
Changes in Particle Size, Sedimentation, and Protein Microstructure of Ultra-High-Temperature Skim Milk Considering Plasmin Concentration and Storage Temperature


Abstract
:1. Introduction
2. Results and Discussion
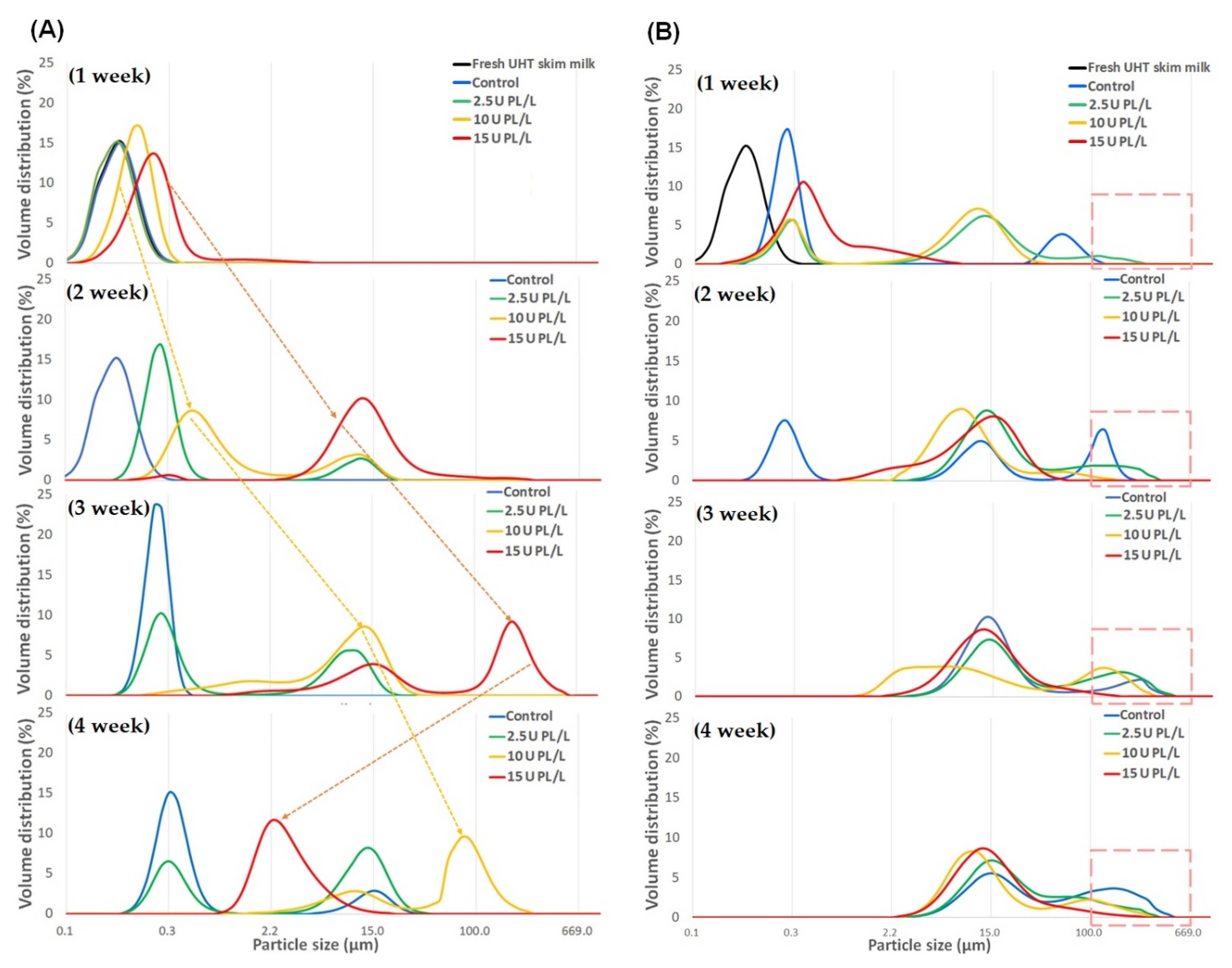
2.1. Changes in Particle Size Distribution of PL-Added UHT Skim Milk Stored at Different Temperatures
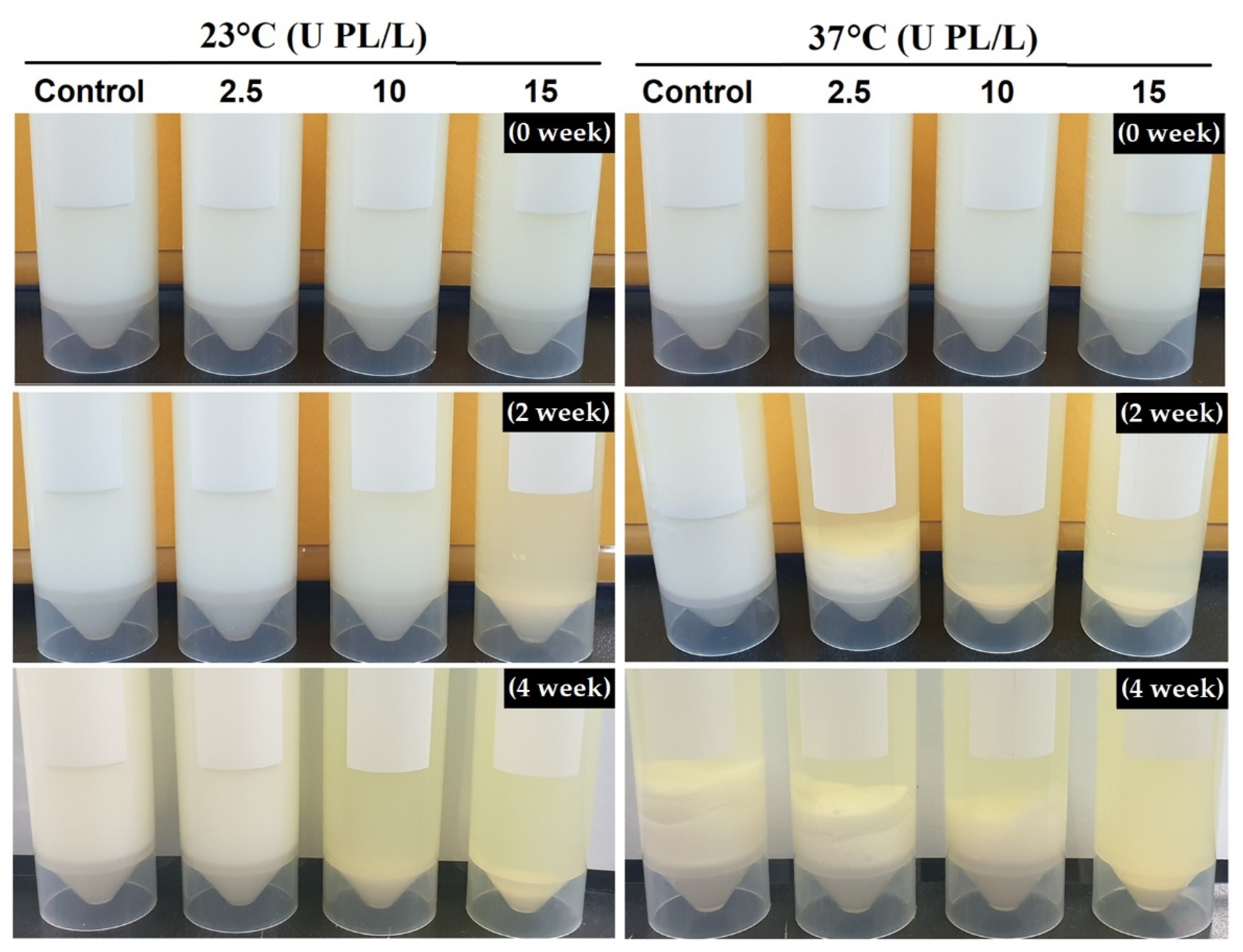
2.2. Changes in Protein Sedimentation Content of PL-Added UHT Skim Milk Stored at Different Temperatures
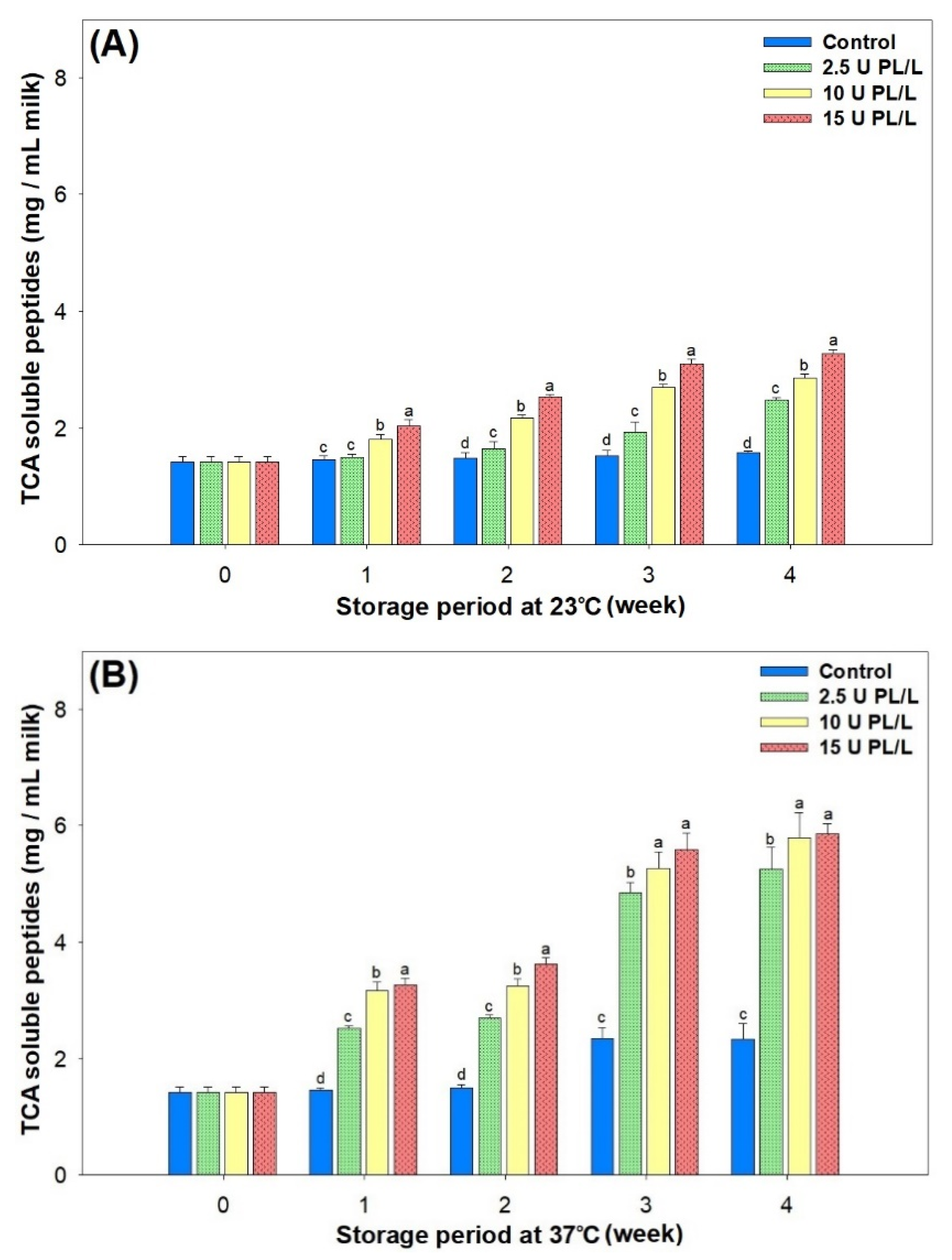
2.3. Changes in Proteolysis in PL-Added UHT Skim Milk Stored at Different Temperatures
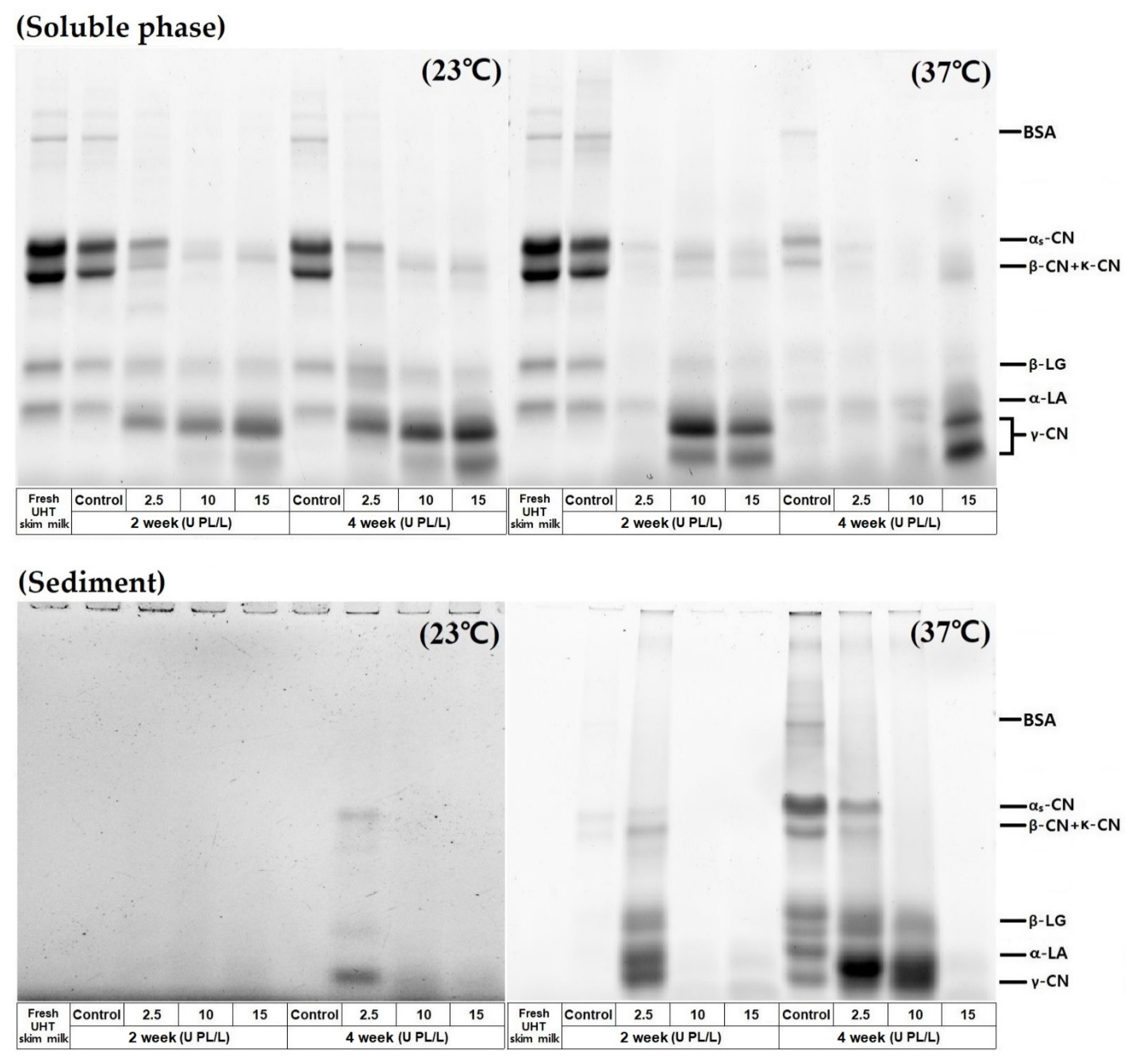
2.4. Protein Profiles of the Soluble Phase and Sediment in PL-Added UHT Skim Milk Stored at Different Temperatures
2.5. Changes in Microstructure of PL-Added UHT Skim Milk Storored at Different Temperatures
2.6. PL Concentration-Dependent Secondary Structure Changes in UHT Skim Milk during Storage
3. Materials and Methods
3.1. Preparation of UHT Skim Milk Containing Various Concentrations of PL
3.2. Particle Size Distribution
3.3. Sediment Analysis
3.4. Protein Profiles of the Soluble Phase and Sediment
3.5. Extent of Proteolysis
3.6. Transmission Eectron Microscopy (TEM)
3.7. FT-IR
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Chavan, R.S.; Chavan, S.R.; Khedkar, C.D.; Jana, A.H. UHT Milk Processing and Effect of Plasmin Activity on Shelf Life: A Review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 251–268. [Google Scholar] [CrossRef]
- Mordor Intelligence Global UHT Milk Market–Market-Growth, Trends, and Forecast (2020–2025). Available online: https://www.mordorintelligence.com/industry-reports/global-uht-milk-market (accessed on 6 November 2020).
- Kim, S.-C.; Yun, S.-Y.; Ahn, N.-H.; Kim, S.-M.; Imm, J.-Y. Effect of Homogenization Pressure on Plasmin Activity and Mechanical Stress-Induced Fat Aggregation of Commercially Sterilized Ultra High Temperature Milk during Storage. Food Sci. Anim. Resour. 2020, 40, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Datta, N.; Deeth, H.C. Age gelation of UHT milk—a review. Food Bioprod. Process. 2001, 79, 197–210. [Google Scholar] [CrossRef]
- McMahon, D.J. Age-gelation of UHT milk: Changes that occur during storage, their effect on shelf life and the mechanism by which age-gelation occurs. In Heat Treatments and Alternative Methods, IDF Symposium; International Dairy Federation: Brussels, Belgium, 1996; pp. 315–326. [Google Scholar]
- Ismail, B.S.; Nielsen, S.S. Invited review: Plasmin protease in milk: Current knowledge and relevance to dairy industry. J. Dairy Sci. 2010, 93, 4999–5009. [Google Scholar] [CrossRef] [Green Version]
- Prado, B.; Ismail, B.; Ramos, O.; Hayes, K. Thermal stability of plasminogen activators and plasminogen activation in heated milk. Int. Dairy J. 2007, 17, 1028–1033. [Google Scholar] [CrossRef]
- Stoeckel, M.; Lidolt, M.; Stressler, T.; Fischer, L.; Wenning, M.; Hinrichs, J. Heat stability of indigenous milk plasmin and proteases from Pseudomonas: A challenge in the production of ultra-high temperature milk products. Int. Dairy J. 2016, 61, 250–261. [Google Scholar] [CrossRef]
- Bastian, E.; Lo, C.; David, K. Plasminogen Activation in Cheese Milk: Influence on Swiss Cheese Ripening. J. Dairy Sci. 1997, 80, 245–251. [Google Scholar] [CrossRef]
- Mara, O.; Roupie, C.; Duffy, A.; Kelly, A.L. The Curd-forming Properties of Milk as Affected by the Action of Plasmin. Int. Dairy J. 1998, 8, 807–812. [Google Scholar] [CrossRef]
- Gazi, I.; Vilalva, I.C.; Huppertz, T. Plasmin activity and proteolysis in milk protein ingredients. Int. Dairy J. 2014, 38, 208–212. [Google Scholar] [CrossRef]
- Anema, S.G. Age Gelation, Sedimentation, and Creaming in UHT Milk: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 140–166. [Google Scholar] [CrossRef] [Green Version]
- Walstra, P.; Wouters, J.T.M.; Geurts, T.J. Dairy Science and Technology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 109–158. [Google Scholar]
- Manji, B.; Kakuda, Y. The role of protein denaturation, extent of hydrolysis and storage temperature on the mechanism of age gelation in a model system. J. Dairy Sci. 1988, 71, 1455–1463. [Google Scholar] [CrossRef]
- Malmgren, B.; Ardö, Y.; Langton, M.; Altskär, A.; Bremer, M.G.; Dejmek, P.; Paulsson, M. Changes in proteins, physical stability and structure in directly heated UHT milk during storage at different temperatures. Int. Dairy J. 2017, 71, 60–75. [Google Scholar] [CrossRef]
- Anema, S.G. Storage stability and age gelation of reconstituted ultra-high temperature skim milk. Int. Dairy J. 2017, 75, 56–67. [Google Scholar] [CrossRef]
- Gaur, V.; Schalk, J.; Anema, S.G. Sedimentation in UHT milk. Int. Dairy J. 2018, 78, 92–102. [Google Scholar] [CrossRef]
- Akkerman, M.; Johansen, L.B.; Rauh, V.; Sørensen, J.; Larsen, L.B.; Poulsen, N.A. Relationship between casein micelle size, protein composition and stability of UHT milk. Int. Dairy J. 2021, 112, 104856. [Google Scholar] [CrossRef]
- Sunds, A.V.; Rauh, V.M.; Sørensen, J.; Larsen, L.B. Maillard reaction progress in UHT milk during storage at different temperature levels and cycles. Int. Dairy J. 2018, 77, 56–64. [Google Scholar] [CrossRef]
- Rauh, V.M.; Johansen, L.B.; Ipsen, R.; Paulsson, M.; Larsen, L.B.; Hammershøj, M. Plasmin activity in UHT milk: Re-lationship between proteolysis, age gelation, and bitterness. J. Agric. Food Chem. 2014, 62, 6852–6860. [Google Scholar] [CrossRef]
- Bastian, E.D.; Brown, R.J. Plasmin in milk and dairy products: An update. Int. Dairy J. 1996, 6, 435–457. [Google Scholar] [CrossRef]
- Scollard, P.G.; Beresford, T.P.; Murphy, P.M.; Kelly, A.L. Barostability of milk plasmin activity. Le Lait 2000, 80, 609–619. [Google Scholar] [CrossRef]
- Nielsen, S.S. Plasmin System and Microbial Proteases in Milk: Characteristics, Roles, and Relationship. J. Agric. Food Chem. 2002, 50, 6628–6634. [Google Scholar] [CrossRef]
- Gaucher, I.; Mollé, D.; Gagnaire, V.; Gaucheron, F. Effects of storage temperature on physico-chemical characteristics of semi-skimmed UHT milk. Food Hydrocoll. 2008, 22, 130–143. [Google Scholar] [CrossRef]
- Huppertz, T.; Fox, P.F.; Kelly, A.L. Plasmin activity and proteolysis in high pressure-treated bovine milk. Le Lait 2004, 84, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Holland, J.W.; Gupta, R.; Deeth, H.C.; Alewood, P.F. Proteomic Analysis of Temperature-Dependent Changes in Stored UHT Milk. J. Agric. Food Chem. 2011, 59, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Raynes, J.K.; Day, L.; Crepin, P.; Horrocks, M.H.; Carver, J.A. Coaggregation of κ-casein and β-lactoglobulin produces morphologically distinct amyloid fibrils. Small 2017, 13, 1603591. [Google Scholar] [CrossRef] [PubMed]
- Morgan, P.E.; Treweek, T.M.; Lindner, R.A.; Price, W.E.; Carver, J.A. Casein Proteins as Molecular Chaperones. J. Agric. Food Chem. 2005, 53, 2670–2683. [Google Scholar] [CrossRef]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited review: Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef]
- Grewal, M.K.; Chandrapala, J.; Donkor, O.; Apostolopoulos, V.; Stojanovska, L.; Vasiljevic, T. Fourier transform infrared spectroscopy analysis of physicochemical changes in UHT milk during accelerated storage. Int. Dairy J. 2017, 66, 99–107. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. In The Protein Protocols Handbook; Humana Press: Totowa, NJ, USA, 2009; pp. 11–15. [Google Scholar]
- Qi, P.X.; Ren, D.; Xiao, Y.; Tomasula, P.M. Effect of homogenization and pasteurization on the structure and stability of whey protein in milk. J. Dairy Sci. 2015, 98, 2884–2897. [Google Scholar] [CrossRef] [Green Version]
- Carbonaro, M.; Nucara, A. Secondary structure of food proteins by Fourier transform spectroscopy in the mid-infrared region. Amino Acids 2009, 38, 679–690. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Storage Period (Week, 23 °C) | Protein Sediment (mg) | Storage Period (Week, 37 °C) | Protein Sediment (mg) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Added PL Concentration (U PL/L) | Added PL Concentration (U PL/L) | |||||||||
| Control | 2.5 | 10 | 15 | Control | 2.5 | 10 | 15 | |||
| 0 | 0.31 ± 0.01 Az | 0.31 ± 0.01 Az | 0.31 ± 0.01 Az | 0.31 ± 0.01 Az | 0 | 0.31 ± 0.01 Az | 0.31 ± 0.01 Az | 0.31 ± 0.01 Az | 0.31 ± 0.01 Az | |
| 1 | 0.43 ± 0.02 Cy | 0.48 ± 0.01 Cy | 6.39 ± 0.12 By | 7.17 ± 0.17 Aw | 1 | 0.69 ± 0.01 Cz | 21.25 ± 0.36 Ay | 6.48 ± 0.48 By | 5.90 ± 0.09 By | |
| 2 | 0.58 ± 0.01 Dx | 1.02 ± 0.01 Cx | 6.66 ± 0.20 By | 7.69 ± 0.18 Bv | 2 | 5.01 ± 0.10 Dy | 49.39 ± 0.23 Ax | 8.22 ± 0.13 Bx | 7.66 ± 0.25 Cx | |
| 3 | 0.53 ± 0.06 Dx | 3.00 ± 0.11 Cw | 9.43 ± 0.20 Aw | 5.19 ± 0.11 Cx | 3 | 70.13 ± 0.25 Bx | 71.05 ± 0.45 Aw | 47.13 ± 0.22 Cw | 11.68 ± 0.19 Dw | |
| 4 | 0.60 ± 0.02 Dx | 3.83 ± 0.03 Bv | 8.14 ± 0.03 Ax | 2.98 ± 0.08 By | 4 | 93.45 ± 1.99 Aw | 95.50 ± 0.59 Av | 80.55 ± 0.42 Bv | 14.05 ± 0.36 Cv | |
| Two-way ANOVA analysis | Significance | Two-way ANOVA analysis | Significance | |||||||
| Interactions of storage time × added PL concentration | ** | Interactions of storage time × added PL concentration | ** | |||||||
| Protein Sediment | ||
|---|---|---|
| 23 °C | 37 °C | |
| Added PL concentration | 0.615 ** | −0.358 ** |
| Storage period | 0.361 ** | 0.768 ** |
| Samples | Intermolecular β-Sheet (%) | Turns (%) | α-Helix (%) | Disordered (%) | Intramolecular β-Sheet (%) |
|---|---|---|---|---|---|
| Control | 15 | 30 | 18 | 23 | 15 |
| 2.5 U PL/L | 19 | 20 | 22 | 24 | 15 |
| 10 U PL/L | 21 | 21 | 22 | 22 | 14 |
| 15 U PL/L | 28 | 20 | 15 | 22 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, S.-Y.; Imm, J.-Y. Changes in Particle Size, Sedimentation, and Protein Microstructure of Ultra-High-Temperature Skim Milk Considering Plasmin Concentration and Storage Temperature. Molecules 2021, 26, 2339. https://doi.org/10.3390/molecules26082339
Yun S-Y, Imm J-Y. Changes in Particle Size, Sedimentation, and Protein Microstructure of Ultra-High-Temperature Skim Milk Considering Plasmin Concentration and Storage Temperature. Molecules. 2021; 26(8):2339. https://doi.org/10.3390/molecules26082339
Chicago/Turabian StyleYun, So-Yul, and Jee-Young Imm. 2021. "Changes in Particle Size, Sedimentation, and Protein Microstructure of Ultra-High-Temperature Skim Milk Considering Plasmin Concentration and Storage Temperature" Molecules 26, no. 8: 2339. https://doi.org/10.3390/molecules26082339