
Simultaneous Quantitative Analysis of Ginsenosides Isolated from the Fruit of Panax ginseng C.A. Meyer and Regulation of HO-1 Expression through EGFR Signaling Has Anti-Inflammatory and Osteogenic Induction Effects in HPDL Cells

Abstract
:1. Introduction
2. Result
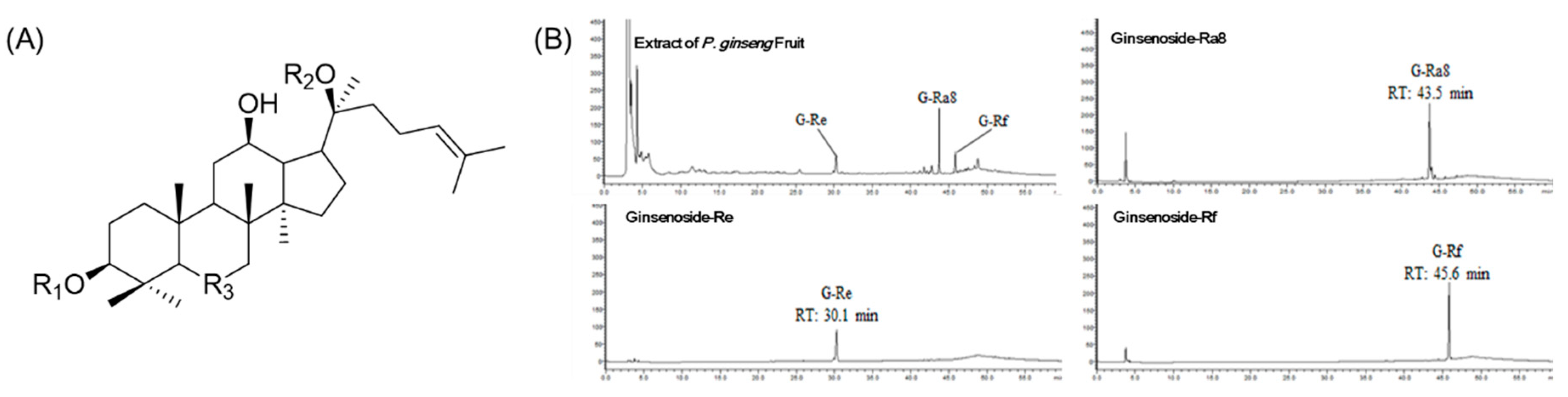
2.1. Structure and Specificity of Isolated Ginsenosides
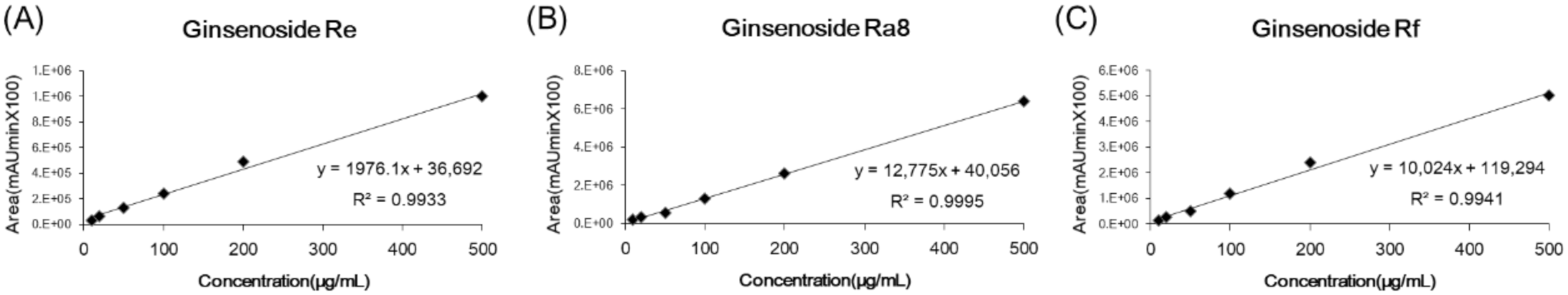
2.2. Linearity, Limit of Detection (LOD) and Limit of Quantitation (LOQ)
2.3. Precision and Accuracy of the Analysis Method
2.4. Simultaneous Quantitative Analysis of G-Re, G-Ra8, and G-Rf Isolates from P. ginseng Fruit Extracts
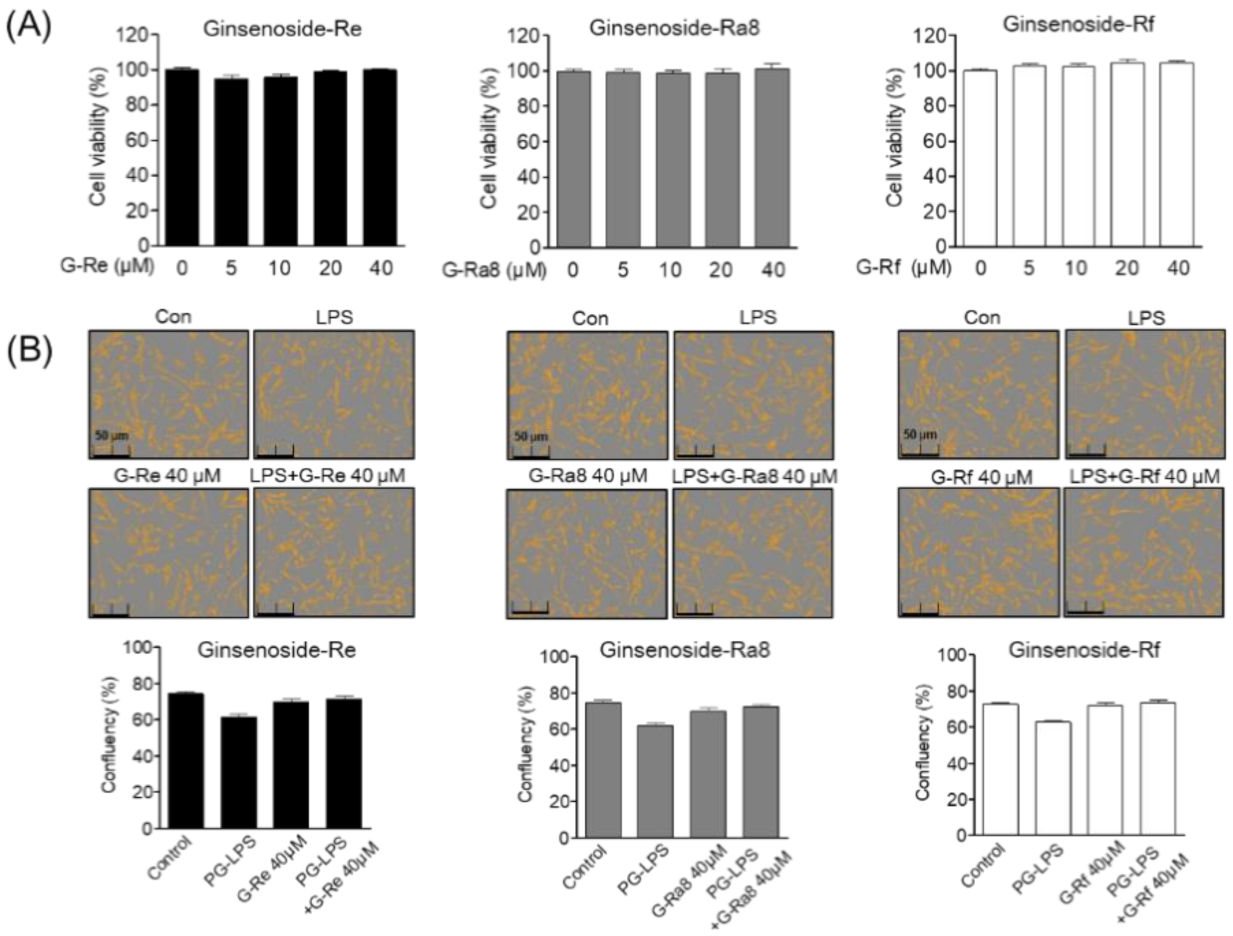
2.5. Effect of Ginsenoside G-Re, G-Ra8, and G-Rf on Human Periodontal Ligament (HPDL) Cell Viability and Confluence
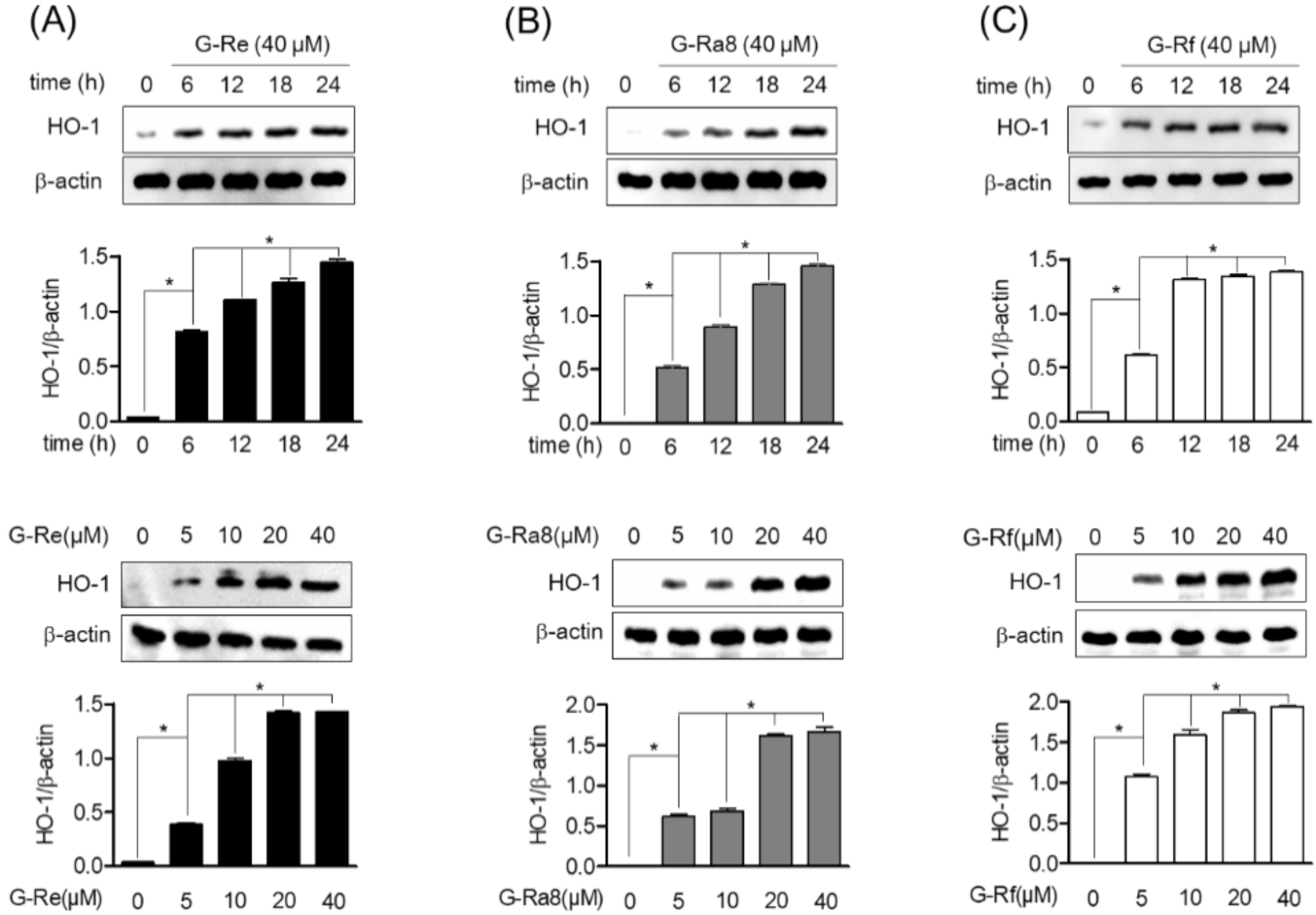
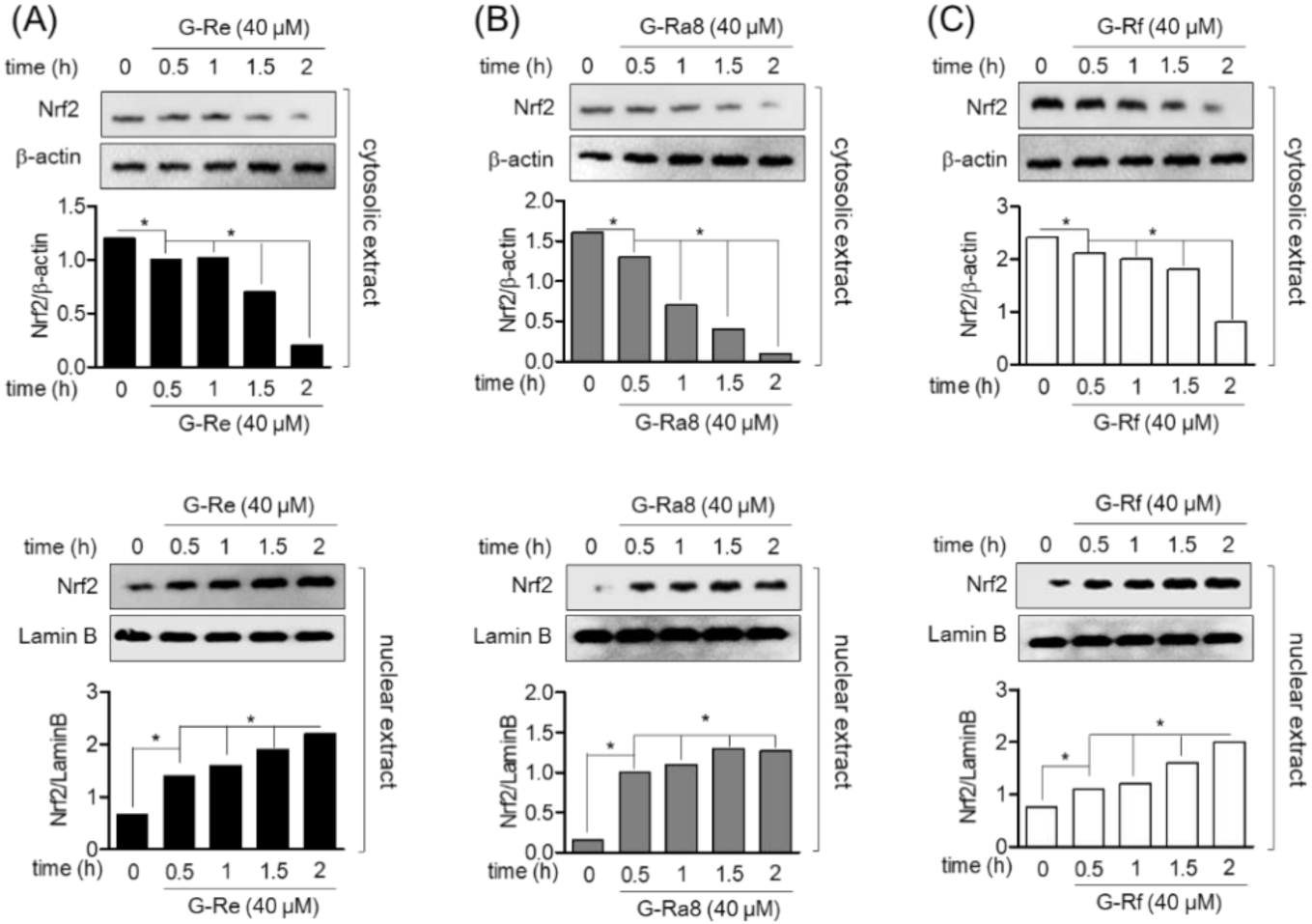
2.6. Ginsenosides Induces HO-1 Expression by Promotion of Nrf2 in HPDL Cells
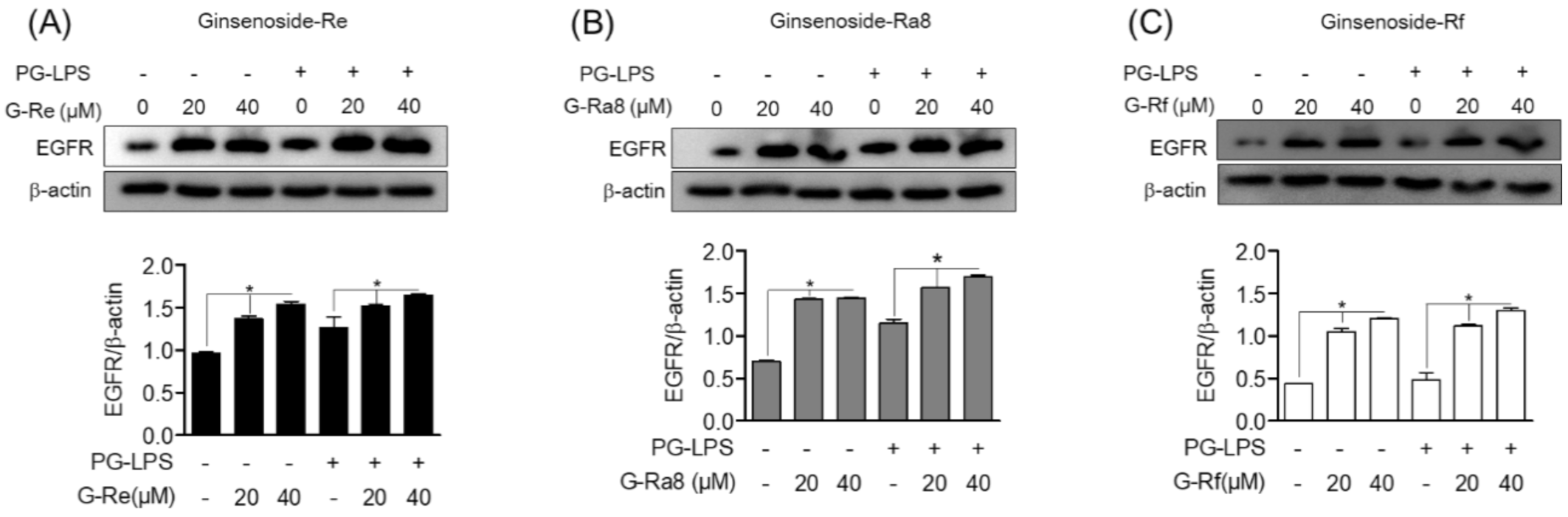
2.7. The Regulatory Effect of Ginsenosides on EGFR Expression
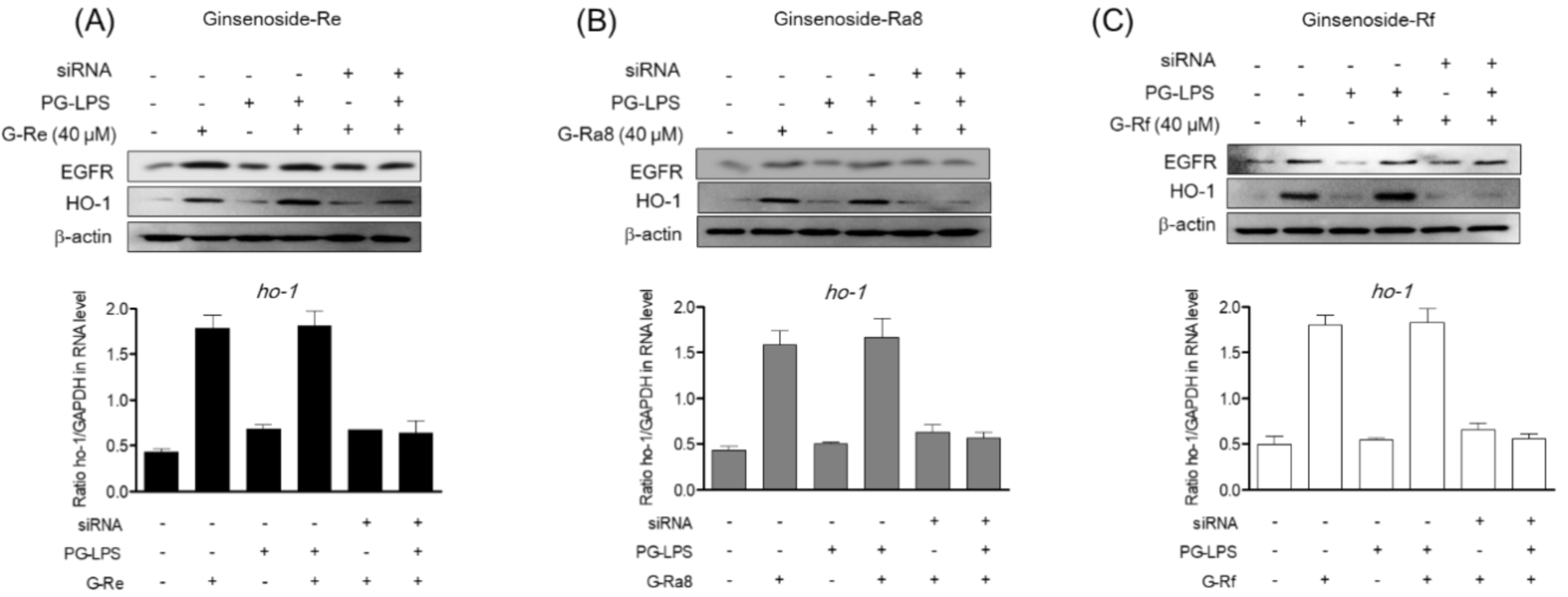
2.8. Induction of HO-1 Protein by Ginsenosides Is Regulated by EGFR
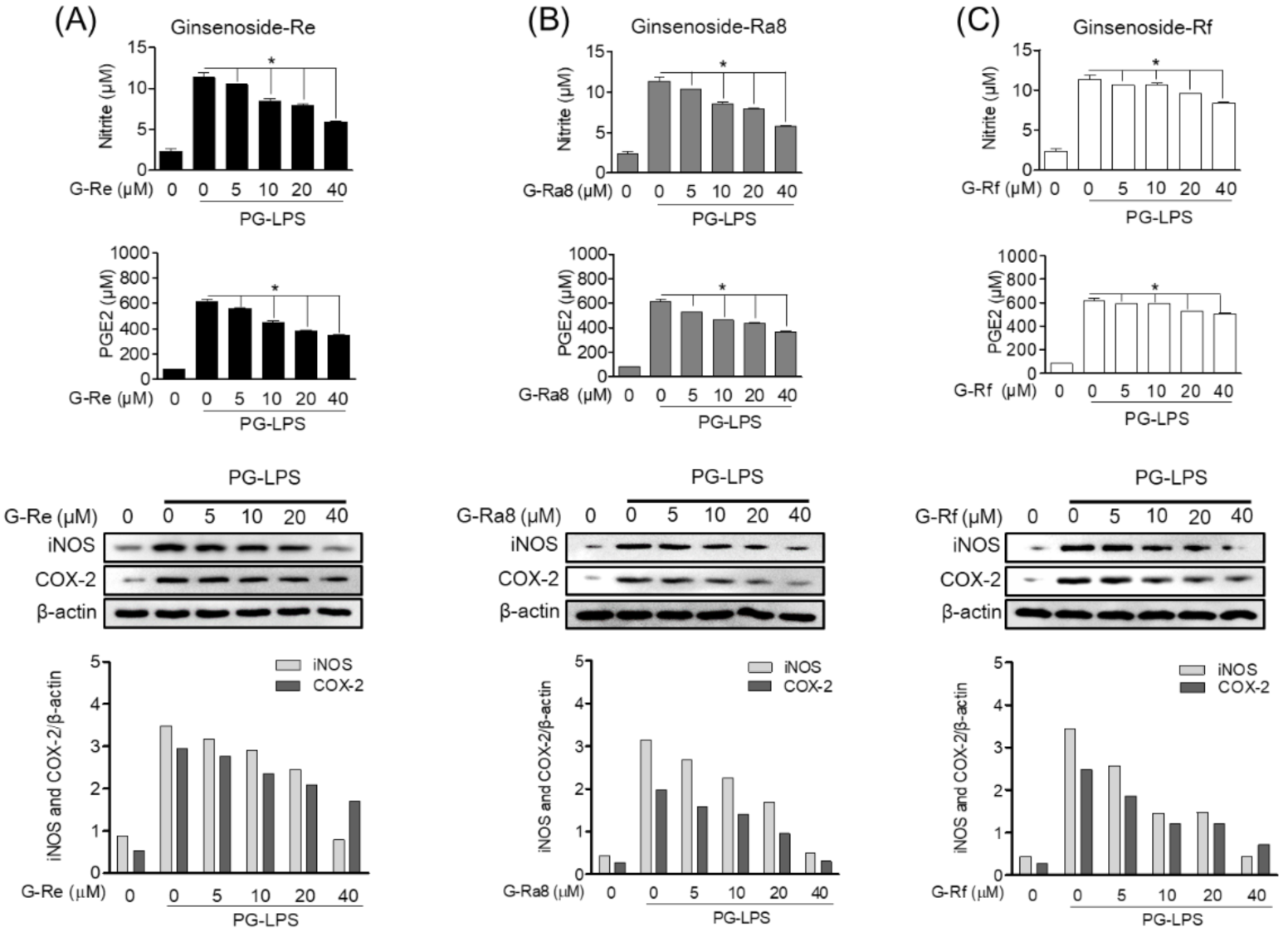
2.9. Ginsenoside Inhibits PG-LPS-Induced Pro-Inflammatory Mediator Production in HPDL Cells
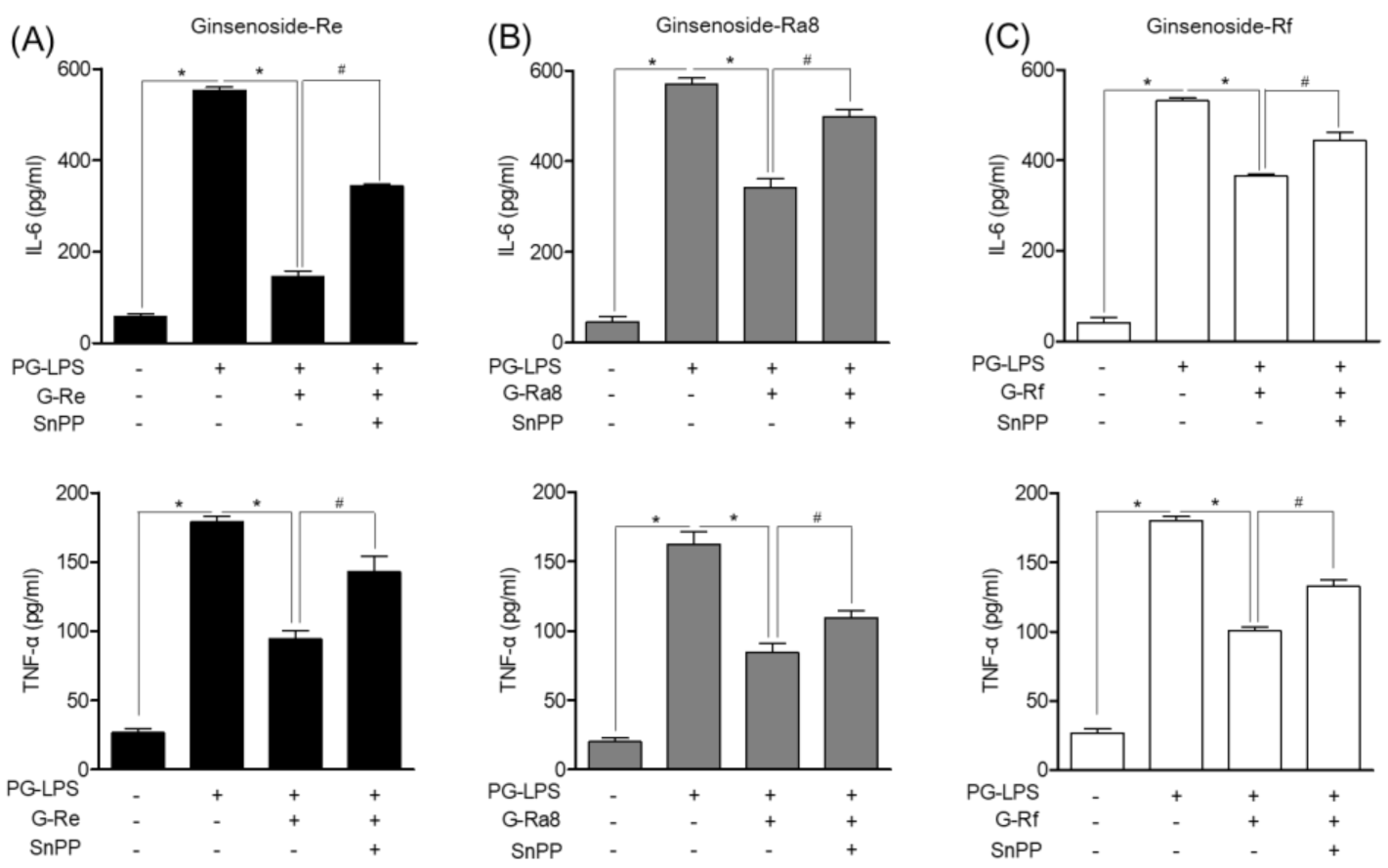
2.10. HO-1 Induced by Ginsenosides in PG-LPS-Stimulated HPDL Cells Regulates Pro-Inflammatory Cytokines
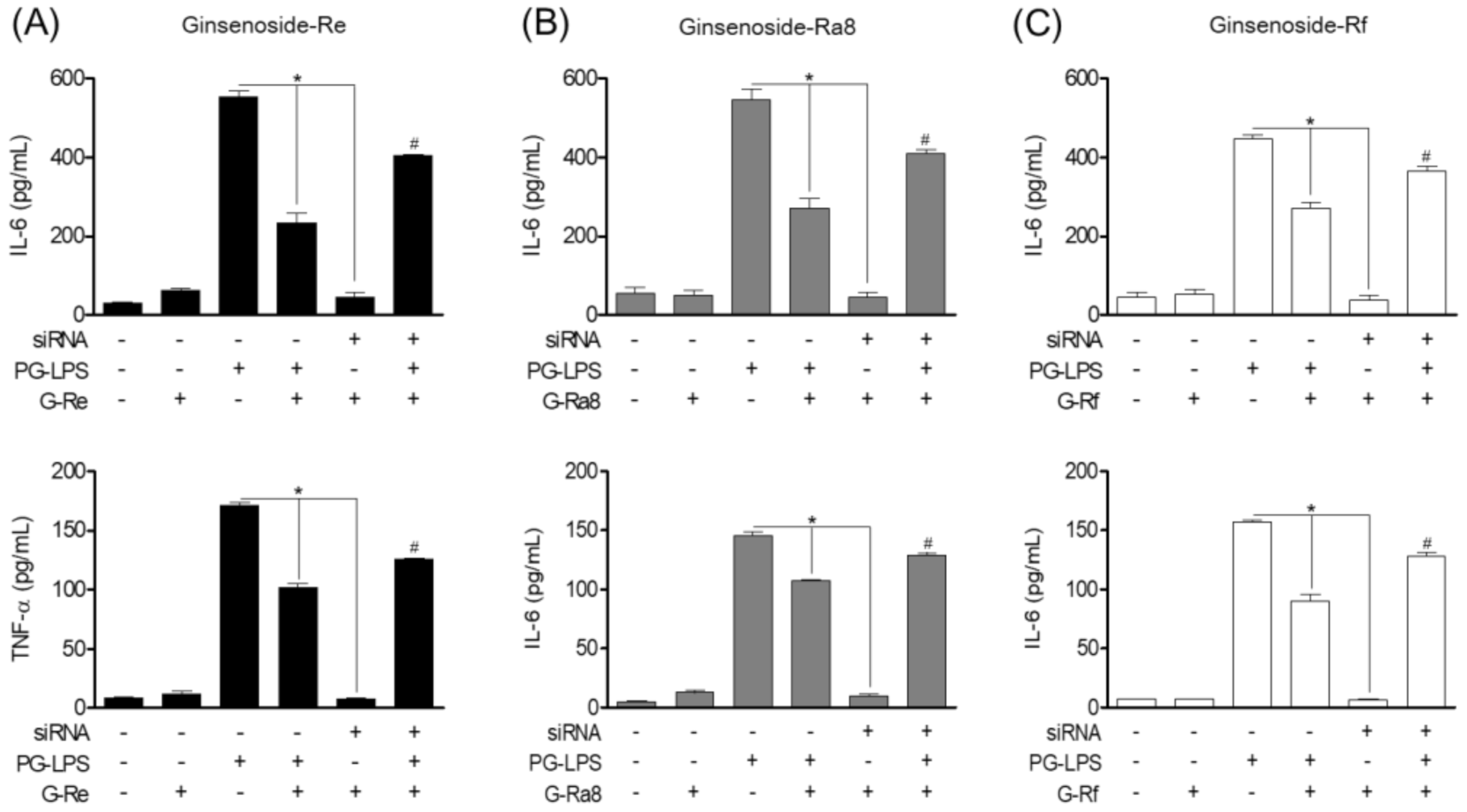
2.11. Effect of Ginsenosides Though EGFR-Mediated HO-1 Expression on Pro-Inflammatory Cytokines
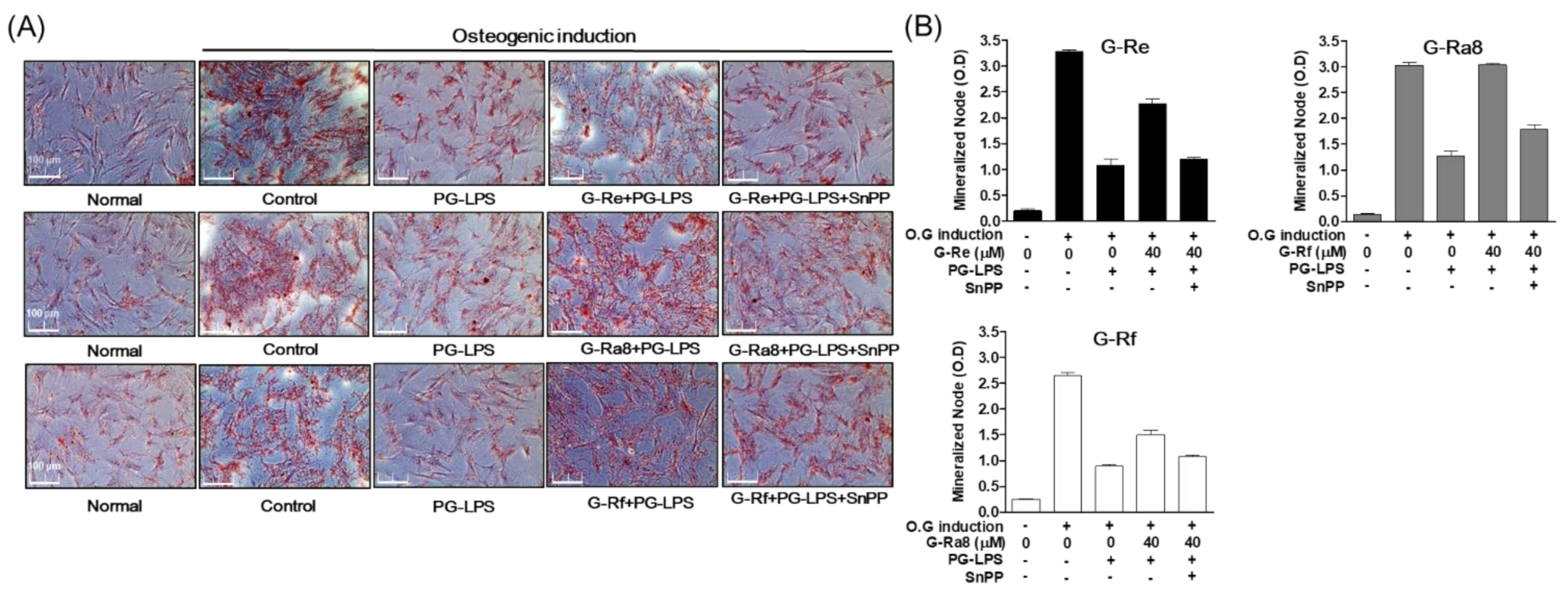
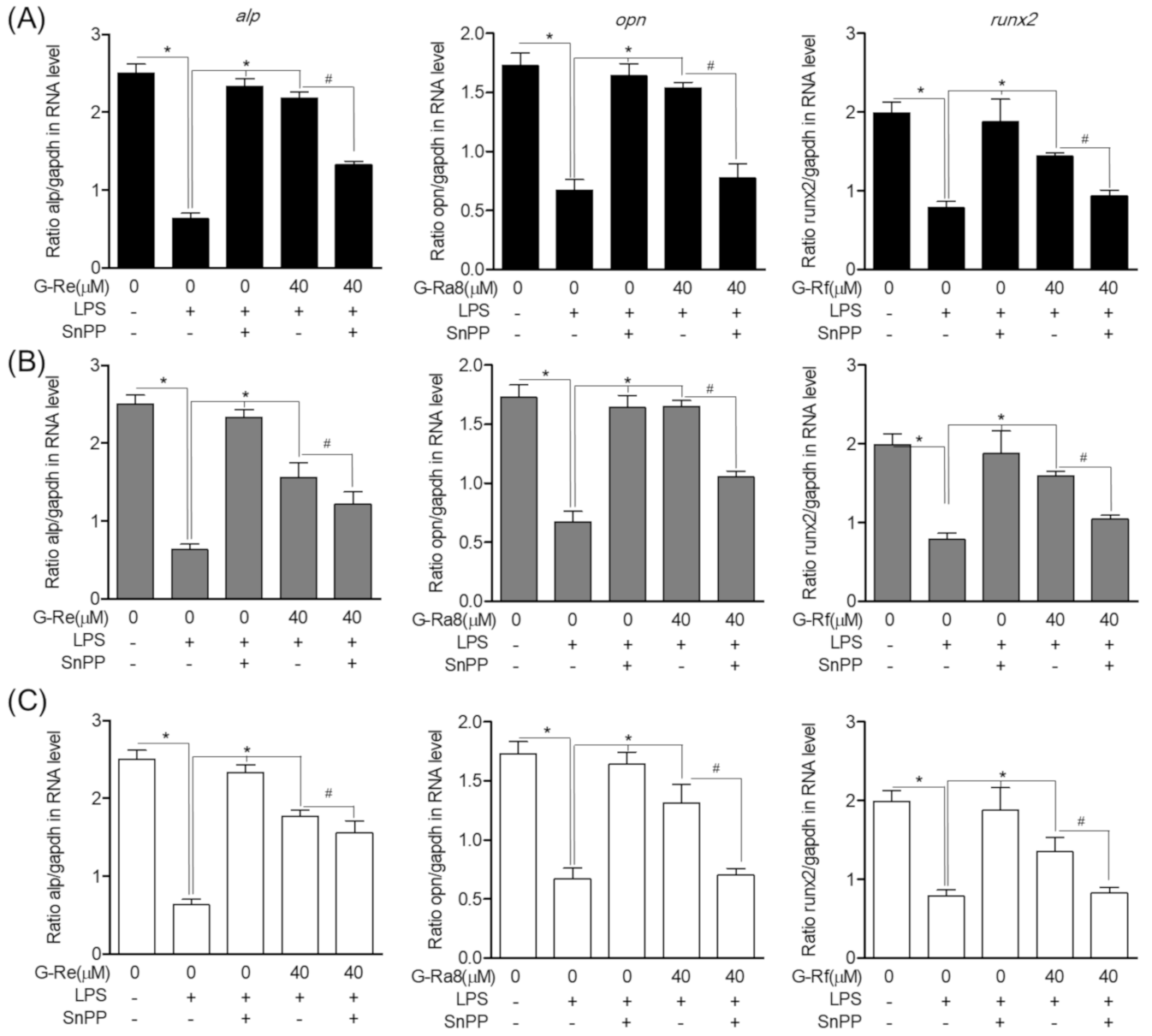
2.12. Effects of HO-1 Expression Induced by Ginsenosides on Osteoblast Differentiation and Osteoblast-Specific Gene Expression
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Extraction of Plant Material and Ginsenosides Isolation
4.3. High-Performance Liquid Chromatography (HPLC) Analysis
4.4. Validation of HPLC Analysis Method
4.5. Inter-Day and Intra-Day Tests for Method Validation
4.6. Preparation of Human Periodontal Ligament Cells
4.7. The Cell Viability and Coefficient Assays of HPDL Cells
4.8. Nitric Oxide and Cytokines Production Assay
4.9. Evaluation of Mineralization Analysis by Ginsenosides
4.10. Western Blot Analysis
4.11. Real-Time Quantitative PCR
4.12. Small Interfering RNA (siRNA)-Induced EGFR Gene Silencing
4.13. Statistical Analysis
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Joanne, M.L. Dental plaque revisited: Bacteria associated with periodontal disease. J. N. Z. Soc. Periodontol. 2004, 87, 7–21. [Google Scholar]
- Botero, J.E.; Rosing, C.K.; Duque, A.; Jaramillo, A.; Contreas, A. Periodontal disease in children and adolescent of Latin America. Periodontology 2000 2015, 67, 34–57. [Google Scholar] [CrossRef] [PubMed]
- Albandar, J.M. Aggressive periodontitis: Case definition and diagnostic criteria. Periodontology 2000 2014, 65, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Walter, J.L. Microbiology of Dental Decay and Periodontal Disease. In Medical Microbiology, 4th ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; Chapter 99. [Google Scholar]
- Cho, M.I.; Garant, P.R. Development and general structure of the periodontium. Periodontology 2000 2000, 24, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Coyac, B.R.; Detzen, L.; Doucet, P.; Baroukh, B.; Llorens, A.; Bonnaure, M.M.; Gosset, M.; Barritault, D.; Colombier, M.L.; Saffar, J.L. Periodontal reconstruction by heparan sulfate mimetic-based matrix therapy in Porphyromonas gingivalis-infected mice. Heliyon 2018, 4, e00719. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Takanche, J.S.; Yun, B.S.; Yi, H.K. Anti-inflammatory character of Phelligridin D modulates periodontal regeneration in lipopolysaccharide-induced human periodontal ligament cells. J. Periodontal. Res. 2018, 53, 816–824. [Google Scholar] [CrossRef]
- Walker, C.B. The acquisition of antibiotic resistance in the periodontal microflora. Periodontology 2000 1996, 10, 79–88. [Google Scholar] [CrossRef]
- Lindhe, J.; Haffajee, A.D.; Socransky, S.S. Progression of periodontal disease in adult subjects in the absence of periodontal therapy. J. Clin. Periodontol. 1983, 10, 433–442. [Google Scholar] [CrossRef]
- Eija, K.; Mervi, G.; Ulvi, K.G. Periodontitis: A Multifaceted Disease of Tooth-Supporting Tissues. J. Clin. Med. 2019, 8, 1135. [Google Scholar]
- Lee, I.T.; Luo, S.F.; Lee, C.W.; Wang, S.W.; Lin, C.C.; Chang, C.C.; Chen, Y.L.; Chau, L.Y.; Yang, C.M. Overexpression of HO-1 protects against TNF-a-mediated airway inflammation by down-regulation of TNFR1-dependent oxidative stress. Am. J. Pathol. 2009, 175, 519–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, I.T.; Wang, S.W.; Lee, C.W.; Chang, C.C.; Lin, C.C.; Luo, S.F.; Yang, C.M. Lipoteichoic acid induces HO-1 expression via the TLR2/MyD88/c-Src/NADPH oxidase pathway and Nrf2 in human tracheal smooth muscle cells. J. Immunol. 2008, 181, 5098–5110. [Google Scholar] [CrossRef] [Green Version]
- Sato, A.Y.; Tu, X.; McAndrews, K.A.; Plotkin, L.I.; Bellido, T. Prevention of glucocorticoid induced apoptosis of osteoblasts and osteocytes by protecting against endoplasmic reticulum (ER) stress in vitro and in vivo in female mice. Bone 2015, 73, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Arai, M.; Shibata, Y.; Pugdee, K.; Abiko, Y.; Ogata, K. Effects of reactive oxygen species (ROS) on antioxidant system and osteoblastic differentiation in MC3T3-E1 cells. IUBMB 2007, 59, 27–33. [Google Scholar] [CrossRef]
- Tjabringa, G.S.; Aarbou, J.; Ninaber, D.K.; Drijfhout, J.W.; Drijfhout, J.W.; SØrensen, O.E.; Borragaard, N.; Rabe, K.F.; Hiemstra, P.S. The antimicrobial peptide LL-37 activates innate immunity at the airway epithelial surface by transactivation of the epidermal growth factor receptor. J. Immunol. 2003, 171, 6690–6696. [Google Scholar] [CrossRef] [Green Version]
- Koff, J.L.; Shao, M.X.; Kim, S.; Ueki, I.F.; Nadel, J.A. Pseudomonas lipopolysaccharide accelerates wound repair via activation of a novel epithelial cell signaling cascade. J. Immunol. 2006, 177, 8693–8700. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, I. Epidermal growth factor receptor in relation to tumor development: EGFR-targeted anticancer therapy. FEBS. J. 2010, 277, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Chunhuia, Y.; Wenjuna, C.; Huia, W.; Liquana, S.; Changweia, Z.; Tianzhub, Z.; Wenhai, Z. Pilose antler peptide protects osteoblasts from inflammatory andoxidative injury through EGF/EGFR signaling. Int. J. Biol. Macromol. 2017, 99, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Sun, Z.; Tong, G.; Yi, W.; Ma, L.; Zhao, B.; Cheng, L.; Zhang, J.; Cao, F.; Yi, D. a-Lipoic acid reduces infarct size and preserves cardiac function in rat myocardial ischemia/reperfusion injury through activation of PI3K/Akt/Nrf2 pathway. PLoS ONE 2013, 8, 58371. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.M.; Lin, C.C.; Lee, I.T.; Hsu, C.K.; Tai, Y.C.; Hsieh, H.L.; Chi, P.L.; Hsiao, L.D. c-Src-Dependent Transactivation of EGFR Mediates CORM-2-Induced HO-1 Expression in Human Tracheal Smooth Muscle Cells. J. Cell. Physiol. 2015, 230, 2351–2361. [Google Scholar] [CrossRef]
- Yoon, S.J.; Park, J.Y.; Choi, S.; Lee, J.B.; Jung, H.; Kim, T.D.; Yoon, S.R.; Choi, I.; Shim, S.; Park, Y.J. Ginsenoside Rg3 regulates S-nitrosylation of the NLRP3 inflammasome via suppression of iNOS. Biochem. Biophys. Res. Commun. 2015, 463, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- De Smet, P.A. Herbal remedies. N. Engl. J. Med. 2002, 347, 2046–2056. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Chen, Y.; Ye, Y. Ginsenoside Re and Notoginsenoside R1: Immunologic Adjuvants with Low Haemolytic Effect. Chem. Biodivers. 2006, 3, 718–726. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; In, G.; Han, S.T.; Lee, M.W.; Kim, S.Y.; Kim, K.T.; Cho, B.G.; Han, G.H.; Chang, I.M. Simultaneous determination of 30 ginsenosides in Panax ginseng preparations using ultra performance liquid chromatography. J. Ginseng. Res. 2013, 37, 457–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assuma, R.; Oates, T.; Cochran, D.; Amar, S.; Graves, D.T. IL-1 and TNF antagonists inhibit the inflammatory response and bone loss in experimental periodontitis. J. Immunol. 1998, 160, 403–409. [Google Scholar] [PubMed]
- Hana, D.; Gub, X.; Gao, J.; Wang, Z.; Liua, G.; Barkemad, H.W.; Han, B. Chlorogenic acid promotes the Nrf2/HO-1 anti-oxidative pathway by activating p21Waf1/Cip1 to resist dexamethasone-induced apoptosis in osteoblastic cells. Free Radic. Biol. Med. 2019, 137, 1–12. [Google Scholar] [CrossRef]
- Fontana, A.O.; Piffaretti, D.; Marchi, F.; Burgio, F.; Faia-Torres, A.B.; Paganetti, P.; Pinton, S.; Pieles, U.; Reinert, M. Epithelial growth factor receptor expression influences 5-ALA induced glioblastoma fluorescence. J. Neurooncol. 2017, 133, 497–507. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.G.; In, S.J.; Jung, Y.J.; Cha, B.J.; Lee, D.Y.; Kim, Y.B.; Yeom, M.; Baek, N.I. Re-evaluation of physicochemical and NMR data of triol ginsenosides Re, Rf, Rg2, and 20-gluco-Rf from Panax ginseng roots. J. Ginseng Res. 2014, 38, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | R1 | R2 | R3 |
|---|---|---|---|
| 1 (G-Re) | H | O-Glc | Glc(2,1)Rha |
| 2 (G-Ra8) | Glc(2,1)Glc-4-butenoly | - | Glc(6,1)Ara(f) |
| 3 (G-Rf) | H | H | Glc(2,1)Glc |
| Linear Range (μg/mL) | Compounds | Regression Equation | R2 | LOD (μg/mL) | LOQ (μg/mL) |
|---|---|---|---|---|---|
| 10–500 | 1 (G-Re) | y = 1976.1x + 36,692 | 0.9933 | 0.37 | 1.13 |
| 2 (G-Ra8) | y = 12,775x + 40,056 | 0.9995 | 0.69 | 2.08 | |
| 3 (G-Rf) | y = 10,024x + 119,294 | 0.9941 | 0.23 | 0.70 |
| Compounds | Concentration (μg/mL) | Intra-Day (n = 3) | Inter-Day (n = 3) | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SD | RSD (%) | Accuracy | Mean ± SD | RSD (%) | Accuracy | ||
| G-Re | 25 | 23.41 ± 0.58 | 2.50 | 93.6 | 24.03 ± 0.22 | 0.92 | 96.1 |
| 125 | 128.56 ± 2.92 | 2.27 | 102.8 | 126.88 ± 1.46 | 1.15 | 101.5 | |
| 250 | 252.11 ± 1.42 | 1.16 | 100.8 | 249.08 ± 0.29 | 0.12 | 99.6 | |
| 25 | 24.69 ± 0.58 | 2.37 | 98.8 | 25.06 ± 0.22 | 0.88 | 100.2 | |
| G-Ra8 | 125 | 136.61 ± 1.21 | 0.96 | 101.3 | 126.24 ± 1.46 | 1.16 | 101.0 |
| 250 | 228.33 ± 1.20 | 0.52 | 91.3 | 224.16 ± 0.29 | 0.13 | 89.7 | |
| 25 | 27.43 ± 0.27 | 1.23 | 109.7 | 27.49 ± 0.07 | 0.24 | 110.0 | |
| G-Rf | 125 | 127.33 ± 2.01 | 0.59 | 101.9 | 128.65 ± 0.76 | 0.56 | 102.9 |
| 250 | 272.09 ± 4.75 | 1.75 | 108.8 | 265.50 ± 3.49 | 1.31 | 106.2 | |
| Sample | Ginsenoside-Re | Ginsenoside-Ra8 | Ginsenoside-Rf |
|---|---|---|---|
| Contents (mg/g) | 1.01 ± 0.03 | 0.33 ± 0.01 | 0.55 ± 0.04 |
| Parameters | Conditions | ||
|---|---|---|---|
| Analytical column | Phenomenex C18 (4.6 × 250 mm) | ||
| HPLC detector | Diode array detector (203 nm) | ||
| Solvent A: Water Solvent B: ACN | |||
| Mobile phase | Final time | Solvent | |
| (min) | A (%) | B (%) | |
| 0 | 83 | 17 | |
| 20 | 75 | 25 | |
| 38 | 58 | 42 | |
| 60 | 83 | 17 | |
| Flow rate | 0.8 mL/min | ||
| Column oven temperature | 30 °C | ||
| Injection volume | 10 μL | ||
| Run time | 60 min | ||
| Target Gene | Sequence (5′→3′) | Accession Number | |
|---|---|---|---|
| ho-1 | Forward | CCAGGCAGAGAATGCTGAGTTC | NM_002133 |
| Reverse | AAGACTGGGCTCTCCTTGTTGC | ||
| alp | Forward | TGCAGTACGAGCTGAACAGG | NM_000478 |
| Reverse | GTCAATTCTGCCTCCTTCCA | ||
| opn | Forward | TCAGCTGGATGACCAGAGTG | NM_001040060 |
| Reverse | TTGGGGTCTACAACCAGCAT | ||
| runx2 | Forward | TCTTAGAACAAATTCTGCCCTTT | NM_001024630.3 |
| Reverse | TGCTTTGGTCTTGAAATCACA | ||
| gapdh | Forward | TGTTCGTCATGGGTGTGAAC | NM_002046 |
| Reverse | GTCTTCTGGGTGGCAGTGAT | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, E.-N.; Kaygusuz, O.; Lee, H.-S.; Jeong, G.-S. Simultaneous Quantitative Analysis of Ginsenosides Isolated from the Fruit of Panax ginseng C.A. Meyer and Regulation of HO-1 Expression through EGFR Signaling Has Anti-Inflammatory and Osteogenic Induction Effects in HPDL Cells. Molecules 2021, 26, 2092. https://doi.org/10.3390/molecules26072092
Kim E-N, Kaygusuz O, Lee H-S, Jeong G-S. Simultaneous Quantitative Analysis of Ginsenosides Isolated from the Fruit of Panax ginseng C.A. Meyer and Regulation of HO-1 Expression through EGFR Signaling Has Anti-Inflammatory and Osteogenic Induction Effects in HPDL Cells. Molecules. 2021; 26(7):2092. https://doi.org/10.3390/molecules26072092
Chicago/Turabian StyleKim, Eun-Nam, Oryon Kaygusuz, Hyun-Su Lee, and Gil-Saeng Jeong. 2021. "Simultaneous Quantitative Analysis of Ginsenosides Isolated from the Fruit of Panax ginseng C.A. Meyer and Regulation of HO-1 Expression through EGFR Signaling Has Anti-Inflammatory and Osteogenic Induction Effects in HPDL Cells" Molecules 26, no. 7: 2092. https://doi.org/10.3390/molecules26072092