
Effect of Sodium Valproate on the Conformational Stability of the Visual G Protein-Coupled Receptor Rhodopsin

 , , and
, , and
Abstract
:1. Introduction
2. Results
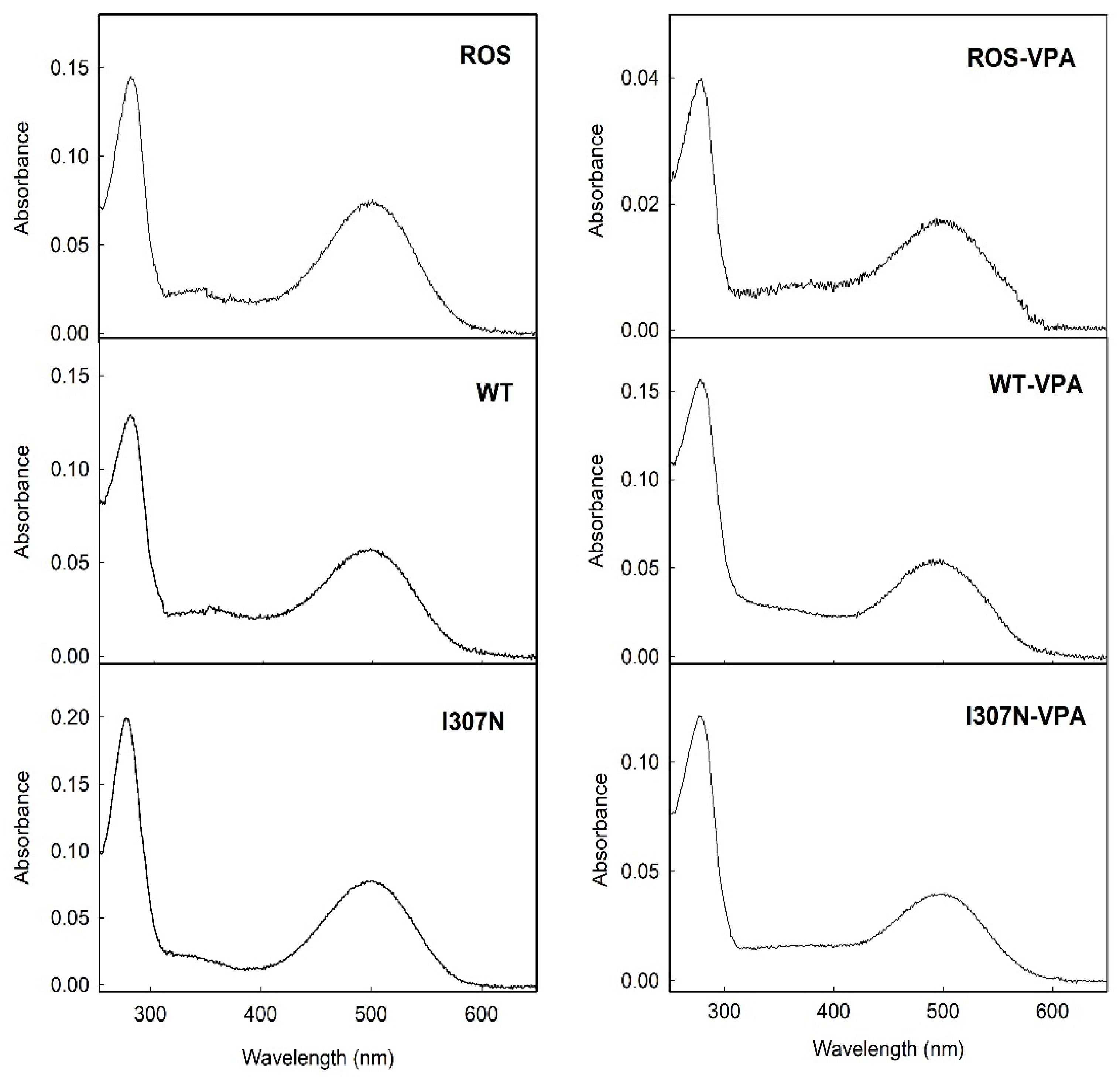
2.1. UV-Vis Spectroscopic Characterization.
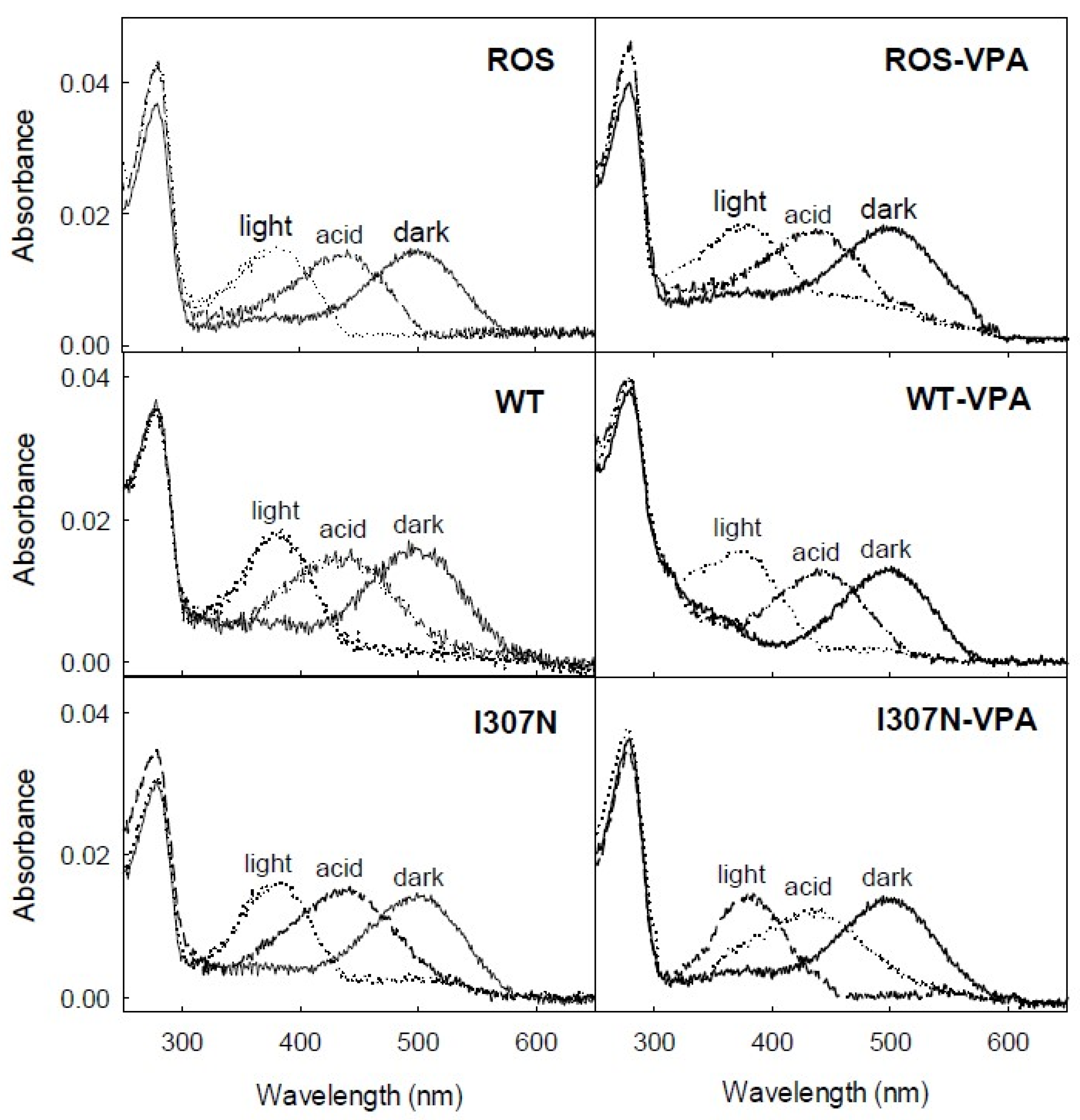
2.2. Photobleaching and Acidification
2.3. Thermal Stability
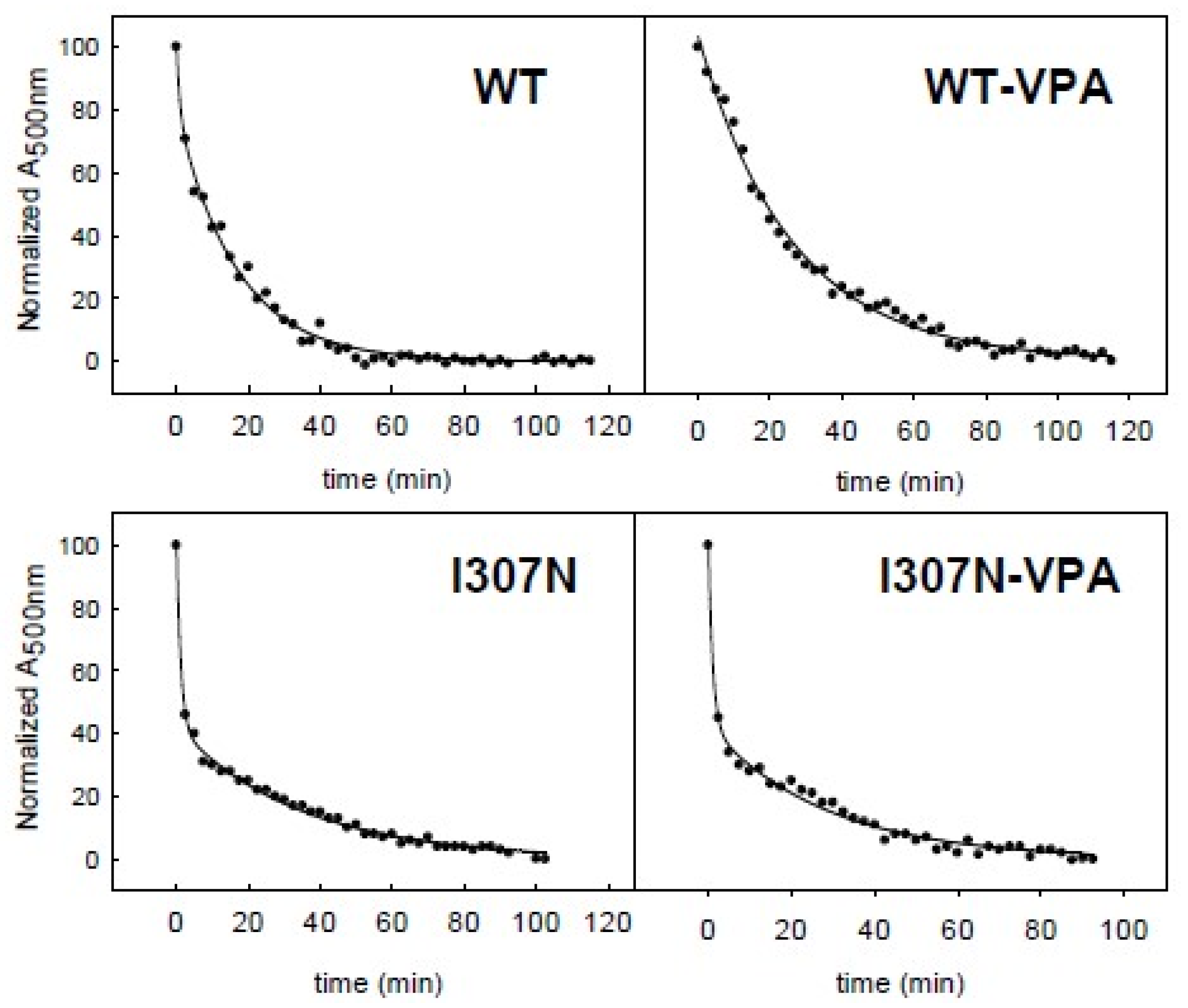
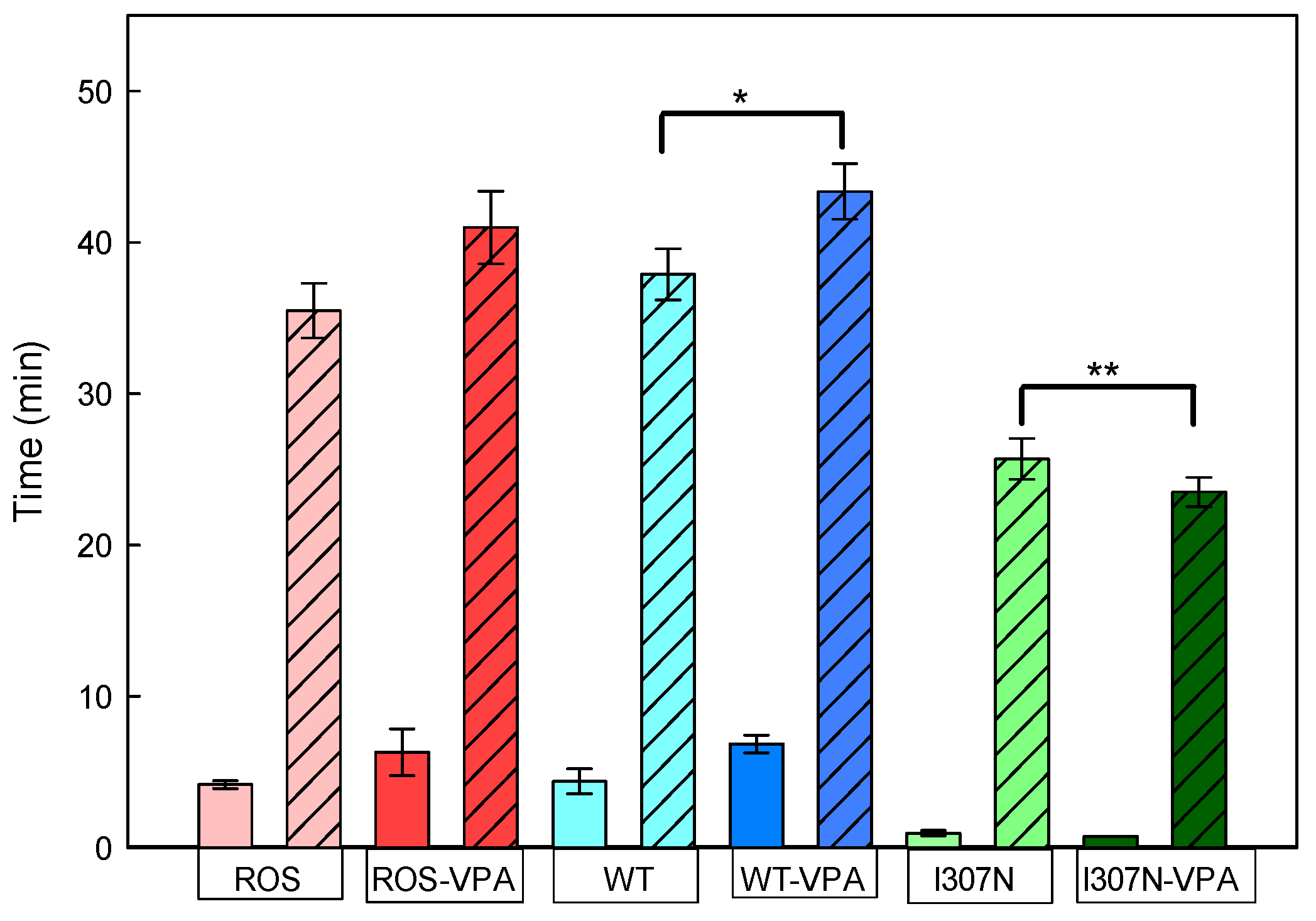
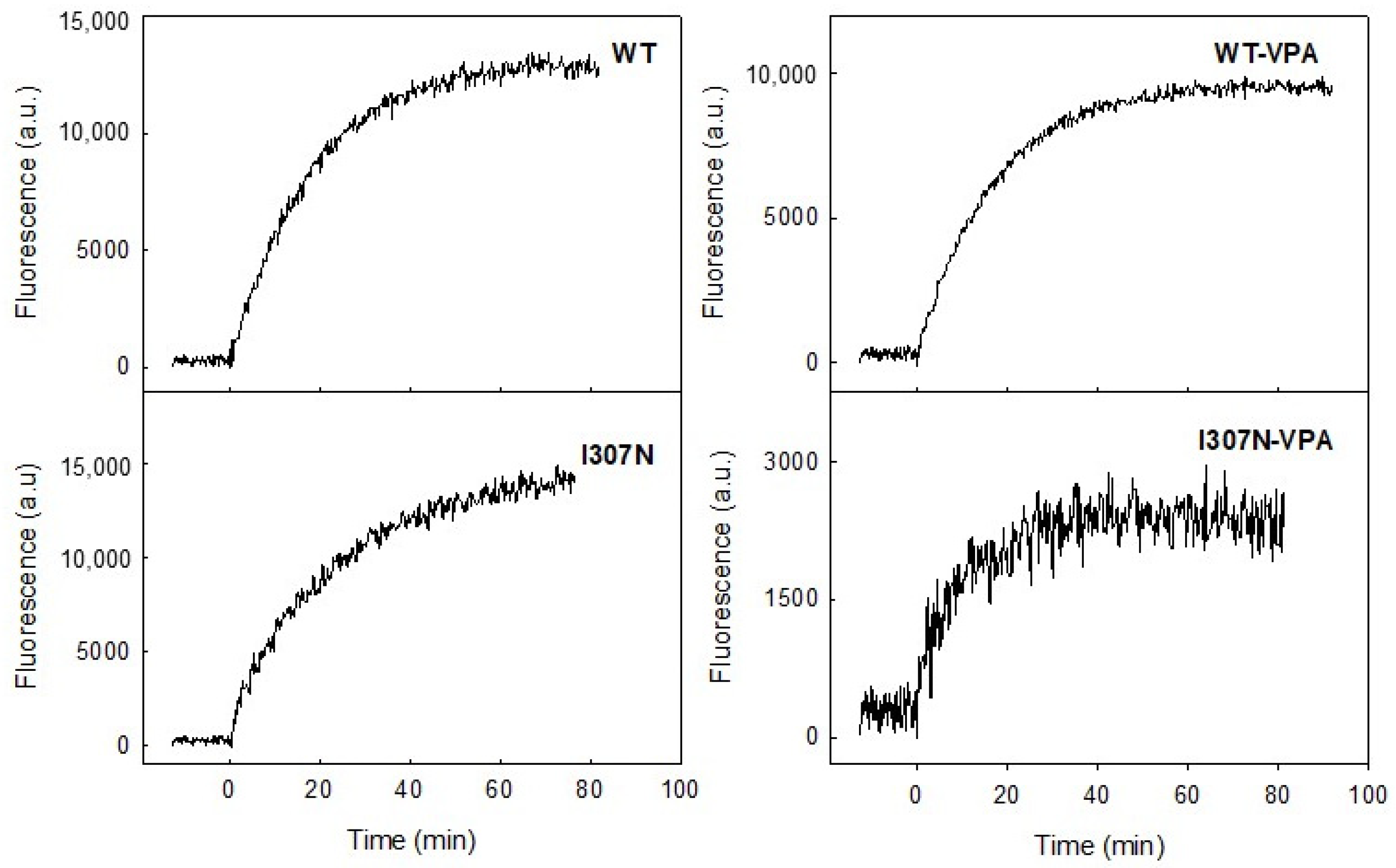
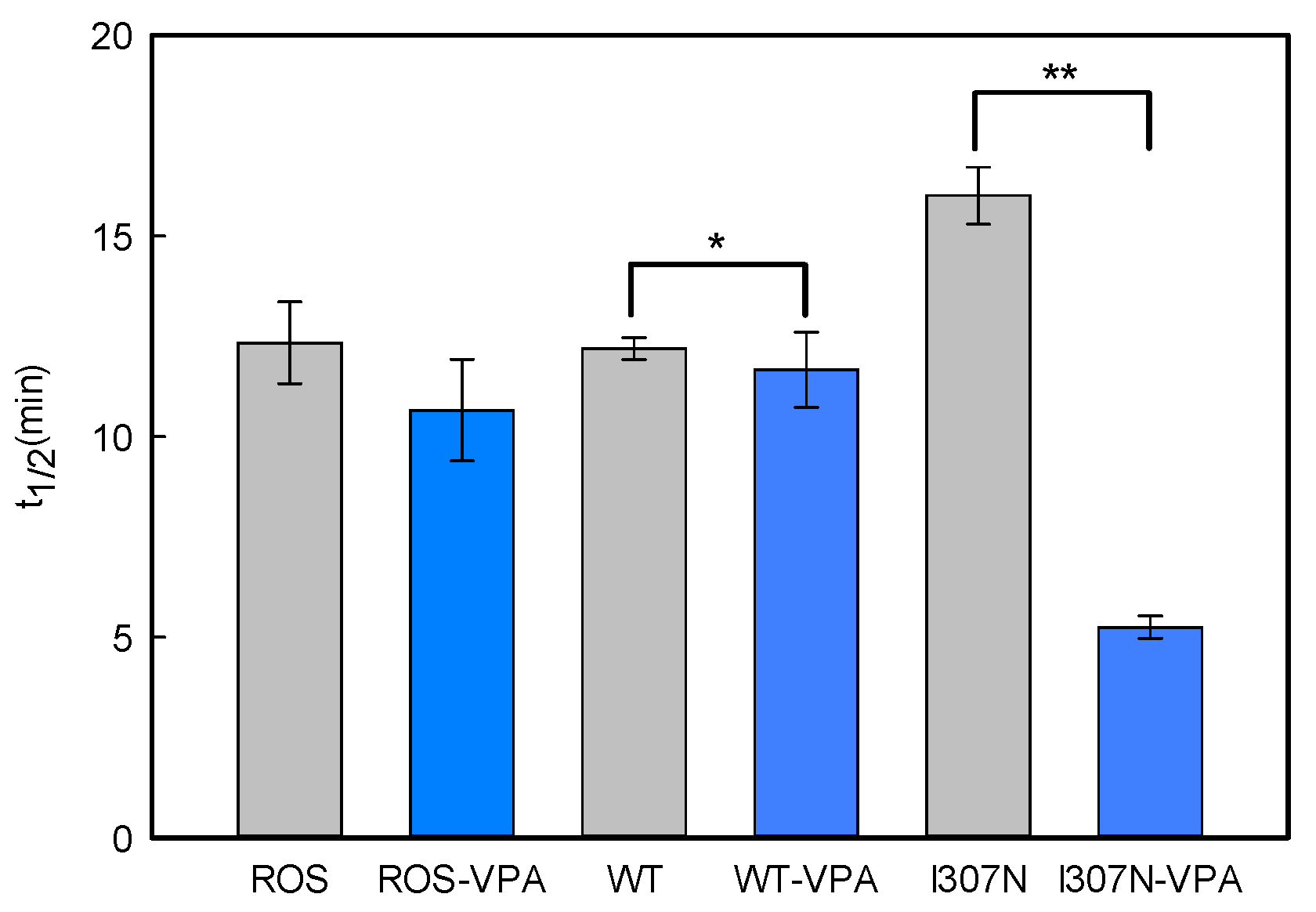
2.4. Meta II Decay
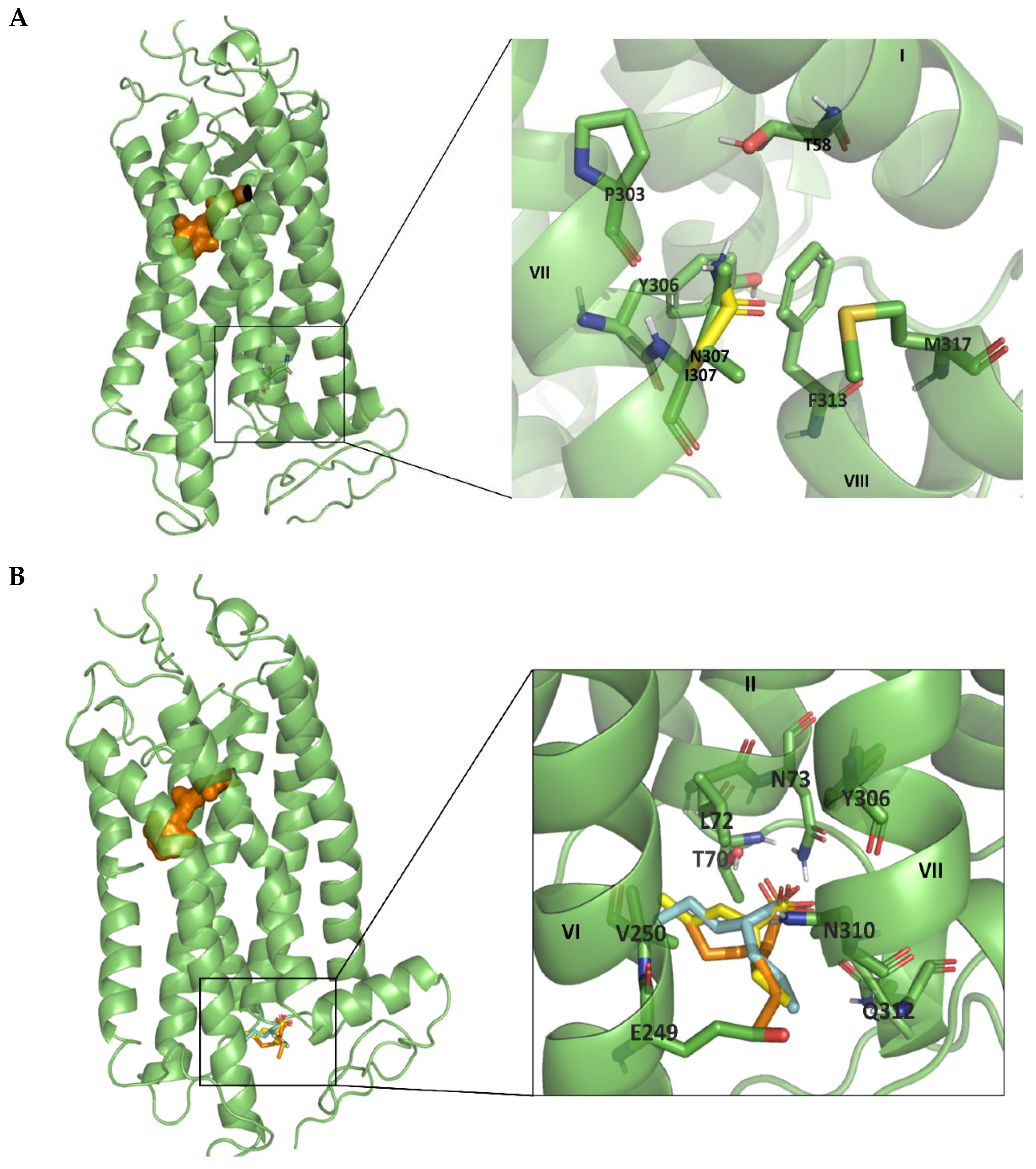
2.5. Molecular Modelling Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Methods
4.2.1. ROS Rho Purification
4.2.2. Construction of Opsin Mutant
- forward primer: CCCGGTCATCTACAACATGATGAACAAGCAGTTCC;
- reverse primer: GGAACTGCTTGTTCATCATGTTGTAGATGACCGGG.
4.2.3. Expression and Purification Bovine Recombinant WT Rho and I307N Mutant Genes
4.2.4. UV-Visible Spectral Characterization
4.2.5. Photobleaching and Acidification of Purified Rho.
4.2.6. Thermal Stability
4.2.7. Metarhodopsin II (Meta II) Decay
4.2.8. Molecular Modelling Studies
4.2.9. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Palczewski, K.; Kumasaka, T.; Hori, T.; Behnke, C.A.; Motoshima, H.; Fox, B.A.; Le Trong, I.; Teller, D.C.; Okada, T.; Stenkamp, R.E.; et al. Crystal structure of rhodopsin: A G protein-coupled receptor. Science 2000, 289, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Petrs-Silva, H.; Linden, R. Advances in gene therapy technologies to treat retinitis pigmentosa. Clin. Ophthalmol. 2014, 8, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickell, S.; Park, P.S.H.; Baumeiter, W.; Palczewski, K. Three-dimensional architecture of murine rod outer segments determined by cryoelectron tomography. J. Cell Biol. 2007, 177, 917–925. [Google Scholar] [CrossRef]
- Fanelli, F.; De Benedetti, P.G. Update 1 of: Computational Modeling Approaches to Structure Function Analysis of G Protein-Coupled Receptors. Chem. Rev. 2011, 111, 438–535. [Google Scholar] [CrossRef]
- Lupala, C.S.; Rasaeifar, B.; Gomez-Gutierrez, P.; Perez, J.J. Using Molecular Dynamics for the refinement of atomistic models of GPCRs by homology modeling. J. Biomol. Struct. Dyn. 2018, 36, 2436–2448. [Google Scholar] [CrossRef] [PubMed]
- Palczewski, K. G protein–coupled receptor rhodopsin. Annu. Rev. Biochem. 2006, 75, 743–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobilka, B. The Structural Basis of G-Protein-Coupled Receptor. Signal. Angewante Chem. Int. Ed. 2013, 52, 6380–6388. [Google Scholar] [CrossRef] [Green Version]
- Lagerstrom, M.C.; Schioth, H.B. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 339–357. [Google Scholar] [CrossRef]
- Kenakin, T. New concepts in pharmacological efficacy at 7TM receptors. IUPHAR review 2. Br. J. Pharmacol. 2013, 168, 554–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; David, E.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Schöneberg, T.; Schulz, A.; Biebermann, H.; Hermsdorf, T.; Römpler, H.; Sangkuhl, K. Mutant G-protein-coupled receptors as a cause of human diseases. Pharm. Ther. 2004, 104, 173–206. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Iorio, E.D.; Barbaro, V.; Ponzin, D.; Sorrentino, F.S.; Parmeggiani, F. Retinitis Pigmentosa: Genes and disease mechanisms. Curr. Genom. 2011, 12, 238–249. [Google Scholar]
- Rayapudi, S.; Schwartz, S.G.; Wang, X.; Chavis, P. Cochrane Eyes and Vision Group. Vitamin A and fish oils for retinitis pigmentosa. Cochrane Database Syst. Rev. 2013, 12, 8428. [Google Scholar]
- Sieving, P.A.; Caruso, R.C.; Tao, W.; Coleman, H.R.; Thompson, D.J.; Fullmer, K.R.; Bush, R.A. Ciliary neurotrophic factor (CNTF) for human retinal degeneration: Phase I trial of CNTF delivered by encapsulated cell intraocular implants. Proc. Natl. Acad. Sci. USA 2006, 103, 3896–3901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henry, T.R. The history of valproate in clinical neuroscience. Psychopharmacol. Bull. 2003, 37, 5–16. [Google Scholar]
- Totan, Y.; Güler, E.; Yüce, A.; Dervişoğulları, M.S. The adverse effects of valproic acid on visual functions in the treatment of retinitis pigmentosa. Indian. J. Ophthalmol. 2017, 65, 984–988. [Google Scholar]
- Güveli, B.T.; Rosti, R.Ö.; Güzeltaş, A.; Tuna, E.B.; Ataklı, D.; Sencer, S.; Yekeler, E.; Kayserili, H.; Dirican, A.; Bebek, N.; et al. Teratogenicity of antiepileptic drugs. Clin. Psychopharmacol. Neurosci. 2017, 15, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Kimura, A.; Namekata, K.; Guo, X.; Harada, C.; Harada, T. Neuroprotection, growth factors and BDNF-TrkB signalling in retinal degeneration. Int. J. Mol. Sci. 2016, 17, 1584. [Google Scholar] [CrossRef] [Green Version]
- Shanmugam, P.M.; Minija, C.K.; Ramanjulu, R.; Tekwani, P.; Saxena, M. Effect of short-term oral valproic acid on vision and visual field in retinitis pigmentosa. Ophtalmol Ther. 2016, 1, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yasuda, S.; Liang, M.H.; Marinova, Z.; Yahyavi, A.; Chuang, D.M. The mood stabilizers lithium and valproate selectively activate the promoter IV of brain-derived neurotrophic factor in neurons. Mol. Psychiatry 2009, 14, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Laia, C.L.; Lub, C.C.; Linc, H.C.; Sungd, Y.F.; Wud, Y.P.; Honge, J.S.; Pengb, G.S. Valproate is protective against 6-OHDA-induced dopaminergic neurodegeneration in rodent midbrain: A potential role of BDNF up-regulation. J. Formos. Med. Assoc. 2019, 118, 420–428. [Google Scholar] [CrossRef]
- Nanau, R.M.; Neuman, M.G. Adverse drug reactions induced by valproic acid. Clin. Biochem. 2013, 46, 1323–1338. [Google Scholar] [CrossRef] [PubMed]
- Clemson, C.M.; Tzekov, R.; Krebs, M.; Checchi, J.M.; Bigelow, C.; Kaushal, S. Therapeutic potential of valproic acid for retinitis pigmentosa. Br. J. Ophthalmol. 2011, 95, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.T.; Wang, Z.; Hunsberger, J.G.; Chuang, D.M. Therapeutic potential of mood stabilizers lithium and valproic acid. beyond bipolar disorder. Pharmacol. Rev. 2013, 65, 105–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frake, R.A.; Ricketts, T.; Menzies, F.M.; Rubinsztein, D.C. Autophagy and neurodegeneration. J. Clin. Invest. 2015, 125, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Midha, N.; Gogia, V.; Gupta, S.; Sehra, S.; Chohan, A. Efficacy of oral valproic acid in patients with retinitis pigmentosa. J. Ocul. Pharmacol. Ther. 2014, 30, 580–586. [Google Scholar] [CrossRef]
- Birch, D.G.; Bernstein, P.S.; Iannacone, A.; Pennesi, M.E.; Lam, B.L.; Heckenlively, J.; Csaky, K.; Hartnett, M.E.; Winthrop, K.L.; Jayasundera, T.; et al. Effect of oral valproic acid vs. Placebo for vision loss in patients with autosomal dominant retinitis pigmentosa. A randomized phase 2 multicenter placebo-controlled clinical trial. JAMA Ophthalmol. 2018, 136, 849–857. [Google Scholar] [CrossRef]
- Dong, X.; Ramon, E.; Herrera-Hernández, M.G.; Garriga, P. Phospholipid bicelles improve the conformational stability of rhodopsin mutants associated with retinitis pigmentosa. Biochemistry 2015, 54, 4795–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramon, E.; Cordomí, A.; Aguilà, M.; Srinivasan, S.; Dong, X.; Moore, A.T.; Andrew, R.; Webster, A.R.; Cheetham, M.E.; Garriga, P. Differential light-induced responses in sectorial inherited retinal degeneration. J. Biol. Chem. 2014, 289, 35918–35928. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.Y.; Liu, J.; Mehrotra, D.; Liu, Y.; Guo, Y.; Baldera-Aguayo, P.A.; Mooney, M.L.; Nour, A.M.; Yan, E.C.Y. Thermal stability of rhodopsin and progression of retinitis pigmentosa: Comparison of S186W and D190N rhodopsin mutants. J. Biol. Chem. 2013, 288, 698–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budzynski, E.; Gross, A.K.; McAlear, S.D.; Peachey, N.S.; Shukla, M.; He, F.; Edwards, M.; Won, J.; Hicks, W.L.; Wensel, T.G.; et al. Mutations of the opsin gene (Y102H and I307N) lead to light-induced degeneration of photoreceptors and constitutive activation of phototransduction in mice. J. Biol. Chem. 2010, 285, 14521–14533. [Google Scholar] [CrossRef] [Green Version]
- Vasudevan, S.; Park, P.S.-H. Differential aggregation properties of mutant human and bovine rhodopsin. Biochemistry 2021, 60, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Aguilà, M.; Toledo, D.; Morillo, M.; Dominguez, M.; Vaz, B.; Alvarez, R.; de Lera, A.R.; Garriga, P. Structural coupling of 11-cis-7-methyl-retinal and amino acids at the ligand binding pocket of rhodopsin. Photochem. Photobiol. 2009, 85, 485–493. [Google Scholar] [CrossRef]
- Gargini, C.; Novelli, E.; Piano, I.; Biagioni, M.; Strettoi, E. Pattern of retinal morphological and functional decay in a light-inducible, rhodopsin mutant mouse. Sci. Rep. 2017, 7, 5730. [Google Scholar] [CrossRef]
- Massengill, M.T.; Young, B.; Patel, D.; Jafri, F.; Sabogal, E.; Ash, N.; Li, H.; Ildefonso, C.J.; Lewin, A.S. Clinically relevant outcome measures for the I307N rhodopsin mouse: A model of inducible autosomal dominant retinitis pigmentosa. Investig. Ophthalmol. Vis. Sci. 2018, 59, 5417–5430. [Google Scholar] [CrossRef] [Green Version]
- Toledo, D.; Ramon, E.; Aguilà, M.; Cordomí, A.; Pérez, J.J.; Mendes, H.F.; Cheetham, M.E.; Garriga, P. Molecular mechanisms of disease for mutations at Gly-90 in rhodopsin. J. Biol. Chem. 2011, 286, 39993–40001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Hernández, M.G. Structural and Functional Effects of Natural Phenolic Compounds on Rhodopsin Mutants Associated with Retinitis Pigmentosa. Ph.D. Thesis, Universitat Politècnica de Catalunya Barcelonatech, Barcelona, Spain, 2017. Available online: https://upcommons.upc.edu/handle/2117/113677 (accessed on 12 April 2021).
- Liu, J.; Liu, M.Y.; Nguyen, J.B.; Bhagat, A.; Mooney, V.; Yan, E.C.Y. Thermal properties of rhodopsin: Insight into the molecular mechanism of dim–light vision. J. Biol. Chem. 2011, 286, 27622–27629. [Google Scholar] [CrossRef] [Green Version]
- Choe, H.-W.; Kim, Y.J.; Park, J.H.; Morizumi, T.; Pai, E.F.; Krauss, N.; Hofmann, K.P.; Scheerer, P.; Ernst, O.P. Crystal structure of metarhodopsin II. Nature 2011, 471, 651–655. [Google Scholar] [CrossRef]
- Herrera-Hernández, M.G.; Ramon, E.; Lupala, C.S.; TenaCampos, M.; Pérez, J.J.; Garriga, P. Flavonoid allosteric modulation of mutated visual rhodopsin associated with retinitis pigmentosa. Sci. Rep. 2017, 7, 11167. [Google Scholar] [CrossRef] [Green Version]
- Vent-Schmidt, R.Y.J.; Wen, R.H.; Zong, Z.; Chiu, C.N.; Tam, B.M.; May, C.G.; Moritz, O.L. Opposing effects of valproic acid treatment mediated by histone deacetylase inhibitor activity in four transgenic X. laevis models of retinitis pigmentosa. J. Neurosci. 2017, 37, 1039–1054. [Google Scholar] [CrossRef] [PubMed]
- Hayani, M.; Wandarian, K. Effect of valproic acid on retina. J. Ophthalmol. Res. 2019, 2, 23–25. [Google Scholar]
- Reyes-Alcaraz, A.; Martinez-Archundia, M.; Ramon, E.; Garriga, P. Salt effects on the conformational stability of the visual G-protein-coupled receptor rhodopsin. Biophys. J. 2011, 101, 2798–2806. [Google Scholar] [CrossRef] [Green Version]
- Ramon, E.; Marron, J.; del Valle, L.; Bosch, A.; Andrés, A.; Manyosa, J.; Garriga, P. Effect of dodecyl maltoside detergent on rhodopsin stability and function. Vision Res. 2003, 43, 3055–3061. [Google Scholar] [CrossRef]
- Sánchez-Martín, M.J.; Ramon, E.; Torrent-Burgués, J.; Garriga, P. Improved conformational stability of the visual G protein-coupled receptor rhodopsin by specific interaction with docosahexaenoic acid phospholipid. ChemBioChem 2013, 14, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Oprian, D.D.; Molday, R.S.; Kaufman, R.J.; Khorana, H.G. Expression of a synthetic bovine rhodopsin gene in monkey kidney cells. Proc. Natl. Acad. Sci. USA 1987, 84, 8874–8878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrodinger, L.L.C. The PyMOL Molecular Graphics System, Version 2.4. 2010. Available online: https://pymol.org/2/ (accessed on 12 April 2021).
- Case, D.A.; Ben-Shalom, I.Y.; Brozell, S.R.; Cerutti, D.S.; Cheatham, T.E.; Cruzeiro, V.W.D., III; Darden, T.A.; Duke, R.E.; Ghoreishi, D.; Gilson, M.K.; et al. AMBER18; University of California: San Francisco, CA, USA, 2018. [Google Scholar]
- Arfken, G.B.; Weber, H. Method of steepest descents. Mathematical Methods for Physicists. Press EA. 2005, 6, 489–497. [Google Scholar]
- Morris, G.M.; Ruth, H.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Software news and updates AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Alhossary, A.; Handoko, S.D.; Mu, Y.; Kwoh, C.K. Fast, accurate, and reliable molecular docking with QuickVina 2. Bioinformatics 2015, 31, 2214–2216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ROS | ROS-VPA | WT | WT–VPA | I307N | I307N-VPA | |
|---|---|---|---|---|---|---|
| λmax (nm) | 500 | 501 | 500 | 497 | 500 | 497 |
| A280/Aλmax | 1.90 ± 0.10 | 2.83 ± 0.20 | 2.22 ± 0.20 | 2.89 ± 0.20 | 2.55 ± 0.30 | 3.02 ± 0.30 |
| ε × 103 (M−1·cm−1) | 42.7 ± 1.1 | 37.6 ± 1.8 | 43.2 ± 0.6 | 37.5 ± 2.2 | 43.8 ± 0.3 | 37.8 ± 2.5 |
| ROS | ROS-VPA | WT | WT-VPA | I307N | I307N-VPA | |
|---|---|---|---|---|---|---|
| T1 (min) | 4.2 ± 0.3 | 6.3 ± 1.5 | 4.4 ± 0.6 | 6.8 ± 0.6 | 0.9 ± 0.2 | 0.7 ± 0.1 |
| T2 (min) | 35.5 ± 1.8 | 41.1 ± 2.4 | 38.1 ± 1.7 | 43.4 ± 1.8 | 25.7 ± 1.3 | 23.5 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razzaghi, N.; Fernandez-Gonzalez, P.; Mas-Sanchez, A.; Vila-Julià, G.; Perez, J.J.; Garriga, P. Effect of Sodium Valproate on the Conformational Stability of the Visual G Protein-Coupled Receptor Rhodopsin. Molecules 2021, 26, 3032. https://doi.org/10.3390/molecules26103032
Razzaghi N, Fernandez-Gonzalez P, Mas-Sanchez A, Vila-Julià G, Perez JJ, Garriga P. Effect of Sodium Valproate on the Conformational Stability of the Visual G Protein-Coupled Receptor Rhodopsin. Molecules. 2021; 26(10):3032. https://doi.org/10.3390/molecules26103032
Chicago/Turabian StyleRazzaghi, Neda, Pol Fernandez-Gonzalez, Aina Mas-Sanchez, Guillem Vila-Julià, Juan Jesus Perez, and Pere Garriga. 2021. "Effect of Sodium Valproate on the Conformational Stability of the Visual G Protein-Coupled Receptor Rhodopsin" Molecules 26, no. 10: 3032. https://doi.org/10.3390/molecules26103032