, pasting and protein characteristics of short and long grain rice flour. Food Res. Int. 2016, 81, 50–57. [Google Scholar] [CrossRef]
- Misra, N.N.; Kaur, S.; Tiwari, B.K.; Kaur, A.; Singh, N.; Cullen, P.J. Atmospheric pressure cold plasma (ACP) treatment of wheat flour. Food Hydrocoll. 2015, 44, 115–121. [Google Scholar] [CrossRef]
- Dong, S.; Gao, A.; Xu, H.; Chen, Y. Effects of Dielectric Barrier Discharges (DBD) Cold Plasma Treatment on Physicochemical and Structural Properties of Zein Powders. Food Bioprocess. Technol. 2017, 10, 434–444. [Google Scholar] [CrossRef]
- Wongsagonsup, R.; Deeyai, P.; Chaiwat, W.; Horrungsiwat, S.; Leejariensuk, K.; Suphantharika, M.; Fuongfuchat, A.; Dangtip, S. Modification of tapioca starch by non-chemical route using jet atmospheric argon plasma. Carbohydr. Polym. 2014, 102, 790–798. [Google Scholar] [CrossRef] [PubMed]
- Scholtz, V.; Šerá, B.; Khun, J.; Šerý, M.; Julák, J. Effects of Nonthermal Plasma on Wheat Grains and Products. J. Food Qual. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, A.I.; Liao, X.; Cullen, P.J.; Liu, D.; Xiang, Q.; Wang, J.; Chen, S.; Ye, X.; Ding, T. Effects of nonthermal plasma technology on functional food components. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1379–1394. [Google Scholar] [CrossRef] [Green Version]
- Pankaj, S.K.; Wan, Z.; Keener, K.M. Effects of Cold Plasma on Food Quality: A Review. Foods 2018, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Yodpitak, S.; Mahatheeranont, S.; Boonyawan, D.; Sookwong, P.; Roytrakul, S.; Norkaew, O. Cold plasma treatment to improve germination and enhance the bioactive phytochemical content of germinated brown rice. Food Chem. 2019, 289, 328–339. [Google Scholar] [CrossRef]
- Suhem, K.; Matan, N.; Nisoa, M. Inhibition of Aspergillus flavus on agar media and brown rice cereal bars using cold atmospheric plasma treatment. Int. J. Food Microbiol. 2013, 161, 107–111. [Google Scholar] [CrossRef]
- Kang, J.H.; Bai, J.; Min, S.C. Inactivation of Indigenous Microorganisms and Salmonella in Korean Rice Cakes by In-Package Cold Plasma Treatment. Int. J. Environ. Res. Public Health 2021, 18, 3360. [Google Scholar] [CrossRef]
- Suttireung, P.; Winuprasith, T.; Srichamnong, W.; Paemuang, W.; Phonyiam, T.; Trachootham, D. Riceberry rice puddings: Rice-based low glycemic dysphagia diets. Asia Pac. J. Clin. Nutr. 2019, 28, 467–475. [Google Scholar] [PubMed]
- Laokuldilok, T.; Shoemaker, C.F.; Jongkaewwattana, S.; Tulyathan, V. Antioxidants and Antioxidant Activity of Several Pigmented Rice Brans. J. Agric. Food Chem. 2011, 59, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Peskin, A.V.; Winterbourn, C.C. A microtiter plate assay for superoxide dismutase using a water-soluble tetrazolium salt (WST-1). Clin. Chim. Acta 2000, 293, 157–166. [Google Scholar] [CrossRef]
- Deng, G.F.; Xu, X.R.; Zhang, Y.; Li, D.; Gan, R.Y.; Li, H.B. Phenolic compounds and bioactivities of pigmented rice. Crit. Rev. Food Sci. Nutr. 2012, 53, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.P.; Lai, H.M. Bioactive compounds and antioxidative activity of colored rice bran. J. Food Drug Anal. 2016, 24, 564–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, M.L.; Chen, P.N.; Chu, S.C.; Kuo, D.Y.; Kuo, W.H.; Chen, J.Y.; Hsieh, Y.S. Peonidin 3-glucoside inhibits lung cancer metastasis by downregulation of proteinases activities and MAPK pathway. Nutr. Cancer 2010, 62, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Yooin, W.; Saenjum, C. Investigation of Pigments in Thai Purple Rice Using Electron Paramagnetic Resonance Imaging and HPLC. J. Nutr. Sci. Vitam. 2019, 65, S217–S221. [Google Scholar] [CrossRef]
- Hu, C.; Zawistowski, J.; Ling, W.; Kitts, D.D. Black rice (Oryza sativa L. indica) pigmented fraction suppresses both reactive oxygen species and nitric oxide in chemical and biological model systems. J. Agric. Food Chem. 2003, 51, 5271–5277. [Google Scholar] [CrossRef]
- Goufo, P.; Trindade, H. Factors influencing antioxidant compounds in rice. Crit. Rev. Food Sci. Nutr. 2015, 57, 893–922. [Google Scholar] [CrossRef]
- Bhawamai, S.; Lin, S.H.; Hou, Y.Y.; Chen, Y.H. Thermal cooking changes the profile of phenolic compounds, but does not attenuate the anti-inflammatory activities of black rice. Food Nutr. Res. 2016, 60, 32941. [Google Scholar] [CrossRef]
- Peanparkdee, M.; Patrawart, J.; Iwamoto, S. Physicochemical stability and in vitro bioaccessibility of phenolic compounds and anthocyanins from Thai rice bran extracts. Food Chem. 2020, 329, 127157. [Google Scholar] [CrossRef]
- Chatthongpisut, R.; Schwartz, S.J.; Yongsawatdigul, J. Antioxidant activities and antiproliferative activity of Thai purple rice cooked by various methods on human colon cancer cells. Food Chem. 2015, 188, 99–105. [Google Scholar] [CrossRef]
- Li, F.; Chen, G.; Zhang, B.; Fu, X. Current applications and new opportunities for the thermal and non-thermal processing technologies to generate berry product or extracts with high nutraceutical contents. Food Res. Int. 2017, 100, 19–30. [Google Scholar] [CrossRef]
- Bursac Kovacevic, D.; Gajdos Kljusuric, J.; Putnik, P.; Vukusic, T.; Herceg, Z.; Dragovic-Uzelac, V. Stability of polyphenols in chokeberry juice treated with gas phase plasma. Food Chem. 2016, 212, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Niemira, B.A.; Gurtler, J.B.; Fan, X.; Sites, J.; Boyd, G.; Chen, H. Atmospheric cold plasma inactivation of aerobic microorganisms on blueberries and effects on quality attributes. Food Microbiol. 2014, 46, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, R.; Gan, Z.; Shao, T.; Zhang, X.; He, M.; Sun, A. Effect of cold plasma on blueberry juice quality. Food Chem. 2019, 290, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Bursac Kovacevic, D.; Putnik, P.; Dragovic-Uzelac, V.; Pedisic, S.; Rezek Jambrak, A.; Herceg, Z. Effects of cold atmospheric gas phase plasma on anthocyanins and color in pomegranate juice. Food Chem. 2015, 190, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Kotani, K.; Sakane, N. White Blood Cells, Neutrophils, and Reactive Oxygen Metabolites among Asymptomatic Subjects. Int. J. Prev. Med. 2012, 3, 428–431. [Google Scholar]
- Stroot, P.G. Blood oxidative stress (BLOS) is a secondary host defense system responding normally to anaerobic wound infection and inadvertently to dietary ultra-exogenous sulfide formation (USF). Med. Hypotheses 2017, 98, 28–34. [Google Scholar] [CrossRef]
- Kurhaluk, N.; Sliuta, A.; Kyriienko, S.; Winklewski, P.J. Melatonin Restores White Blood Cell Count, Diminishes Glycated Haemoglobin Level and Prevents Liver, Kidney and Muscle Oxidative Stress in Mice Exposed to Acute Ethanol Intoxication. Alcohol Alcohol. 2017, 52, 521–528. [Google Scholar] [CrossRef]
- Schümann, K.; Kroll, S.; Weiss, G.; Frank, J.; Biesalski, H.K.; Daniel, H.; Friel, J.; Solomons, N.W. Monitoring of hematological, inflammatory and oxidative reactions to acute oral iron exposure in human volunteers: Preliminary screening for selection of potentially-responsive biomarkers. Toxicology 2005, 212, 10–23. [Google Scholar] [CrossRef]
- Li, J.; Lei, J.; He, L.; Fan, X.; Yi, F.; Zhang, W. Evaluation and Monitoring of Superoxide Dismutase (SOD) Activity and its Clinical Significance in Gastric Cancer: A Systematic Review and Meta-Analysis. Med. Sci. Monit. 2019, 25, 2032–2042. [Google Scholar] [CrossRef]
- Sies, H. Total antioxidant capacity: Appraisal of a concept. J. Nutr. 2007, 137, 1493–1495. [Google Scholar] [CrossRef]
- Young, I.S. Measurement of total antioxidant capacity. J. Clin. Pathol. 2001, 54, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.R.; Shah, Y.M. Iron homeostasis in the liver. Compr. Physiol. 2013, 3, 315–330. [Google Scholar] [PubMed] [Green Version]
- Deugnier, Y.; Turlin, B. Pathology of hepatic iron overload. World J. Gastroenterol. 2007, 13, 4755–4760. [Google Scholar] [CrossRef]
- Anuyahong, T.; Chusak, C.; Thilavech, T.; Adisakwattana, S. Postprandial Effect of Yogurt Enriched with Anthocyanins from Riceberry Rice on Glycemic Response and Antioxidant Capacity in Healthy Adults. Nutrients 2020, 12, 2930. [Google Scholar] [CrossRef] [PubMed]
- Chusak, C.; Pasukamonset, P.; Chantarasinlapin, P.; Adisakwattana, S. Postprandial Glycemia, Insulinemia, and Antioxidant Status in Healthy Subjects after Ingestion of Bread made from Anthocyanin-Rich Riceberry Rice. Nutrients 2020, 12, 782. [Google Scholar] [CrossRef] [Green Version]
- Hou, Z.; Qin, P.; Ren, G. Effect of anthocyanin-rich extract from black rice (Oryza sativa L. Japonica) on chronically alcohol-induced liver damage in rats. J. Agric. Food Chem. 2012, 58, 3191–3196. [Google Scholar] [CrossRef]
- Wu, T.; Guo, X.; Zhang, M.; Yang, L.; Liu, R.; Yin, J. Anthocyanins in black rice, soybean and purple corn increase fecal butyric acid and prevent liver inflammation in high fat diet-induced obese mice. Food Funct. 2017, 8, 3178–3186. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ling, W.; Wang, Q.; Liu, C.; Hu, Y.; Xia, M.; Feng, X.; Xia, X. Effect of anthocyanin-rich extract from black rice (Oryza sativa L. indica) on hyperlipidemia and insulin resistance in fructose-fed rats. Plant. Foods Hum. Nutr. 2007, 62, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Arjinajarn, P.; Chueakula, N.; Pongchaidecha, A.; Jaikumkao, K.; Chatsudthipong, V.; Mahatheeranont, S.; Norkaew, O.; Chattipakorn, N.; Lungkaphin, A. Anthocyanin-rich Riceberry bran extract attenuates gentamicin-induced hepatotoxicity by reducing oxidative stress, inflammation and apoptosis in rats. Biomed. Pharm. 2017, 92, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Itani, T.; Morimitsu, Y.; Okada, K.; Ozeki, M.; Kondo, S.; Uchida, K.; Osawa, T.; Hiai, H.; Tashiro, T. Protective effect of colored rice over white rice on Fenton reaction-based renal lipid peroxidation in rats. Free Radic. Res. 2002, 36, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Cásedas, G.; González-Burgos, E.; Smith, C.; López, V.; Gómez-Serranillos, M.P. Regulation of redox status in neuronal SH-SY5Y cells by blueberry (Vaccinium myrtillus L.) juice, cranberry (Vaccinium macrocarpon A.) juice and cyanidin. Food Chem. Toxicol. 2018, 118, 572–580. [Google Scholar] [CrossRef]
- Deng, J.; Cheng, J.; Liao, X.; Zhang, T.; Leng, X.; Zhao, G. Comparative study on iron release from soybean (Glycine max) seed ferritin induced by anthocyanins and ascorbate. J. Agric. Food Chem. 2010, 58, 635–641. [Google Scholar] [CrossRef]
Sample Availability: Not available. |






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Settapramote, N.; Utama-ang, N.; Petiwathayakorn, T.; Settakorn, K.; Svasti, S.; Srichairatanakool, S.; Koonyosying, P. Antioxidant Effects of Anthocyanin-Rich Riceberry™ Rice Flour Prepared Using Dielectric Barrier Discharge Plasma Technology on Iron-Induced Oxidative Stress in Mice. Molecules 2021, 26, 4978. https://doi.org/10.3390/molecules26164978
Settapramote N, Utama-ang N, Petiwathayakorn T, Settakorn K, Svasti S, Srichairatanakool S, Koonyosying P. Antioxidant Effects of Anthocyanin-Rich Riceberry™ Rice Flour Prepared Using Dielectric Barrier Discharge Plasma Technology on Iron-Induced Oxidative Stress in Mice. Molecules. 2021; 26(16):4978. https://doi.org/10.3390/molecules26164978
Chicago/Turabian StyleSettapramote, Natwalinkhol, Niramon Utama-ang, Touchwin Petiwathayakorn, Kornvipa Settakorn, Saovaros Svasti, Somdet Srichairatanakool, and Pimpisid Koonyosying. 2021. "Antioxidant Effects of Anthocyanin-Rich Riceberry™ Rice Flour Prepared Using Dielectric Barrier Discharge Plasma Technology on Iron-Induced Oxidative Stress in Mice" Molecules 26, no. 16: 4978. https://doi.org/10.3390/molecules26164978