
In Vitro Antiviral Activity of α-Mangostin against Dengue Virus Serotype-2 (DENV-2)

 ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of Compound with Maximum Antiviral Activity
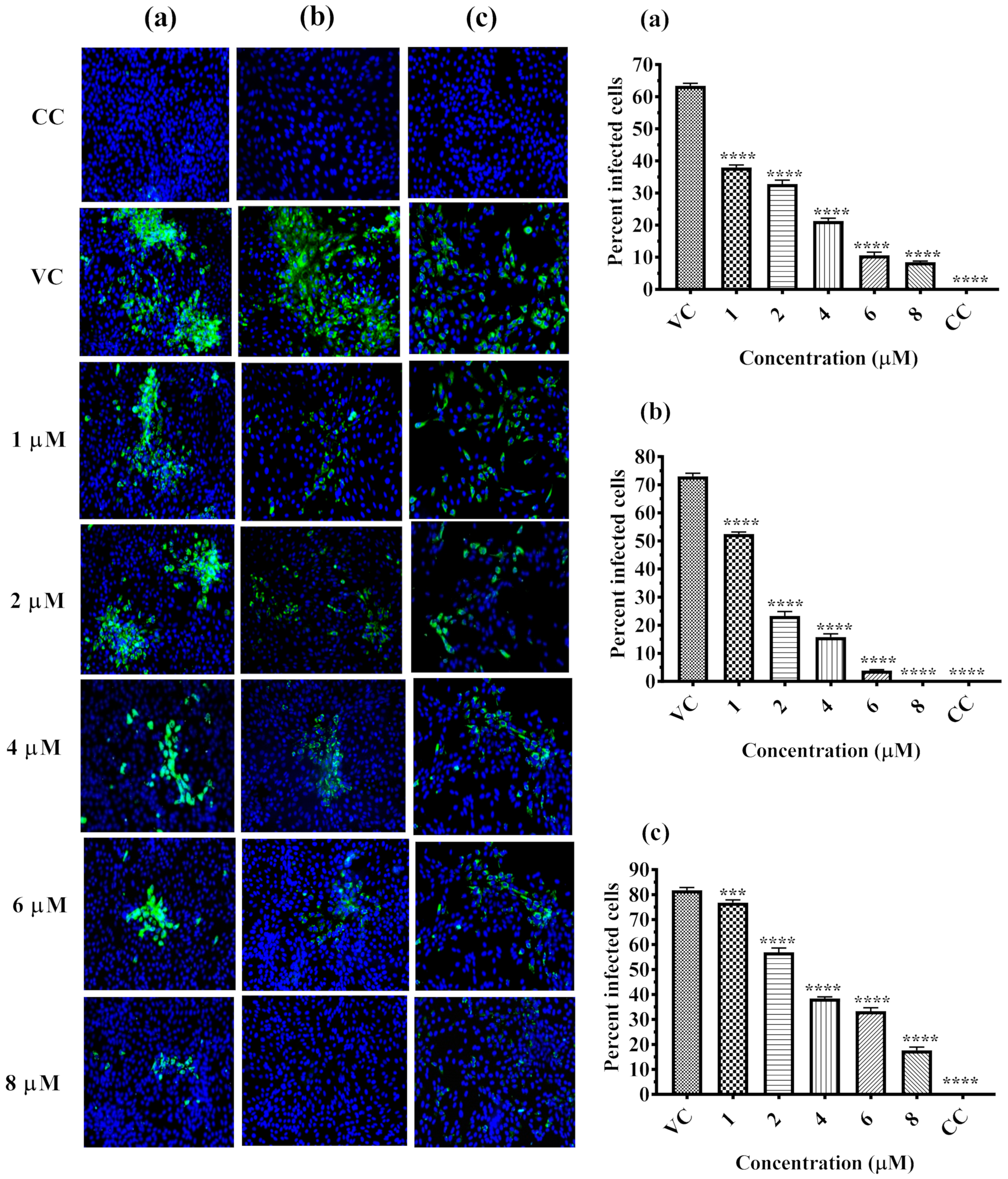
2.2. Inhibitory Effect of α-Mangostin on DENV-2 Replication
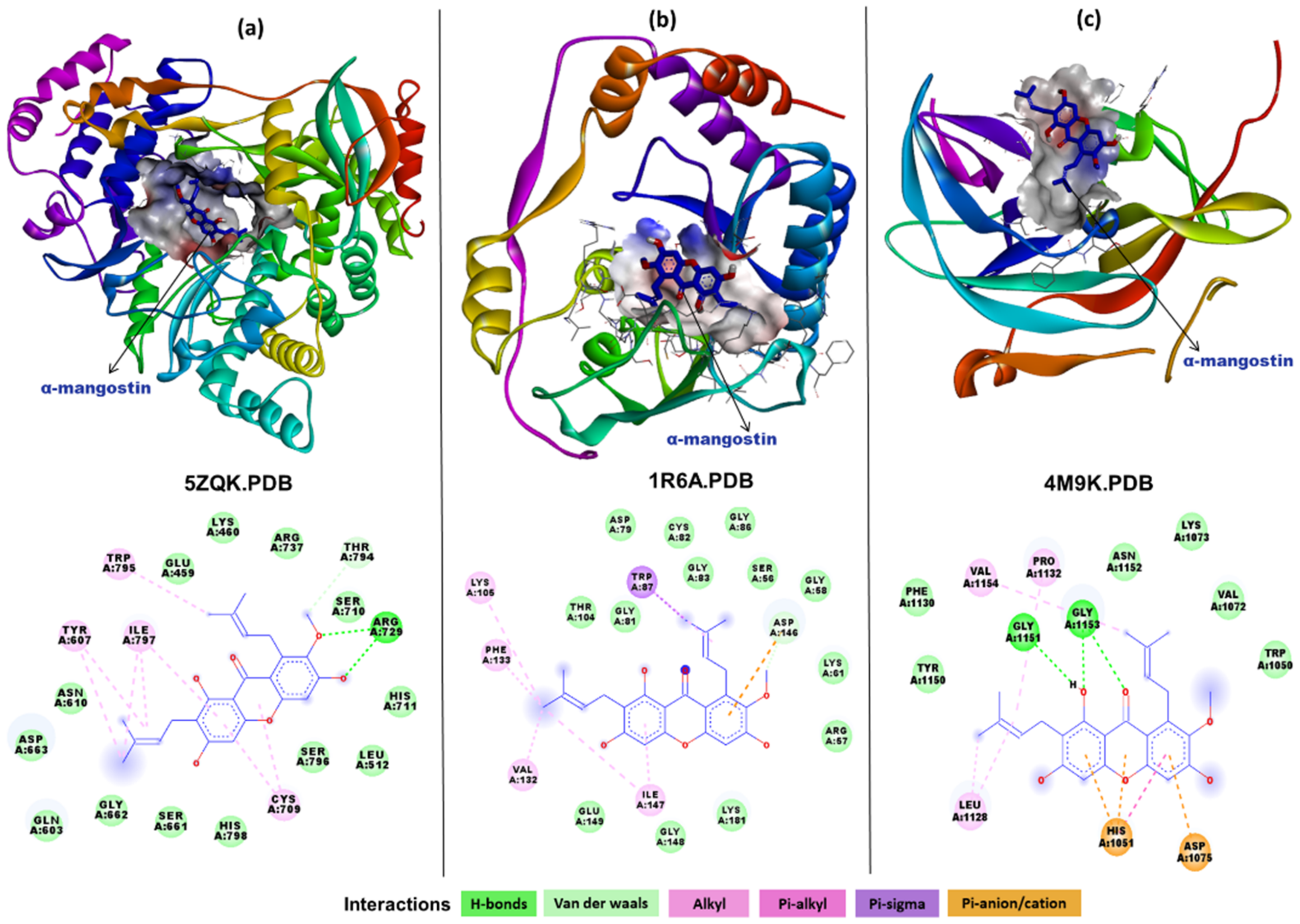
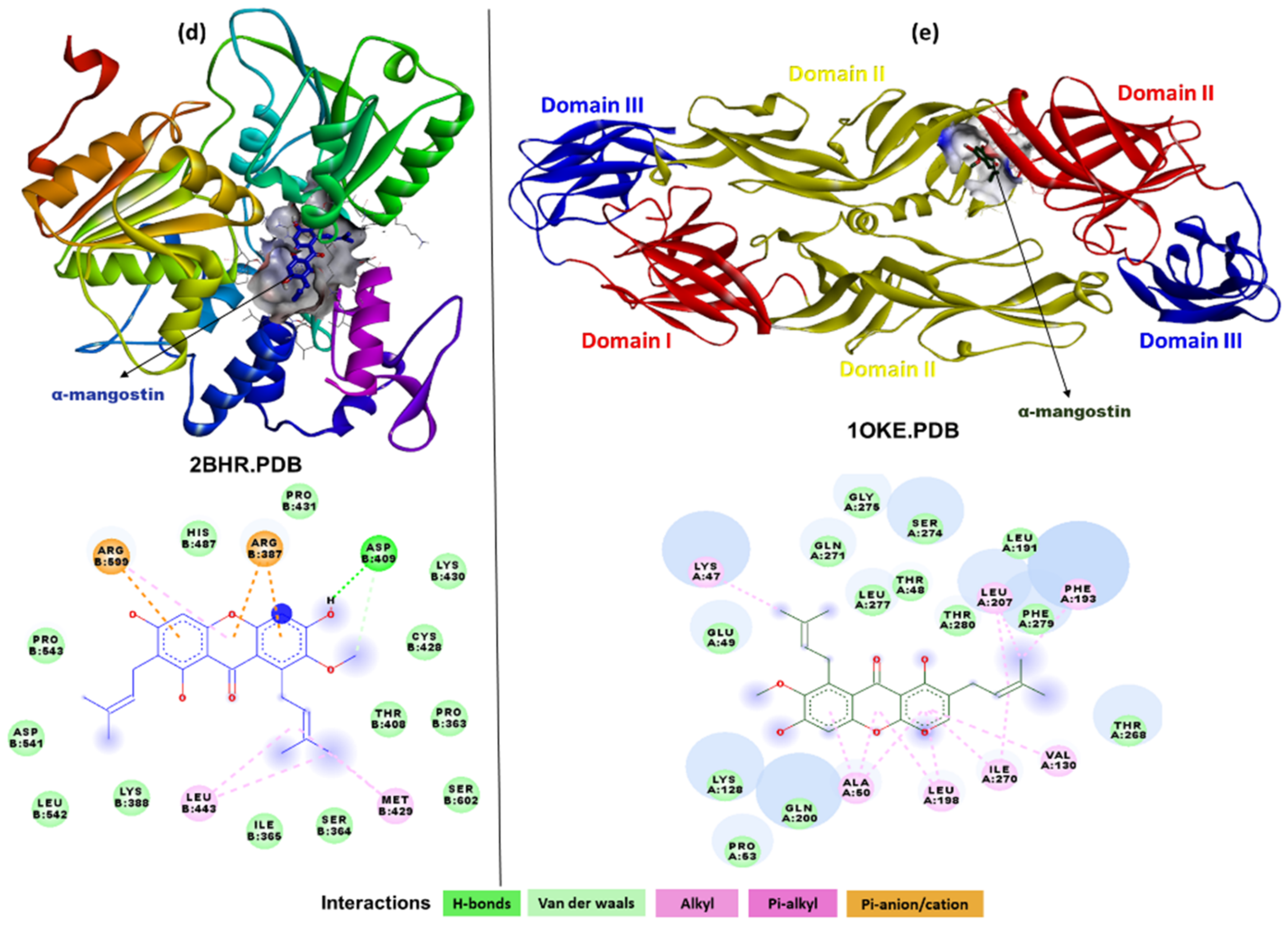
2.3. In Silico Interaction Studies of α-Mangostin with DENV Protein Targets
3. Discussion
4. Materials and Methods
4.1. Cells and Virus Maintenance
4.2. Compounds Stock Preparation
4.3. Cell Cytotoxicity Assay of Compounds
4.4. Antiviral Assays
4.4.1. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) for Estimating Viral RNA Copy Number
4.4.2. Focus Forming Unit (FFU) Assay
4.4.3. Immunofluorescence Assay
4.5. Compound, Molecular Modelling and Docking Studies with Viral Proteins Targets
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dengue and Severe Dengue; World Health Organization: Geneva, Switzerland, 2020.
- Murugesan, A.; Manoharan, M. Dengue Virus. In Emerging and Reemerging Viral Pathogens; Academic Press: Lodon, UK, 2020; pp. 281–359. [Google Scholar]
- Smartt, C.T.; Shin, D.; Alto, B.W. Dengue serotype-specific immune response in Aedes aegypti and Aedes albopictus. Mem. Inst. Oswaldo Cruz 2017, 112, 829–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higa, L.M.; Curi, B.M.; Aguiar, R.S.; Cardoso, C.C.; De Lorenzi, A.G.; Sena, S.L.; Zingali, R.B.; Da Poian, A.T. Modulation of α-enolase post-translational modifications by dengue virus: Increased secretion of the basic isoforms in infected hepatic cells. PLoS ONE 2014, 9, e88314. [Google Scholar]
- Bollati, M.; Alvarez, K.; Assenberg, R.; Baronti, C.; Canard, B.; Cook, S.; Coutard, B.; Decroly, E.; de Lamballerie, X.; Gould, E.A. Structure and functionality in flavivirus NS-proteins: Perspectives for drug design. Antivir. Res. 2010, 87, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Low, J.G.; Ooi, E.E.; Vasudevan, S.G. Current status of dengue therapeutics research and development. J. Infect. Dis. 2017, 215, S96–S102. [Google Scholar] [CrossRef] [Green Version]
- Poddar, A.; Shukla, R.; Beesetti, H.; Arora, U.; Rajpoot, R.K.; Shanmugam, R.K.; Palla, S.; Nayyar, K.; Singh, D.; Singamaneni, V. Cocculus hirsutus-derived phytopharmaceutical drug has potent anti-dengue activity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omonike, O.O.; Adekunle, J.; Edith, A.O.; Festus, A.D. Anti-poliovirus activity of medicinal plants selected from the Nigerian ethno-medicine. Afr. J. Biotechnol. 2013, 12, 3878–3883. [Google Scholar]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Gorlach, S.; Fichna, J.; Lewandowska, U. Polyphenols as mitochondria-targeted anticancer drugs. Cancer Lett. 2015, 366, 141–149. [Google Scholar] [CrossRef]
- Iranshahi, M.; Rezaee, R.; Parhiz, H.; Roohbakhsh, A.; Soltani, F. Protective effects of flavonoids against microbes and toxins: The cases of hesperidin and hesperetin. Life Sci. 2015, 137, 125–132. [Google Scholar] [CrossRef]
- Naithani, R.; Huma, L.C.; Holland, L.E.; Shukla, D.; McCormick, D.L.; Mehta, R.G.; Moriarty, R.M. Antiviral activity of phytochemicals: A comprehensive review. Mini Rev. Med. Chem. 2008, 8, 1106–1133. [Google Scholar] [CrossRef]
- Zakaryan, H.; Arabyan, E.; Oo, A.; Zandi, K. Flavonoids: Promising natural compounds against viral infections. Arch. Virol. 2017, 162, 2539–2551. [Google Scholar] [CrossRef]
- Simões, L.; Maciel, G.; Brandão, G.; Kroon, E.; Castilho, R.; Oliveira, A. Antiviral activity of Distictella elongata (Vahl) Urb.(Bignoniaceae), a potentially useful source of anti-dengue drugs from the state of Minas Gerais, Brazil. Lett. Appl. Microbiol. 2011, 53, 602–607. [Google Scholar] [CrossRef]
- Zandi, K.; Teoh, B.-T.; Sam, S.-S.; Wong, P.-F.; Mustafa, M.R.; AbuBakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J. 2011, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiat, T.S.; Pippen, R.; Yusof, R.; Ibrahim, H.; Khalid, N.; Abd Rahman, N. Inhibitory activity of cyclohexenyl chalcone derivatives and flavonoids of fingerroot, Boesenbergia rotunda (L.), towards dengue-2 virus NS3 protease. Bioorg. Med. Chem. Lett. 2006, 16, 3337–3340. [Google Scholar] [CrossRef] [PubMed]
- Leardkamolkarn, V.; Sirigulpanit, W.; Phurimsak, C.; Kumkate, S.; Himakoun, L.; Sripanidkulchai, B. The inhibitory actions of Houttuynia cordata aqueous extract on dengue virus and dengue-infected cells. J. Food Biochem. 2012, 36, 86–92. [Google Scholar] [CrossRef]
- Fried, J.R.; Gibbons, R.V.; Kalayanarooj, S.; Thomas, S.J.; Srikiatkhachorn, A.; Yoon, I.-K.; Jarman, R.G.; Green, S.; Rothman, A.L.; Cummings, D.A. Serotype-specific differences in the risk of dengue hemorrhagic fever: An analysis of data collected in Bangkok, Thailand from 1994 to 2006. PLoS Negl. Trop Dis. 2010, 4, e617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidari, K.I.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef]
- Yin Low, J.; Chen, K.; Wu, K.; Mah-LeeNm, H. Antiviral activity of emetine dihydrochloride against dengue virus infection. J. Antivir. Antiretrovir. 2009, 1, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Allard, P.-M.; Leyssen, P.; Martin, M.-T.; Bourjot, M.; Dumontet, V.; Eydoux, C.; Guillemot, J.-C.; Canard, B.; Poullain, C.; Guéritte, F. Antiviral chlorinated daphnane diterpenoid orthoesters from the bark and wood of Trigonostemon cherrieri. Phytochemistry 2012, 84, 160–168. [Google Scholar] [CrossRef]
- Laurent, D.; Baumann, F.; Benoit, A.G.; Mortelecqe, A.; Nitatpattana, N.; Desvignes, I.; Debitus, C.; Laille, M.; Gonzalez, J.; Chungue, E. Structure-activity relationships of dengue antiviral polycyclic quinones. Southeast Asian J. Trop. Med. Public Health 2005, 36, 901. [Google Scholar] [PubMed]
- Bourjot, M.; Leyssen, P.; Eydoux, C.; Guillemot, J.-C.; Canard, B.; Rasoanaivo, P.; Guéritte, F.; Litaudon, M. Flacourtosides A–F, phenolic glycosides isolated from Flacourtia ramontchi. J. Nat. Prod. 2012, 75, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Kohn, L.; Foglio, M.; Rodrigues, R.; Sousa, I.d.O.; Martini, M.; Padilla, M.; de Lima Neto, D.; Arns, C. In vitro antiviral activities of extracts of plants of the Brazilian cerrado against the avian metapneumovirus (aMPV). Braz. J. Poult. Sci. 2015, 17, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Do, H.T.T.; Cho, J. Mangosteen pericarp and its bioactive xanthones: Potential therapeutic value in Alzheimer’s disease, Parkinson’s Disease, and depression with pharmacokinetic and safety profiles. Int. J. Mol. Sci. 2020, 21, 6211. [Google Scholar] [CrossRef] [PubMed]
- Obolskiy, D.; Pischel, I.; Siriwatanametanon, N.; Heinrich, M. Garcinia mangostana L.: A phytochemical and pharmacological review. Phytother. Res. 2009, 31, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.; Kim, Y.-M.; Lee, S.; Chin, Y.-W.; Lee, C. Mangosteen xanthones suppress hepatitis C virus genome replication. Virus Genes 2014, 49, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Tarasuk, M.; Songprakhon, P.; Chimma, P.; Sratongno, P.; Na-Bangchang, K.; Yenchitsomanus, P.-T. Alpha-mangostin inhibits both dengue virus production and cytokine/chemokine expression. Virus Res. 2017, 240, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Sugiyanto, Z.; Yohan, B.; Hadisaputro, S.; Dharmana, E.; Suharti, C.; Djamiatun, K.; Rahmi, F.L.; Sasmono, R.T. Inhibitory Effect of Alpha-Mangostin to Dengue Virus Replication and Cytokines Expression in Human Peripheral Blood Mononuclear Cells. Nat. Prod. Bioprospect. 2019, 9, 345–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Modis, Y.; Ogata, S.; Clements, D.; Harrison, S.C. A ligand-binding pocket in the dengue virus envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2003, 100, 6986–6991. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Barragán, J.J.; del Angel, R.M. Identification of a putative coreceptor on Vero cells that participates in dengue 4 virus infection. J. Virol. 2001, 75, 7818–7827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potisopon, S.; Priet, S.; Collet, A.; Decroly, E.; Canard, B.; Selisko, B. The methyltransferase domain of dengue virus protein NS5 ensures efficient RNA synthesis initiation and elongation by the polymerase domain. Nucleic Acids Res. 2016, 44, 2974. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Deval, J.; Johnson, K.A.; Swinney, D.C. Characterization of the elongation complex of dengue virus RNA polymerase: Assembly, kinetics of nucleotide incorporation, and fidelity. J. Biol. Chem. 2011, 286, 2067–2077. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.P.; Noble, C.G.; Seh, C.C.; Soh, T.S.; El Sahili, A.; Chan, G.K.Y.; Lescar, J.; Arora, R.; Benson, T.; Nilar, S. Potent allosteric dengue virus NS5 polymerase inhibitors: Mechanism of action and resistance profiling. PLoS Pathog. 2016, 12, e1005737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokokawa, F.; Nilar, S.; Noble, C.G.; Lim, S.P.; Rao, R.; Tania, S.; Wang, G.; Lee, G.; Hunziker, J.R.; Karuna, R. Discovery of potent non-nucleoside inhibitors of dengue viral RNA-dependent RNA polymerase from a fragment hit using structure-based drug design. J. Med. Chem. 2016, 59, 3935–3952. [Google Scholar] [CrossRef] [PubMed]
- Noble, C.G.; Lim, S.P.; Arora, R.; Yokokawa, F.; Nilar, S.; Seh, C.C.; Wright, S.K.; Benson, T.E.; Smith, P.W.; Shi, P.-Y. A conserved pocket in the dengue virus polymerase identified through fragment-based screening. J. Biol. Chem. 2016, 291, 8541–8548. [Google Scholar] [CrossRef] [Green Version]
- Yap, T.L.; Xu, T.; Chen, Y.-L.; Malet, H.; Egloff, M.-P.; Canard, B.; Vasudevan, S.G.; Lescar, J. Crystal structure of the dengue virus RNA-dependent RNA polymerase catalytic domain at 1.85-angstrom resolution. J. Virol. 2007, 81, 4753–4765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Bock, S.; Snitko, M.; Berger, T.; Weidner, T.; Holloway, S.; Kanitz, M.; Diederich, W.E.; Steuber, H.; Walter, C. Novel dengue virus NS2B/NS3 protease inhibitors. Antimicrob. Agents Chemother. 2015, 59, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Sun, Z.; Pei, J.; Luo, Q.; Zeng, X.; Li, Q.; Yang, Z.; Quan, J. Identification of α-Mangostin as an Agonist of Human STING. ChemMedChem 2018, 13, 2057–2064. [Google Scholar] [CrossRef]
- Grandvaux, N.; Servant, M.J.; Sen, G.C.; Balachandran, S.; Barber, G.N.; Lin, R.; Hiscott, J. Transcriptional profiling of interferon regulatory factor 3 target genes: Direct involvement in the regulation of interferon-stimulated genes. J. Virol. 2002, 76, 5532–5539. [Google Scholar] [CrossRef] [Green Version]
- Song, J.-H.; Ahn, J.-H.; Kim, S.-R.; Cho, S.; Hong, E.-H.; Kwon, B.-E.; Kim, D.-E.; Choi, M.; Choi, H.-J.; Cha, Y. Manassantin B shows antiviral activity against coxsackievirus B3 infection by activation of the STING/TBK-1/IRF3 signalling pathway. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Patil, P.; Agrawal, M.; Almelkar, S.; Jeengar, M.K.; More, A.; Alagarasu, K.; Kumar, N.V.; Mainkar, P.S.; Parashar, D.; Cherian, S. In vitro and in vivo studies reveal α-Mangostin, a xanthonoid from Garcinia mangostana, as a promising natural antiviral compound against chikungunya virus. Virol. J. 2021, 18, 47. [Google Scholar] [CrossRef] [PubMed]
- Parashar, D.; Paingankar, M.S.; Kumar, S.; Gokhale, M.D.; Sudeep, A.; Shinde, S.B.; Arankalle, V. Administration of E2 and NS1 siRNAs inhibit chikungunya virus replication in vitro and protects mice infected with the virus. PLoS Negl. Trop Dis. 2013, 7, e2405. [Google Scholar] [CrossRef] [PubMed]
- Gurukumar, K.; Priyadarshini, D.; Patil, J.; Bhagat, A.; Singh, A.; Shah, P.; Cecilia, D. Development of real time PCR for detection and quantitation of dengue viruses. Virol. J. 2009, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Alagarasu, K.; Patil, P.; Shil, P.; Seervi, M.; Kakade, M.; Tillu, H.; Salunke, A. In vitro effect of human cathelicidin antimicrobial peptide LL-37 on dengue virus type 2. Peptides 2017, 92, 23–30. [Google Scholar] [CrossRef]
- El Sahili, A.; Soh, T.S.; Schiltz, J.; Gharbi-Ayachi, A.; Seh, C.C.; Shi, P.-Y.; Lim, S.P.; Lescar, J. NS5 from dengue virus serotype 2 can adopt a conformation analogous to that of its Zika virus and Japanese encephalitis virus homologues. J. Virol. 2019, 94. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panda, K.; Alagarasu, K.; Patil, P.; Agrawal, M.; More, A.; Kumar, N.V.; Mainkar, P.S.; Parashar, D.; Cherian, S. In Vitro Antiviral Activity of α-Mangostin against Dengue Virus Serotype-2 (DENV-2). Molecules 2021, 26, 3016. https://doi.org/10.3390/molecules26103016
Panda K, Alagarasu K, Patil P, Agrawal M, More A, Kumar NV, Mainkar PS, Parashar D, Cherian S. In Vitro Antiviral Activity of α-Mangostin against Dengue Virus Serotype-2 (DENV-2). Molecules. 2021; 26(10):3016. https://doi.org/10.3390/molecules26103016
Chicago/Turabian StylePanda, Kingshuk, Kalichamy Alagarasu, Poonam Patil, Megha Agrawal, Ashwini More, Naveen V. Kumar, Prathama S. Mainkar, Deepti Parashar, and Sarah Cherian. 2021. "In Vitro Antiviral Activity of α-Mangostin against Dengue Virus Serotype-2 (DENV-2)" Molecules 26, no. 10: 3016. https://doi.org/10.3390/molecules26103016