
Inhibitory Effect of LGS and ODE Isolated from the Twigs of Syringa oblata subsp. dilatata on RANKL-Induced Osteoclastogenesis in Macrophage Cells



Abstract
:1. Introduction
2. Result
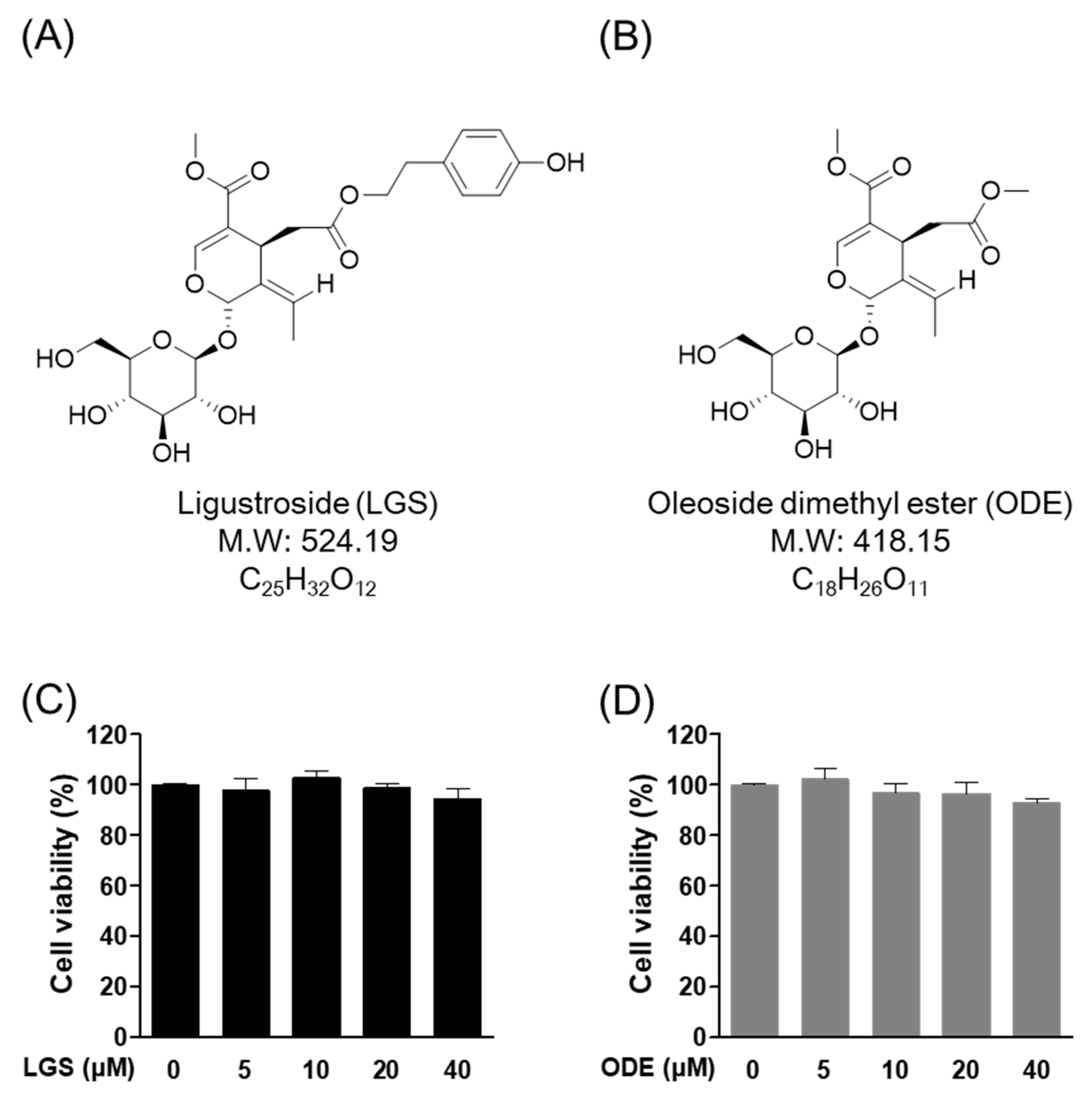
2.1. Effect of LGS and ODE on Cytotoxicity in RAW264.7 Cells
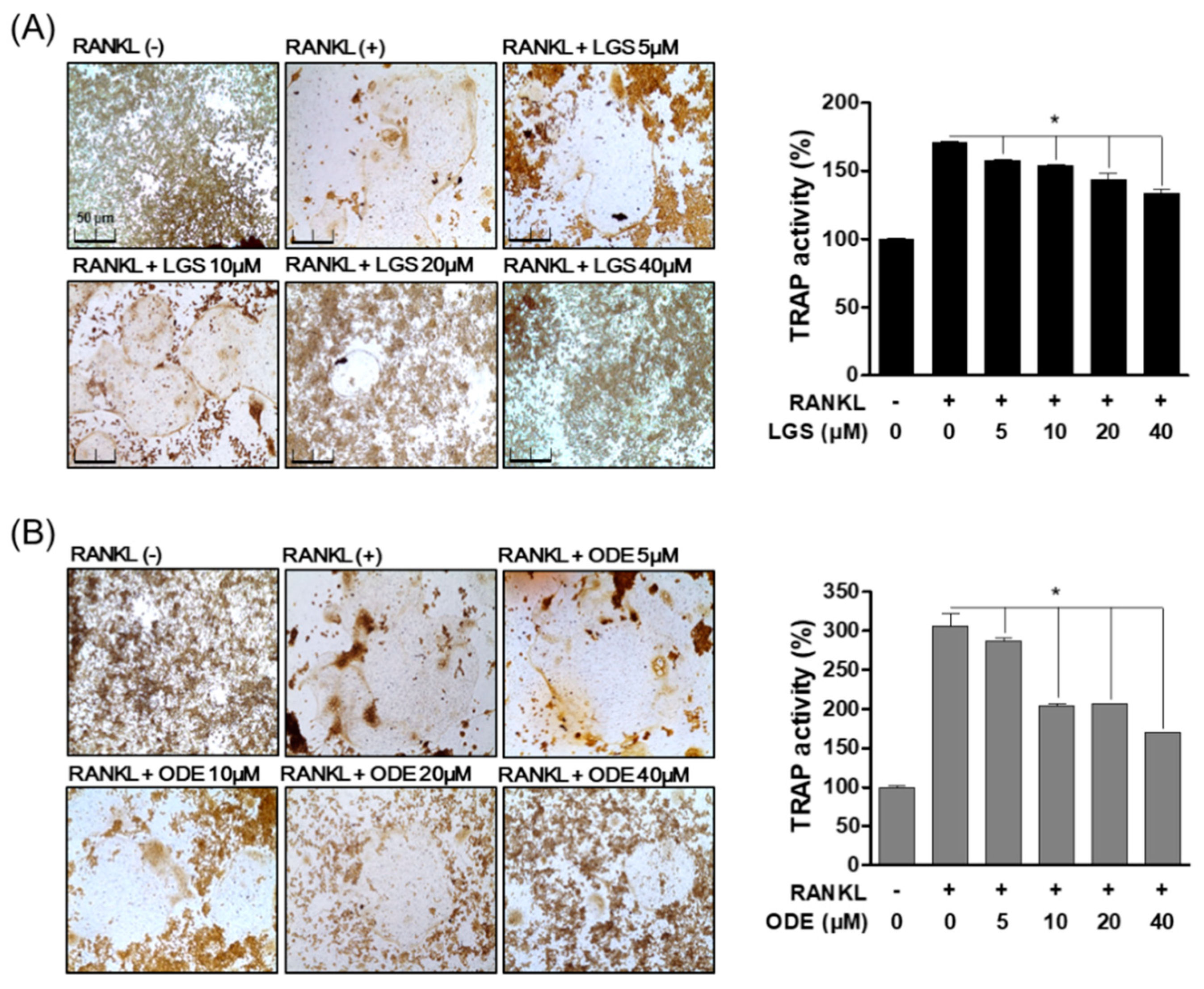
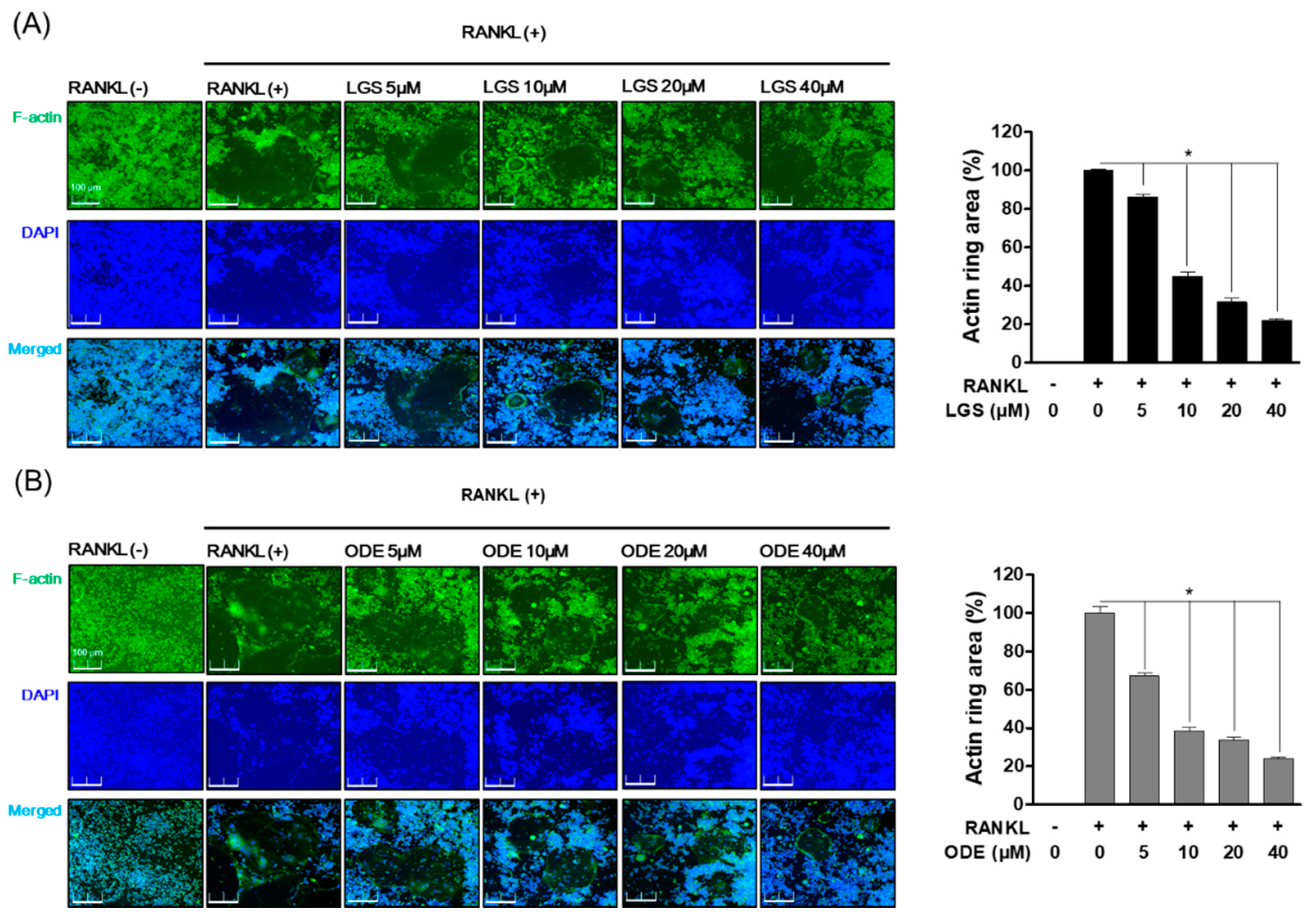
2.2. Inhibitory Effect of LGS and ODE on Actin Ring Structure and TRAP Activity during RANKL-Induced Osteoclast Formation
2.3. LGS and ODE Inhibit Bone Resorption by RANKL-Induced Osteoclasts
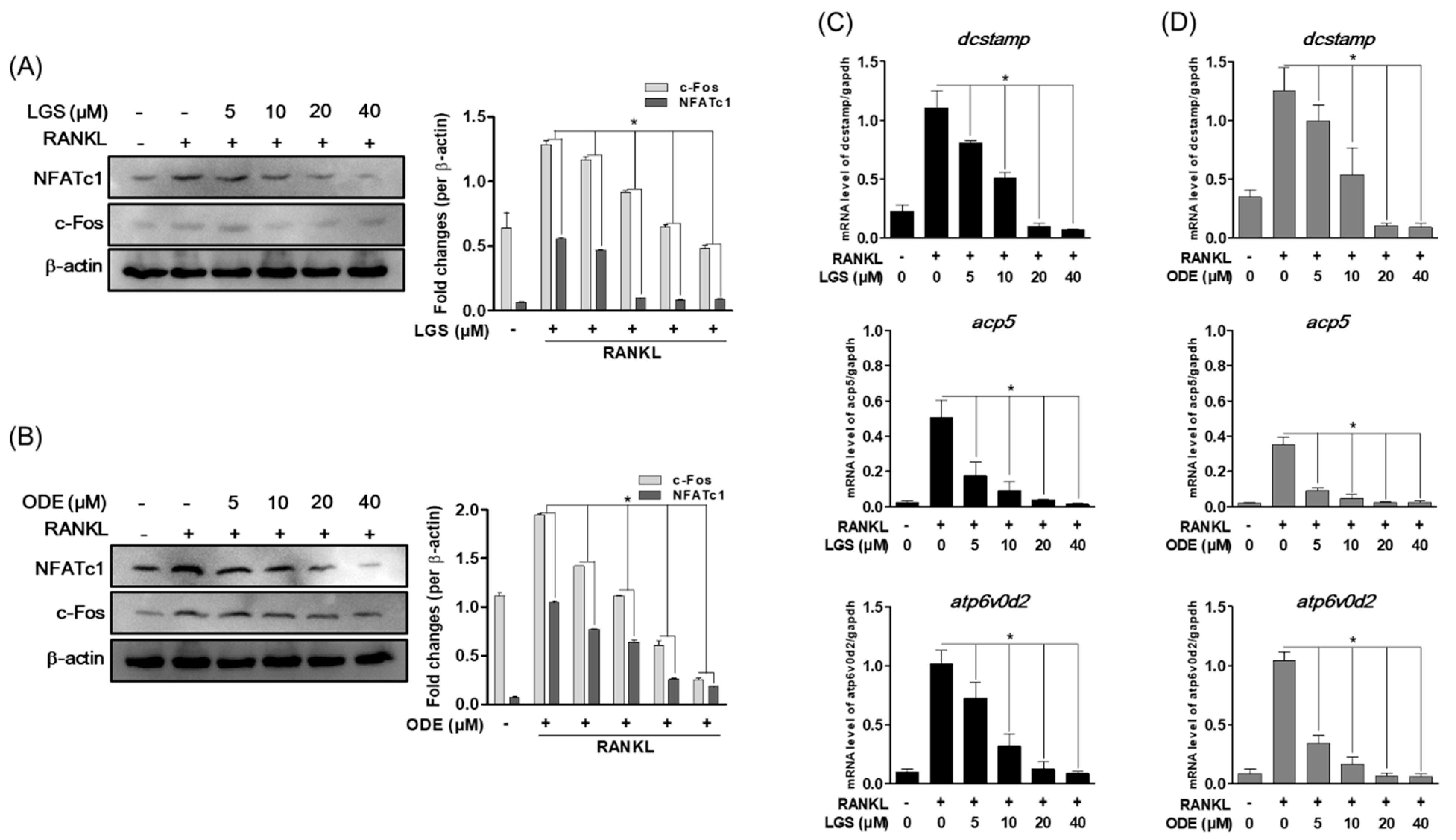
2.4. LGS and ODE Inhibit RANKL-Induced Osteoclast Formation Master Transcription Factor and Osteoclast-Specific Gene Expression
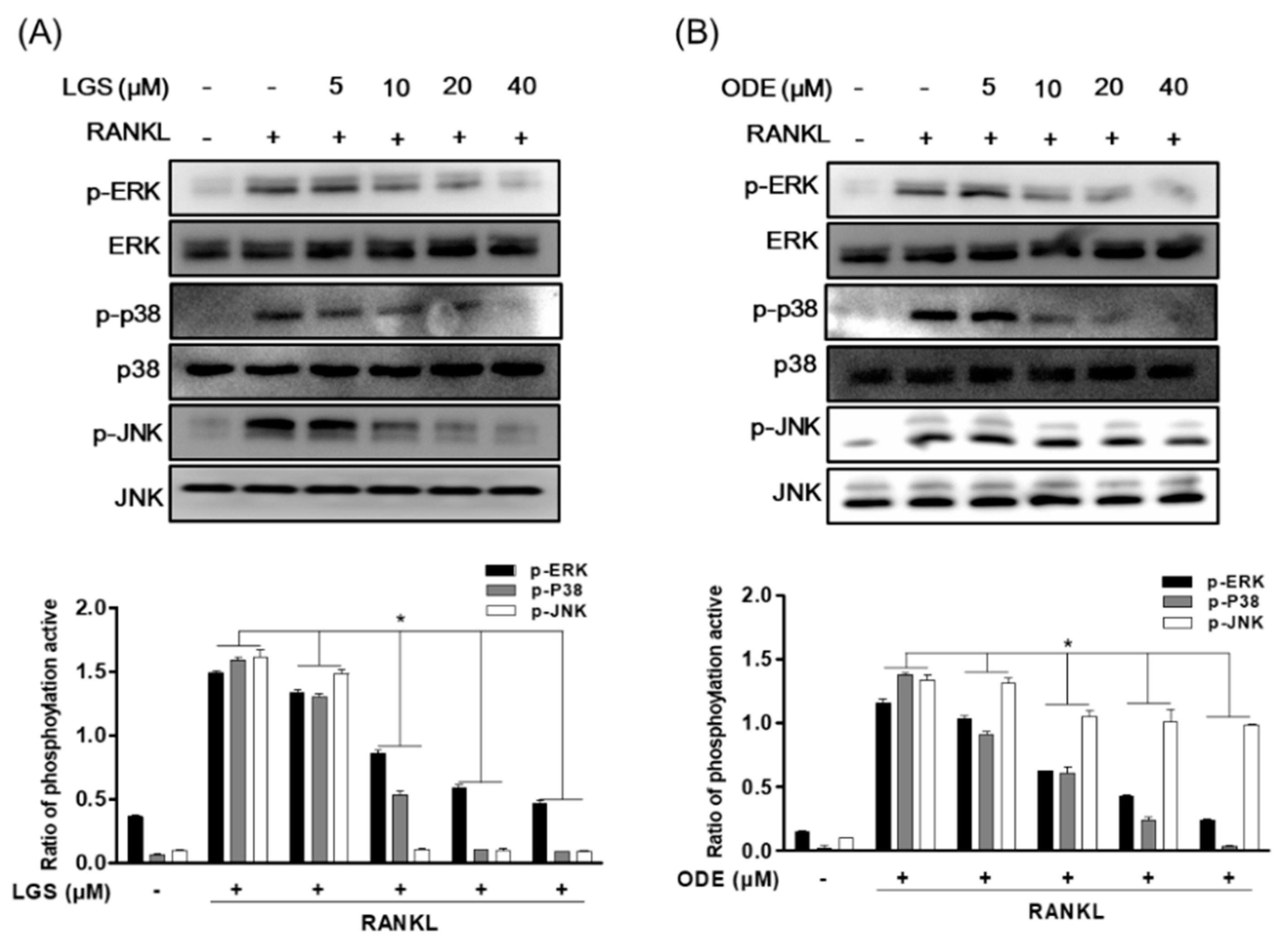
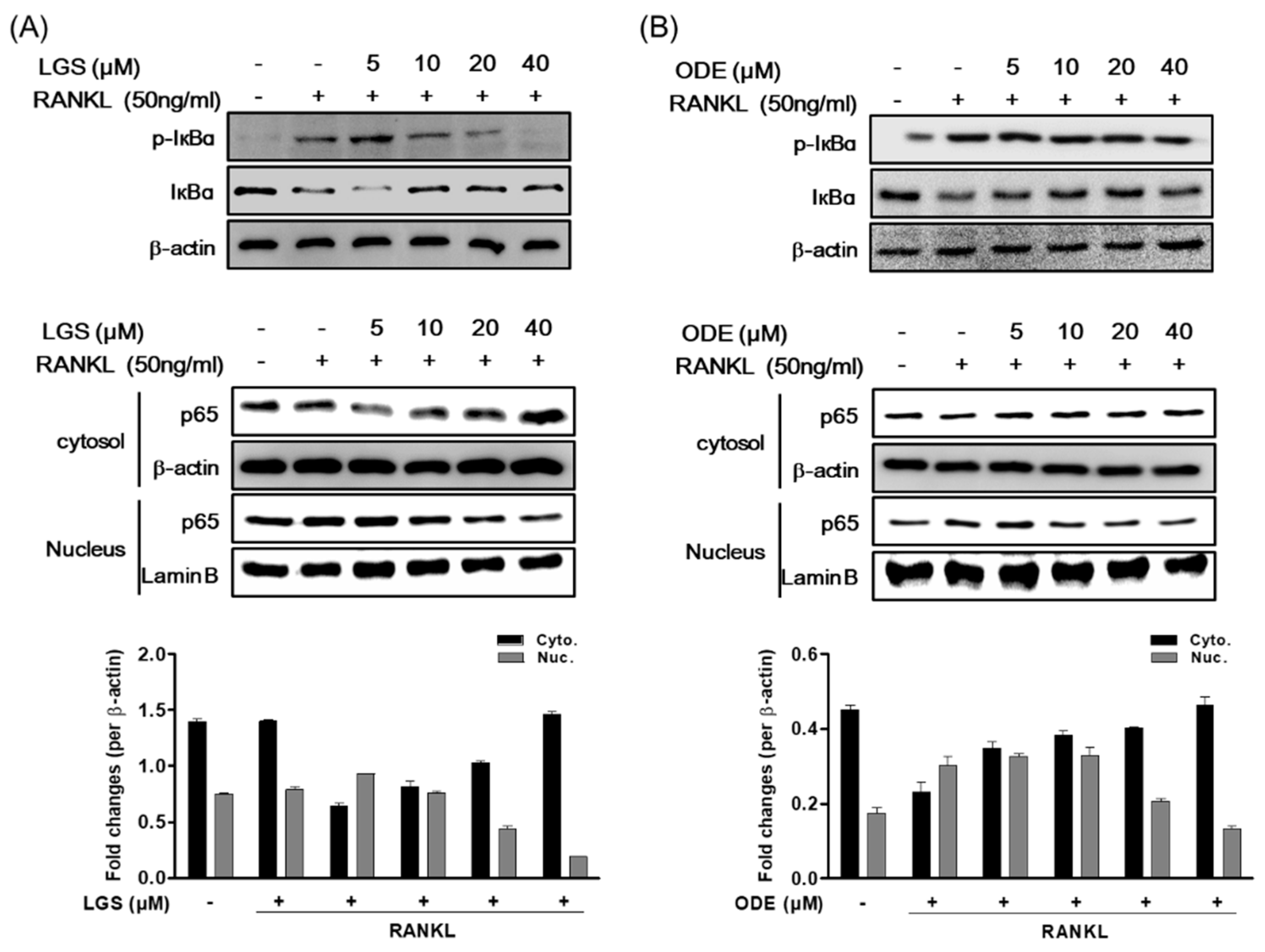
2.5. LGS and ODE Inhibit the Pathways of MAPK and NF-ĸB in Osteoclastogenesis Induced by RANKL
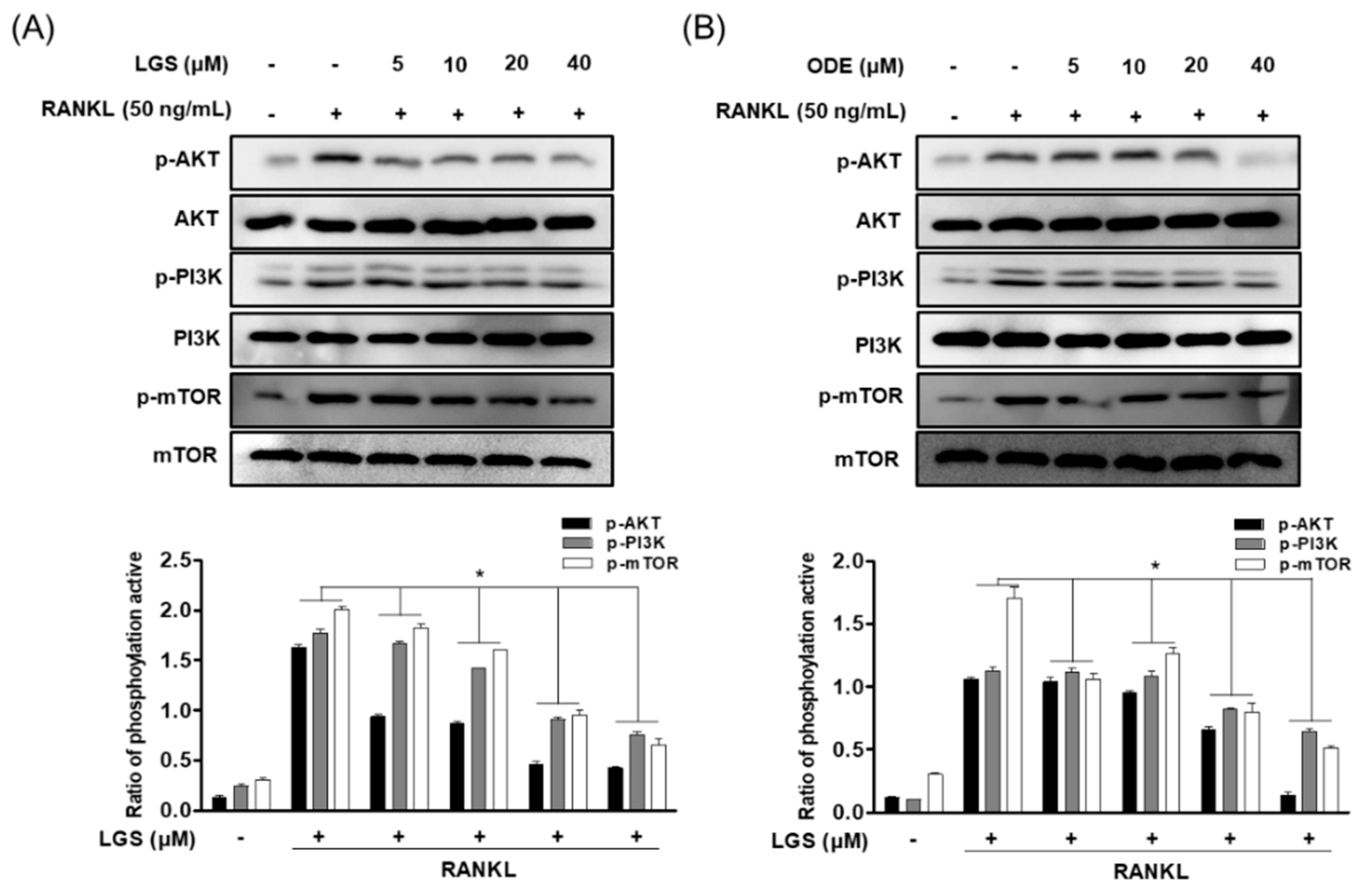
2.6. LGS and ODE Inhibit the Pathways of PI3K/Akt/mTOR in Osteoclastogenesis Induced by RANKL
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Isolation of LGS and ODE from the Twigs of S. oblata subsp. dilatata
4.3. Cell Culture and Osteoclast Differentiation
4.4. Cell Viability Assay
4.5. Tartrate-Resistant Acid Phosphatase (TRAP) Activity and Staining
4.6. Measurement of Actin Ring Formation
4.7. Bone Resorption Assay
4.8. Real-Time Quantitative PCR
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Warriner, A.H.; Patkar, N.M.; Curtis, J.R.; Delzell, E.; Gary, L.; Kilgore, M.; Saag, K. Which fractures are most attributable to osteoporosis? J. Clin. Epidemiol. 2011, 64, 46–53. [Google Scholar] [CrossRef] [Green Version]
- De Villiers, T.J.; Altomare, C.; Particco, M.; Gambacciani, M. Effects of ospemifene on bone in postmenopausal women. Climacteric 2019, 22, 442–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, N.E. Epidemiology, etiology, and diagnosis of osteoporosis. Am. J. Obstet. Gynecol. 2006, 194, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Yu, C.; You, F.; He, C.; Li, Z. Mechanisms of zuogui pill in treating osteoporosis: Perspective from bone marrow mesenchymal stem cells. Evid. Based Complement. Alternat. Med. 2018, 2018, 3717391. [Google Scholar] [CrossRef]
- Shin, C.S.; Cho, H.Y. Bone remodeling and mineralization. J. Korean Soc. Endocrinol. 2005, 20, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Boyle, W.J.; Simonet, W.S.; Lacey, D.L. Osteoclast differentiation and activation. Nature 2003, 423, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Asagiri, M.; Takayanagi, H. The molecular understanding of osteoclast differentiation. Bone 2007, 40, 251–264. [Google Scholar] [CrossRef]
- Deepak, V.; Kasonga, A.; Kruger, M.C.; Coetzee, M. Inhibitory effects of eugenol on RANKL-induced osteoclast formation via attenuation of NF-kappaB and MAPK pathways. Connect. Tissue Res. 2015, 56, 195–203. [Google Scholar] [CrossRef]
- Zhang, S.; Feng, P.; Mo, G.; Li, D.; Li, Y.; Mo, L.; Yang, Z.; Liang, D. Icariin influences adipogenic differentiation of stem cells affected by osteoblast-osteoclast co-culture and clinical research adipogenic. Biomed. Pharmacother. 2017, 88, 436–442. [Google Scholar] [CrossRef]
- Opletal, L.; Sovova, H.; Bartlova, M. Dibenzo[a,c]cyclooctadiene lignans of the genus Schisandra: Importance, isolation and determination. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2004, 812, 357–371. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, J.; Si, S.; Chen, L.; Wang, Z. T63 inhibits osteoclast differentiation through regulating MAPKs and Akt signaling pathways. Eur. J. Pharmacol. 2018, 834, 30–35. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef] [PubMed]
- Tobeiha, M.; Moghadasian, M.H.; Amin, N.; Jafarnejad, S. RANKL/RANK/OPG pathway: A mechanism involved in exercise-induced bone remodeling. Biomed. Res. Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, F.L.; Xu, R.S.; Jiang, D.L.; He, X.L.; Su, Q.; Jin, C.; Li, X. Leonurine hydrochloride inhibits osteoclastogenesis and prevents osteoporosis associated with estrogen deficiency by inhibiting the NF-κB and PI3K/Akt signaling pathways. Bone 2015, 75, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Nakashima, T.; Hiroshi, N.; Penninger, J.M. RANKL-RANK signaling in osteoclastogenesis and bone disease. Trends Mol. Med. 2006, 12, 17–25. [Google Scholar] [CrossRef]
- Choi, J.; Choi, S.Y.; Lee, S.Y.; Lee, J.Y.; Kim, H.S.; Lee, S.Y.; Lee, N.K. Caffeine enhances osteoclast differentiation and maturation through p38 MAP kinase/Mitf and DC-STAMP/CtsK and TRAP pathway. Cell Signal. 2013, 25, 1222–1227. [Google Scholar] [CrossRef]
- Soysa, N.S.; Alles, N.; Shimokawa, H.; Jimi, E.; Aoki, K.; Ohya, K. Inhibition of the classical NF-kappaB pathway prevents osteoclast bone-resorbing activity. J. Bone Miner. Metab. 2009, 27, 131–139. [Google Scholar] [CrossRef]
- Park, K.J.; Subedi, L.; Kim, S.Y.; Lee, K.R. Anti-inflammatory terpenoid derivatives from the twigs of Syringa oblata var. dilatata. Phytochem. Lett. 2018, 27, 183–186. [Google Scholar] [CrossRef]
- Park, K.J.; Suh, W.S.; Subedi, L.; Kim, S.Y.; Choi, S.U.; Lee, K.R. Secoiridoid glucosides from the twigs of Syringa oblata var. dilatata and their neuroprotective and cytotoxic activities. Chem. Pharm. Bull. 2017, 65, 359–364. [Google Scholar]
- Zhao, M.; Tang, W.X.; Li, J.; Bai, L.M.; Wang, J.L.; Zhang, W.Z.; Zhang, S.J. Two new monoterpenoids from the fresh leaves of Syringa oblata. Chem. Nat. Comp. 2016, 52, 1023–1025. [Google Scholar] [CrossRef]
- Oh, H.; Ko, E.K.; Kim, D.H.; Jang, K.K.; Park, S.E.; Lee, H.S.; Kim, Y.C. Secoiridoid glucosides with free radical scavenging activity from the leaves of Syringa dilatata. Phytother. Res. 2003, 17, 417–419. [Google Scholar] [CrossRef]
- Dunlop, E.A.; Tee, A.R. mTOR and autophagy: A dynamic relationship governed by nutrients and energy. Semin. Cell. Dev. Biol. 2014, 36, 121–129. [Google Scholar] [CrossRef]
- Tovar, E.R.; Muriel, P. Phytotherapy for the liver. In Dietary Interventions in Liver Disease; Academic Press: Cambridge, MA, USA, 2019; Chapter 9; pp. 101–121. [Google Scholar]
- Miyazaki, T.; Tokimura, F.; Tanaka, S. A review of denosumab for the treatment of osteoporosis. Patient Prefer. Adherence 2014, 8, 463–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robling, A.G.; Castillo, A.B.; Turner, C.H. Biomechanical and molecular regulation of bone remodeling. Annu. Rev. Biomed. 2006, 8, 455–498. [Google Scholar] [CrossRef]
- Takayanagi, H.; Kim, S.; Koga, T.; Nishina, H.; Isshiki, M.; Yoshida, H.; Saiura, A.; Isobe, M.; Yokochi, T.; Inoue, J.; et al. Induction and activation of the transcription factor NFATc1 (NFAT2) integrate RANKL signaling in terminal differentiation of osteoclasts. Dev. Cell 2002, 3, 889. [Google Scholar] [CrossRef] [Green Version]
- Suda, K.; Woo, J.T.; Takami, M.; Sexton, P.M.; Nagai, K. Lipopolysaccharide supports survival and fusion of preosteoclasts independent of TNF-alpha, IL-1, and RANKL. J. Cell. Physiol. 2002, 190, 101–108. [Google Scholar] [CrossRef]
- Park, B.K.; Zhang, H.; Zeng, Q.; Dai, J.; Keller, E.T.; Giordano, T.; Gu, K.; Shah, V.; Pei, L.; Zarbo, R.J.; et al. NF-kappaB in breast cancer cells promotes osteolytic bone metastasis by inducing osteoclastogenesis via GM-CSF. Nat. Med. 2007, 13, 62–69. [Google Scholar] [CrossRef]
- Li, Y.; Sun, R.; Zou, J.; Ying, Y.; Luo, Z. Dual roles of the AMP-activated protein kinase pathway in angiogenesis. Cells 2019, 8, 752. [Google Scholar] [CrossRef] [Green Version]
- Gan, Z.Y.; Fitter, S.; Vandyke, K.; To, L.B.; Zannettino, A.C.; Martin, S.K. The effect of the dual PI3K and mTOR inhibitor BEZ235 on tumour growth and osteolytic bone disease in multiple myeloma. Eur. J. Haematol. 2015, 94, 343–354. [Google Scholar] [CrossRef]
- Brendan, F.; Boyce, M.D. Advances in osteoclast biology reveal potential new drug targets and new roles for osteoclasts. J. Bone Miner. Res. 2013, 28, 711–722. [Google Scholar]
- Saravanan, S.; Hairul Islam, V.I.; Thirugnanasambantham, K.; Pazhanivel, N.; Raghuraman, N.; Gabriel Paulraj, M.; Ignacimuthu, S. Swertiamarin ameliorates inflammation and osteoclastogenesis intermediates in IL-1β induced rat fibroblast-like synoviocytes. Inflamm. Res. 2014, 63, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.C.; Lin, S.L. Secoiridoid glycosides from Jasminum polyanthum. J. Chin. Chem. Soc. 1996, 43, 171–176. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | Ligustroside (LGS) | Oleoside Dimethyl Ester (ODE) | ||
|---|---|---|---|---|
| δ1H | δ13C | δ1H | δ13C | |
| 1 | 5.94 (1H, s) | 95.3 | 5.95 (1H, s) | 95.3 |
| 2 | - | - | - | - |
| 3 | 7.54 (1H, s) | 155.3 | 7.55 (1H, s) | 155.3 |
| 4 | - | 109.5 | - | 109.5 |
| 5 | 3.98 (1H, dd, J = 9.9, 4.2 Hz) | 32.0 | 4.01 (1H, dd, J = 9.0, 4.2 Hz) | 32.0 |
| 6 | a: 2.72 (1H, dd, J = 14.1, 4.2 Hz) b: 2.44 (1H, dd, J = 14.1, 9.9 Hz) | 41.4 | a: 2.76 (1H, dd, J = 14.1, 4.2 Hz) b: 2.46 (1H, dd, J = 14.1, 9.0 Hz) | 41.2 |
| 7 | - | 173.3 | - | 173.7 |
| 8 | 6.09 (1H, q, J = 6.6 Hz) | 125.0 | 6.13 (1H, q, J = 6.3 Hz) | 125.0 |
| 9 | - | 130.6 | - | 130.6 |
| 10 | 1.67 (3H, d, J = 6.6 Hz) | 13.7 | 1.76 (3H, d, J = 6.3 Hz) | 13.7 |
| 11 | - | 168.8 | - | 168.8 |
| 1′ | a: 4.23 (1H, dt, J = 10.8, 6.9 Hz) b: 4.00 (1H, dd, J = 10.8, 6.9 Hz) | 67.0 | - | - |
| 2′ | 2.83 (2H, t, J = 6.8 Hz) | 35.3 | - | - |
| 3′ | - | 130.2 | - | - |
| 4′ | 7.08 (2H, d, J = 8.4 Hz) | 131.1 | - | - |
| 5′ | 6.75 (2H, d, J = 8.4 Hz) | 116.4 | - | - |
| 6′ | - | 157.2 | - | - |
| 7′ | 6.75 (2H, d, J = 8.4 Hz) | 116.4 | - | - |
| 8′ | 7.08 (2H, d, J = 8.4 Hz) | 131.1 | - | - |
| 1″ | 4.83 (1H, d, J = 7.8 Hz) | 101.0 | 4.83 (1H, d, J = 7.8 Hz) | 101.0 |
| 2″ | 3.33–4.82 (5H, m) | 74.9 | 3.33–4.82 (5H, m) | 74.9 |
| 3″ | 78.6 | - | 78.5 | |
| 4″ | 71.6 | - | 71.6 | |
| 5″ | 78.1 | - | 78.1 | |
| 6″ | 62.9 | - | 62.8 | |
| COOCH3 | 3.69 (3H, s) | 52.0 | 3.71 (3H, s) | 52.0 |
| COOMe | - | - | 3.66 (3H, s) | 52.3 |
| Target Gene | Sequence (5′-3′) | |
|---|---|---|
| dcstamp | Forward | TTTGCCGCTGTGGACTATCTGC |
| Reverse | GCAGAATCATGGACGACTCCTTG | |
| acp5 | Forward | CGTCTCTGCACAGATTGCAT |
| Reverse | GAGTTGCCACACAGCATCAC | |
| Atp6v0d2 | Forward | TGTGTCCCATTCTTGAGTTTGAGG |
| Reverse | AGG GTCTCCCTGTCTTCTTTGCTT | |
| gapdh | Forward | ACAGTCCATGCCATCACTGCC |
| Reverse | GCCTGCTTCACCACCTTCTTG | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.-R.; Kim, E.-N.; Park, K.J.; Kim, K.H.; Jeong, G.-S. Inhibitory Effect of LGS and ODE Isolated from the Twigs of Syringa oblata subsp. dilatata on RANKL-Induced Osteoclastogenesis in Macrophage Cells. Molecules 2021, 26, 1779. https://doi.org/10.3390/molecules26061779
Kim G-R, Kim E-N, Park KJ, Kim KH, Jeong G-S. Inhibitory Effect of LGS and ODE Isolated from the Twigs of Syringa oblata subsp. dilatata on RANKL-Induced Osteoclastogenesis in Macrophage Cells. Molecules. 2021; 26(6):1779. https://doi.org/10.3390/molecules26061779
Chicago/Turabian StyleKim, Ga-Ram, Eun-Nam Kim, Kyoung Jin Park, Ki Hyun Kim, and Gil-Saeng Jeong. 2021. "Inhibitory Effect of LGS and ODE Isolated from the Twigs of Syringa oblata subsp. dilatata on RANKL-Induced Osteoclastogenesis in Macrophage Cells" Molecules 26, no. 6: 1779. https://doi.org/10.3390/molecules26061779