
Garcinielliptone G from Garcinia subelliptica Induces Apoptosis in Acute Leukemia Cells

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
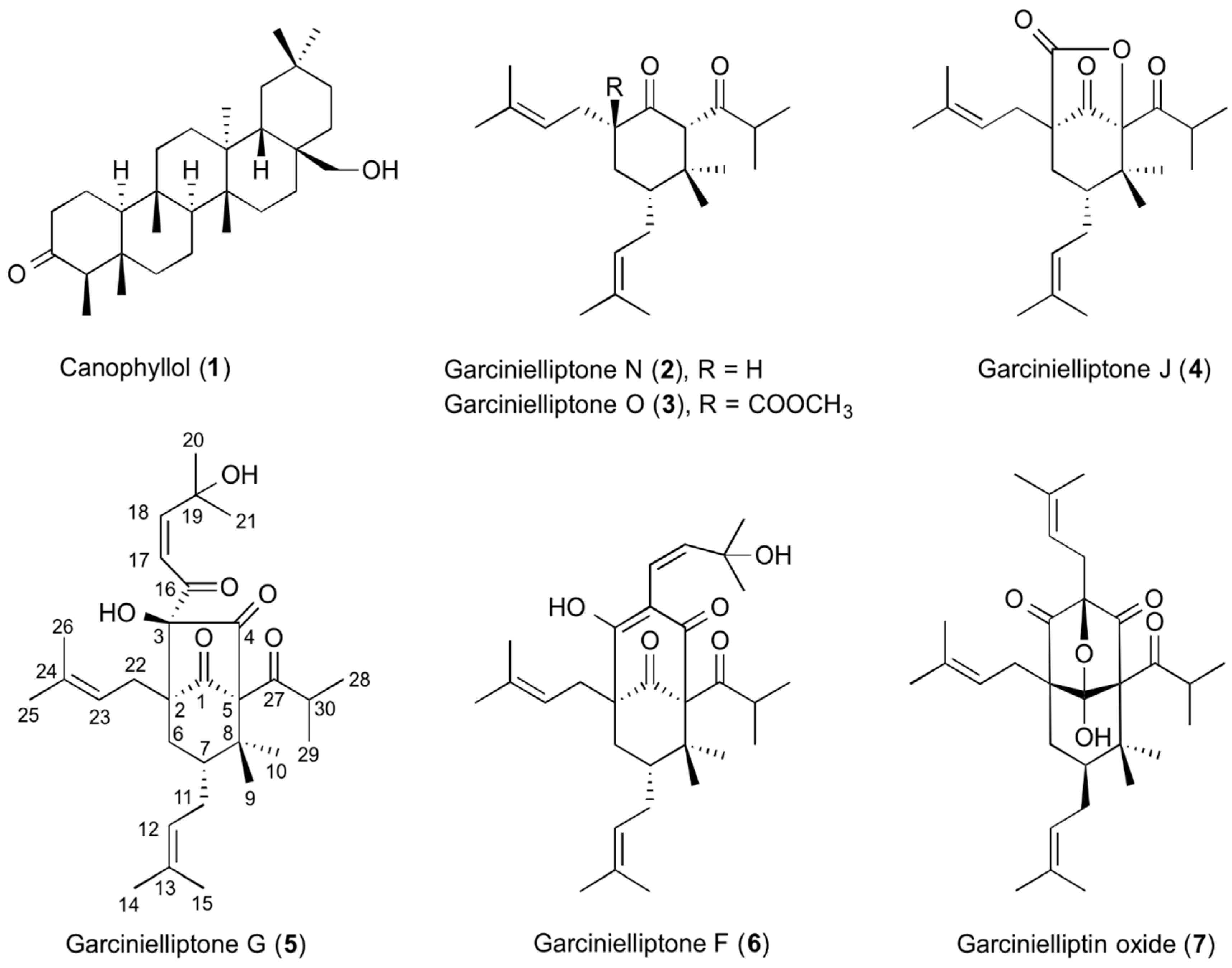
2.1. Purification and Identification of Compounds from G. subelliptica Leaves
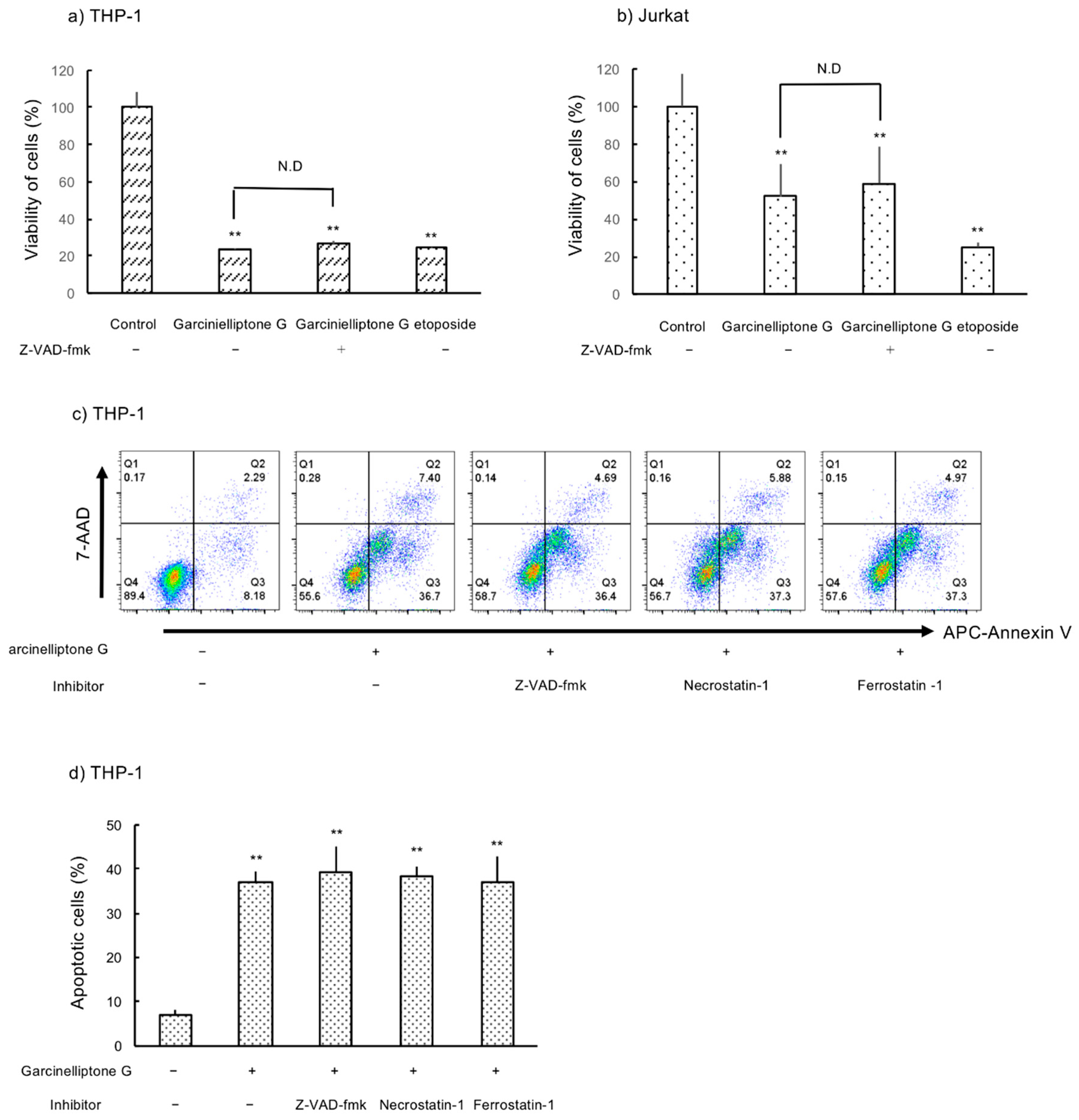
2.2. Inhibition of Cell Growth in THP-1 and Jurkat Cells
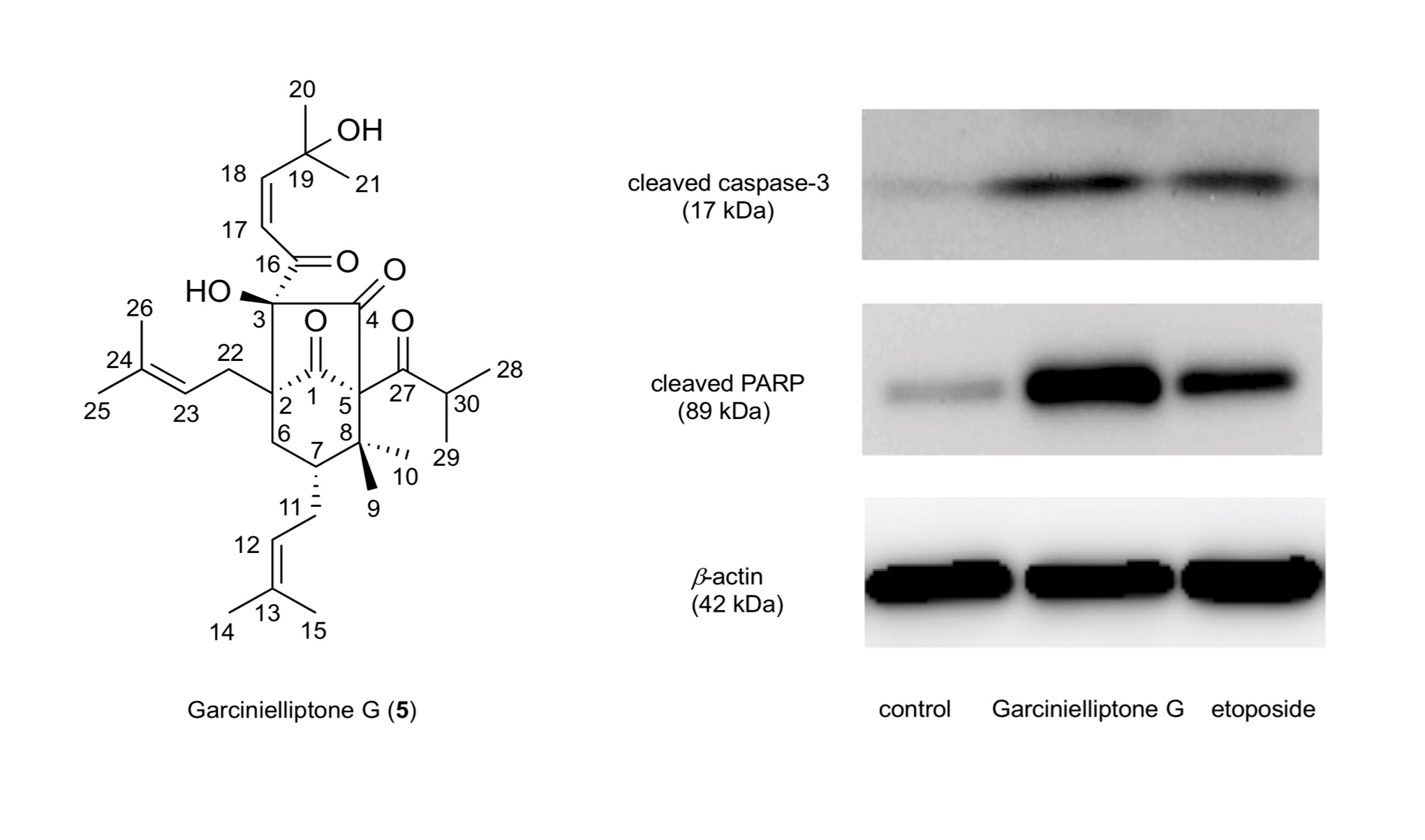
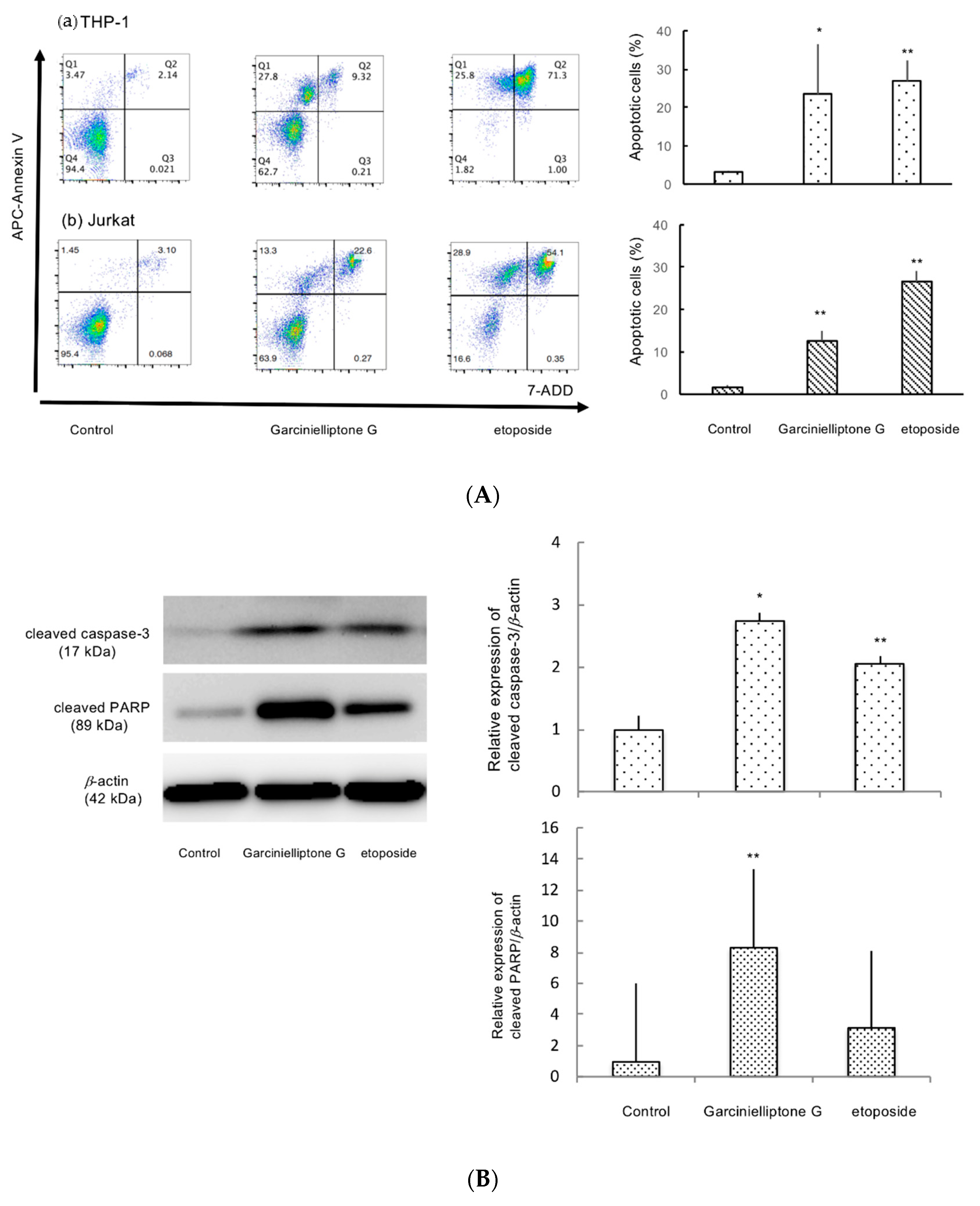
2.3. Apoptosis Induction by Garcinielliptone G (5)
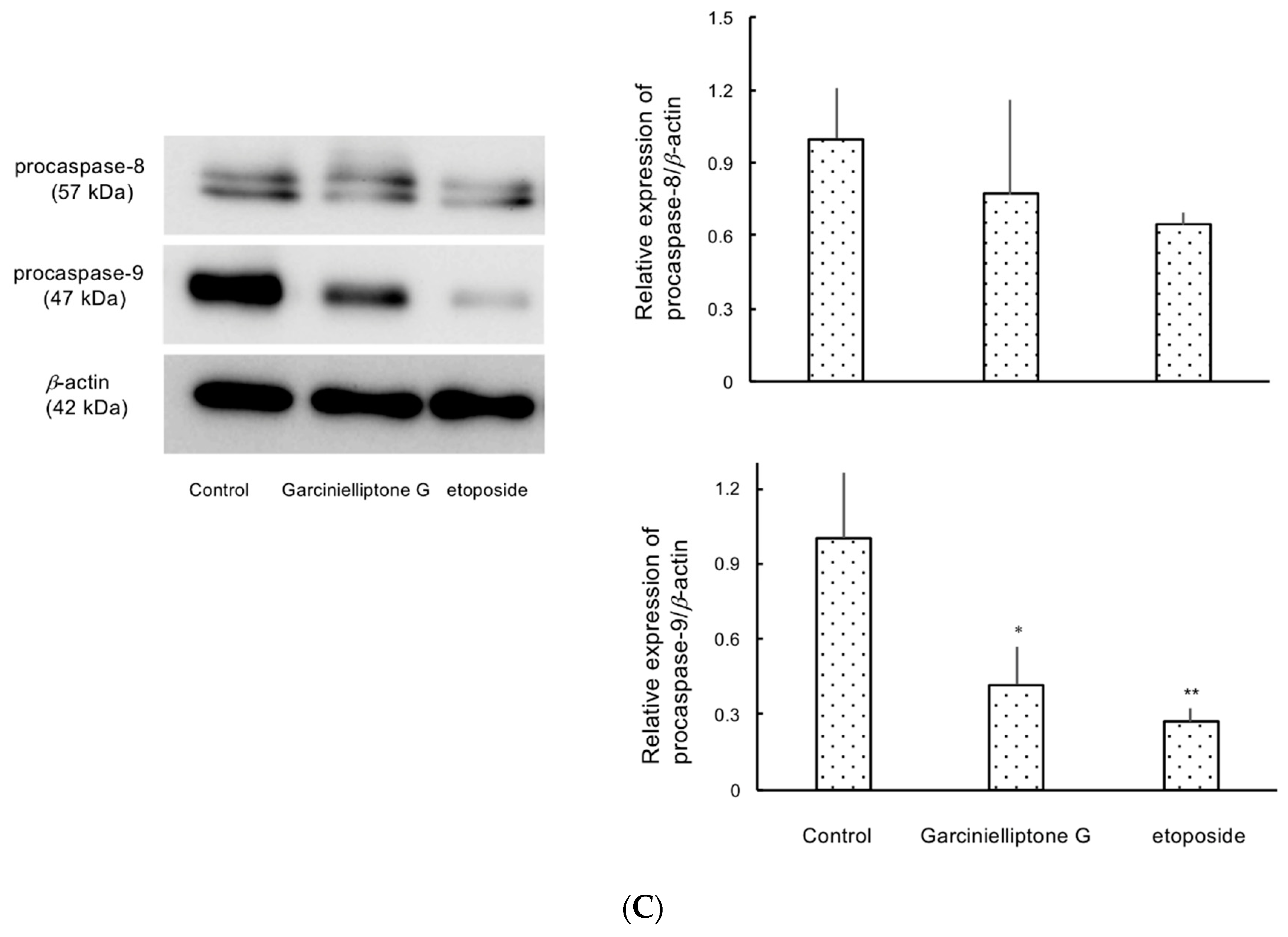
2.4. Caspase-Independent Apoptosis Induction by Garcinielliptone G (5)
3. Discussion
4. Materials and Method
4.1. Plant Materials
4.2. Reagents
4.3. Instrumentation
4.4. Extraction, Isolation and Purification of Compounds from G. subelliptica Leaves
4.5. Cell Culture
4.6. Determination of Cell Viability and Cell Death
4.7. Western Blot Analysis
4.8. Nuclear Extraction
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, J.; Nimer, S. Recent advances in the understanding and treatment of acute myeloid leukemia. F1000Research 2018, 7, 1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasov, A.G.; Walternberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.W.; Liu, P.F.; Tsai, W.L.; Hu, W.H.; Hu, Y.C.; Yang, H.C.; Lin, W.Y.; Weng, J.R.; Shu, C.W. Xanthium strumarium Fruit Extract Inhibits ATG4B and Diminishes the Proliferation and Metastatic Characteristics of Colorectal Cancer Cells. Toxin 2019, 11, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodroj, M.H.; Jardaly, A.; Abi, R.S.; Zouein, A.; Rizk, S. Andrographolide potentiates the antitumor effect of topotecan in acute myeloid leukemia cells through an intrinsic apoptotic pathway. Cancer Manag. Res. 2018, 10, 1079–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalife, R.; Hodroj, M.H.; Fakhoury, R.; Rizk, S. Thymoquinone from Nigella sativa Seeds Promotes the Antitumor Activity of Noncytotoxic Doses of Topotecan in Human Colorectal Cancer Cells in Vitro. Planta Med. 2016, 82, 312–321. [Google Scholar] [CrossRef]
- Hwang, D.W.; Kim, M.S.; Park, H.J.; Jeong, M.I.; Jung, W.J.; Kim, B.L. Natural Products and Acute Myeloid Leukemia: A Review Highlighting Mechanisms of Action. Nutrient 2019, 11, 1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.C.; Cho, S.Y. c‑Jun N‑terminal kinase‑mediated anti‑inflammatory effects of Garcinia subelliptica in macrophages. Mol. Med. Rep. 2016, 13, 2293–2300. [Google Scholar] [CrossRef] [Green Version]
- Tewtrakul, S.; Wattanapiromsakul, C.; Mahabusarakam, E. Effects of compounds from Garcinia mangostana on inflammatory mediators in RAW264.7 macrophage cells. J. Ethnopharmacol. 2009, 121, 379–382. [Google Scholar] [CrossRef]
- Panthong, A.; Norkaew, P.; Kanjanapothi, D.; Taesotikul, T.; Anatachoke, N.; Reutrakul, V. Anti-inflammatory, analgesic and antipyretic activities of the extract of gamboge from Garcinia hanburyi Hook f. J. Ethnopharmacol. 2007, 111, 335–340. [Google Scholar] [CrossRef]
- Castardo, J.C.; Prudente, A.S.; Ferreira, J.; Guimãres, C.L.; Monache, F.D.; Filho, V.C.; Otuki, M.F.; Cabrini, D.A. Anti-inflammatory effects of hydroalcoholic extract and two biflavonoids from Garcinia gardneriana leaves in mouse paw oedema. J. Ethnopharmacol. 2008, 118, 405–411. [Google Scholar] [CrossRef]
- Inoue, T.; Kainuma, M.; Baba, K.; Oshiro, N.; Kimura, N.; Chan, E.W.C. Garcinia subelliptica Merr. (Fukugi): Multipurpose coastal tree with promising medicinal properties. J. Intercult. Ethnopharmacol. 2017, 6, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Nozaki, H.; Suzuki, H.; Hirayama, T.; Kasai, R.; Wu, R.Y.; Lee, K.H. Antitumour Triterpenes of Maytenus diversifolia. Phytochemistry 1986, 25, 479–485. [Google Scholar] [CrossRef]
- Weng, J.R.; Tsao, L.T.; Wang, J.P.; Wu, R.R.; Lin, C.N. Anti-inflammatory Phloroglucinols and Terpenoids from Garcinia subelliptica. J. Nat. Prod. 2004, 67, 1796–1799. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.R.; Lin, C.N.; Tsao, L.T.; Wang, J.P. Terpenoids with a New Skeleton and Novel Triterpenoids with Antiinflammatory Effects from Garcinia subelliptica. J. Chem. Eur. 2003, 9, 5520–5527. [Google Scholar] [CrossRef]
- Lin, C.N.; Kiang, C.W.; Lu, C.M.; Wu, R.R.; Lee, K.H. Garcinielliptin oxide, a triterpenoid with a novel skeleton, isolated from Garcinia subelliptica. Chem. Commun. 1996, 11, 1315–1316. [Google Scholar] [CrossRef]
- Grossman, R.B.; Yang, X.W. Structural Revision of Garcinielliptin Oxide and Garcinielliptone E. J. Nat. Prod. 2020, 83, 2041–2044. [Google Scholar] [CrossRef]
- Kumar, S. Caspase function in programmed cell death. Cell Death. Differ. 2007, 14, 32–43. [Google Scholar] [CrossRef]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspses. Oncogene 2003, 22, 8543–8567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minami, H.; Takahashi, E.; Kodama, M.; Fukuyama, Y. Three Xanthones from Garcinia Subelliptica. Phytochemistry 1996, 41, 629–633. [Google Scholar] [CrossRef]
- Iinuma, M.; Tosa, H.; Tanaka, T.; Asai, F.; Shimano, R. Two xanthones with a 1,1-dimethylallyl group in root bark of Garcinia subelliptica. Phytochemistry 1995, 39, 945–947. [Google Scholar] [CrossRef]
- Ito, T.; Yokota, R.; Watarai, T.; Mori, K.; Oyama, M.; Nagasawa, H.; Matsuda, H.; Iinuma, M. Isolation of Six Isoprenylated Biflavonoids from the Leaves of Garcinia Subelliptica. Chem. Pharm. Bull. 2013, 61, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.C.; Weng, J.R.; Won, S.J.; Lin, C.N. Constituents of the Pericarp of Garcinia subelliptica. J. Nat. Prod. 2005, 68, 1125–1127. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Chiou, C.T.; Cheng, J.J.; Huang, H.C.; Kuo, L.M.Y.; Liao, C.C.; Bastow, K.F.; Lee, K.H.; Kuo, Y.H. Cytotoxic Polyisoprenyl Benzophenonoids from Garcinia subelliptica. J. Nat. Prod. 2010, 73, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Loo, G.V.; Schotte, P.; Gurp, M.V.; Demol, H.; Hoorelbeke, B.; Gevaert, K.; Rodriguez, I.; Ruiz-Carrillo, A.; Vandekerckhove, J.; Declercq, W.; et al. Endonuclease G: A mitochondrial protein released in apoptosis and involved in caspase-independent DNA degradation. Cell Death Differ. 2001, 8, 1136–1142. [Google Scholar] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, Y.; Shioura, M.; Hitotsuyanagi, Y.; Yotsumoto, S.; Takahashi, Y.; Aoyagi, Y.; Kinoshita, T.; Takeya, K.; Inoue, H. Garcinielliptone G from Garcinia subelliptica Induces Apoptosis in Acute Leukemia Cells. Molecules 2021, 26, 2422. https://doi.org/10.3390/molecules26092422
Yun Y, Shioura M, Hitotsuyanagi Y, Yotsumoto S, Takahashi Y, Aoyagi Y, Kinoshita T, Takeya K, Inoue H. Garcinielliptone G from Garcinia subelliptica Induces Apoptosis in Acute Leukemia Cells. Molecules. 2021; 26(9):2422. https://doi.org/10.3390/molecules26092422
Chicago/Turabian StyleYun, YoungSook, Mariko Shioura, Yukio Hitotsuyanagi, Satoshi Yotsumoto, Yuji Takahashi, Yutaka Aoyagi, Takeshi Kinoshita, Koichi Takeya, and Hideshi Inoue. 2021. "Garcinielliptone G from Garcinia subelliptica Induces Apoptosis in Acute Leukemia Cells" Molecules 26, no. 9: 2422. https://doi.org/10.3390/molecules26092422