
Two Key Amino Acids Variant of α-l-arabinofuranosidase from Bacillus subtilis Str. 168 with Altered Activity for Selective Conversion Ginsenoside Rc to Rd

Abstract
:1. Introduction
2. Results and Discussion
2.1. Cloning of BsAbfA Gene and Sequence Analysis
2.2. Identification of Enzymatic Properties of Recombinant BsAbfA
2.3. Temperature and pH Dependence of Recombinant BsAbfA
2.4. Kinetic Analysis of BsAbfA
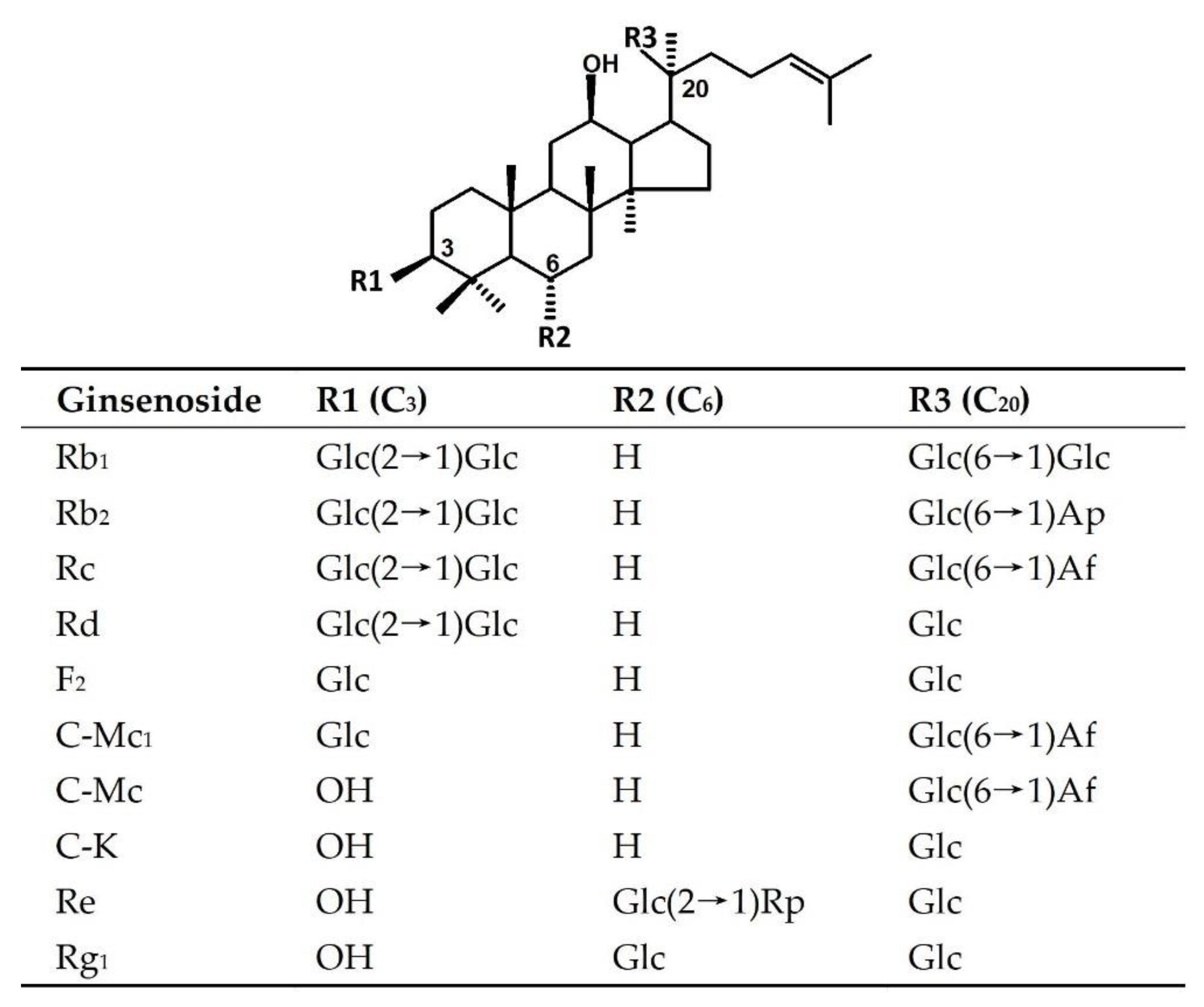
2.5. Substrate Specificity of BsAbfA
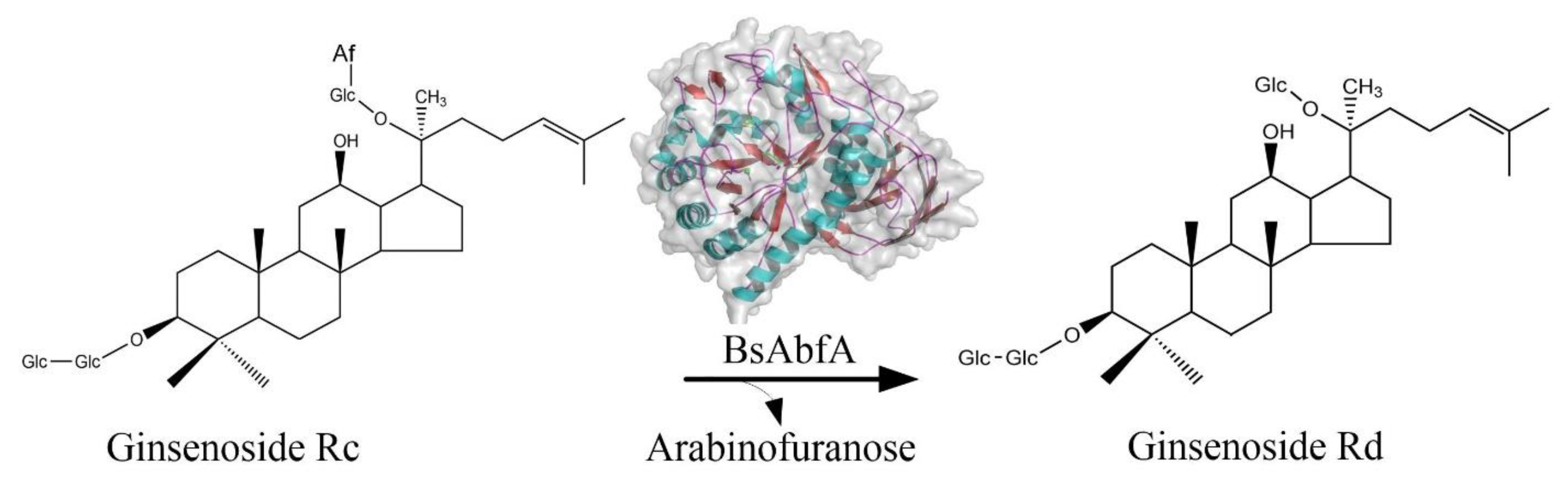
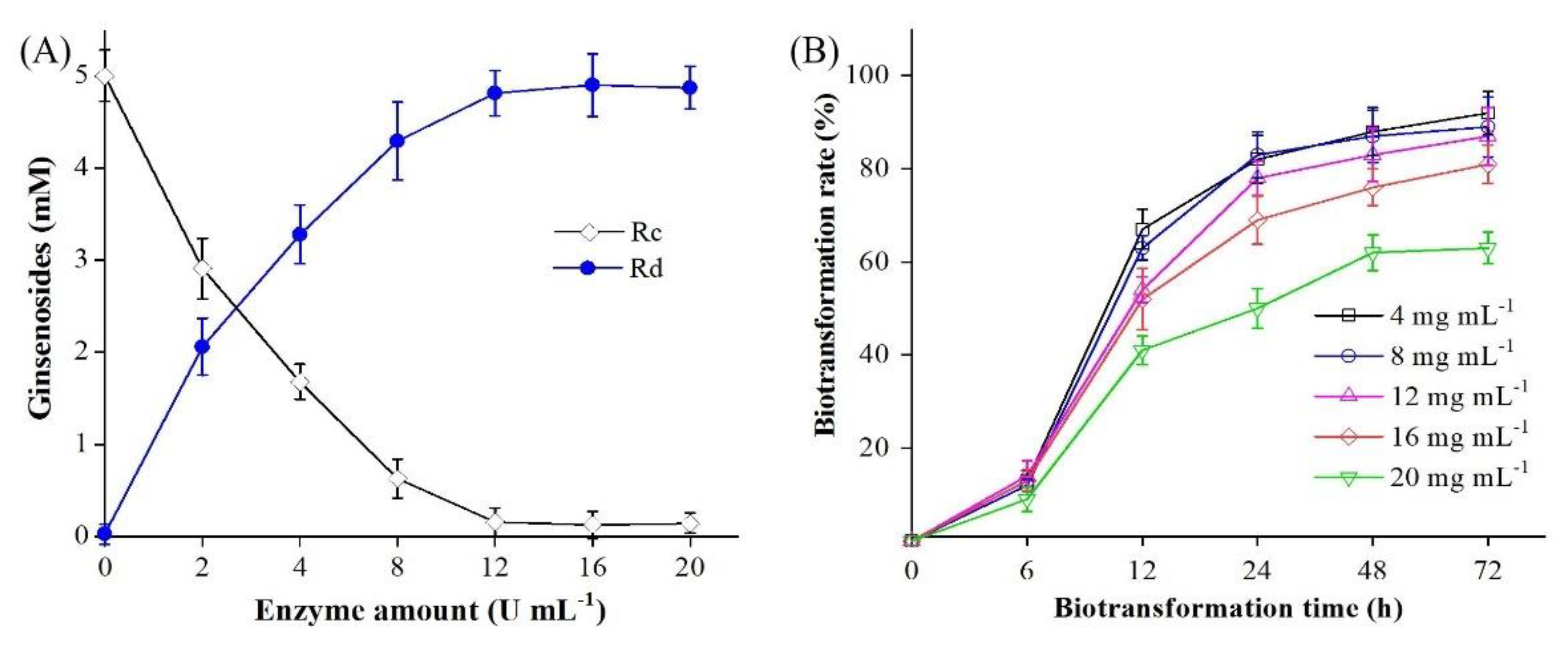
2.6. Biotransformation of Ginsenoside Rc by BsAbfA
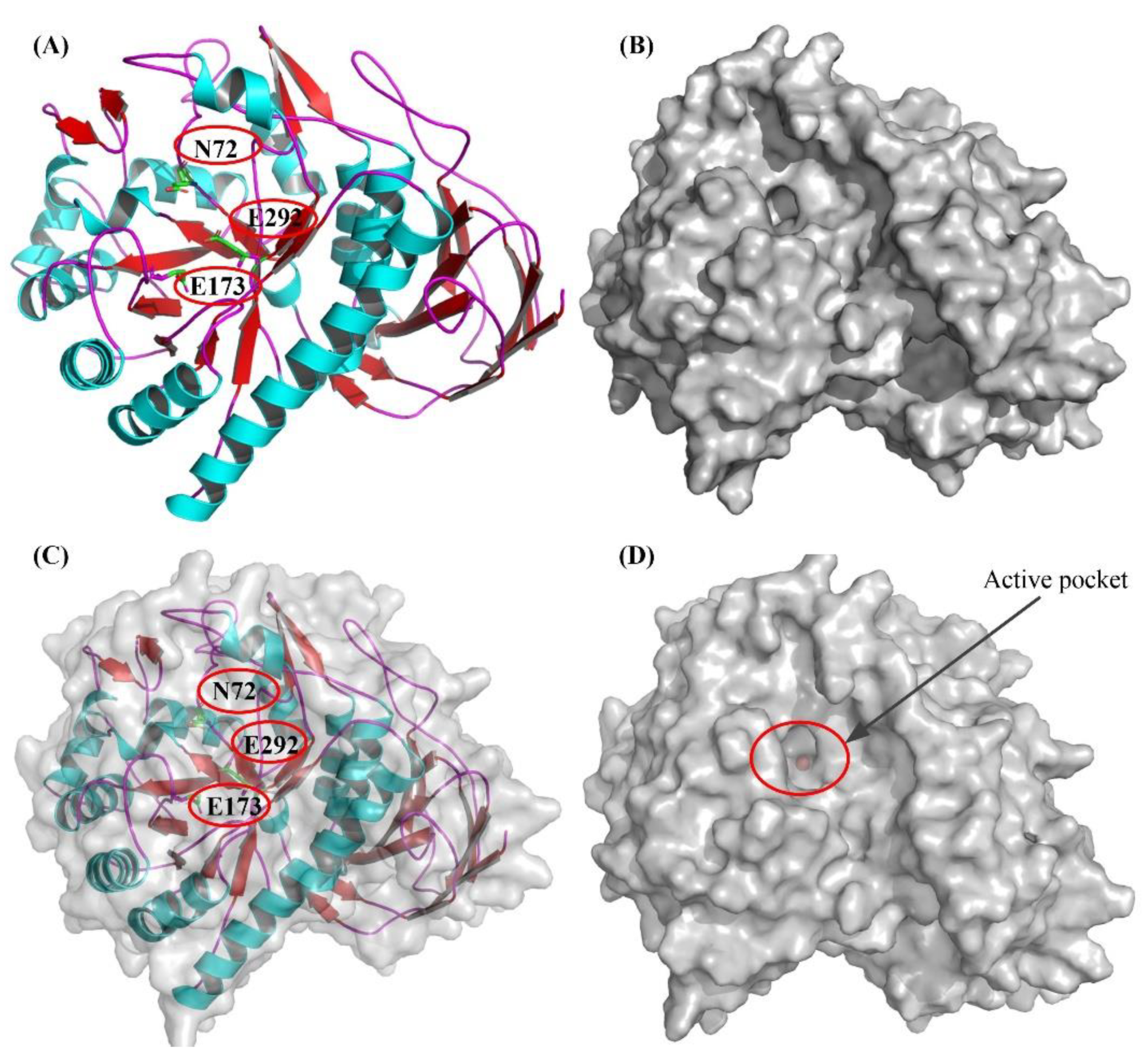
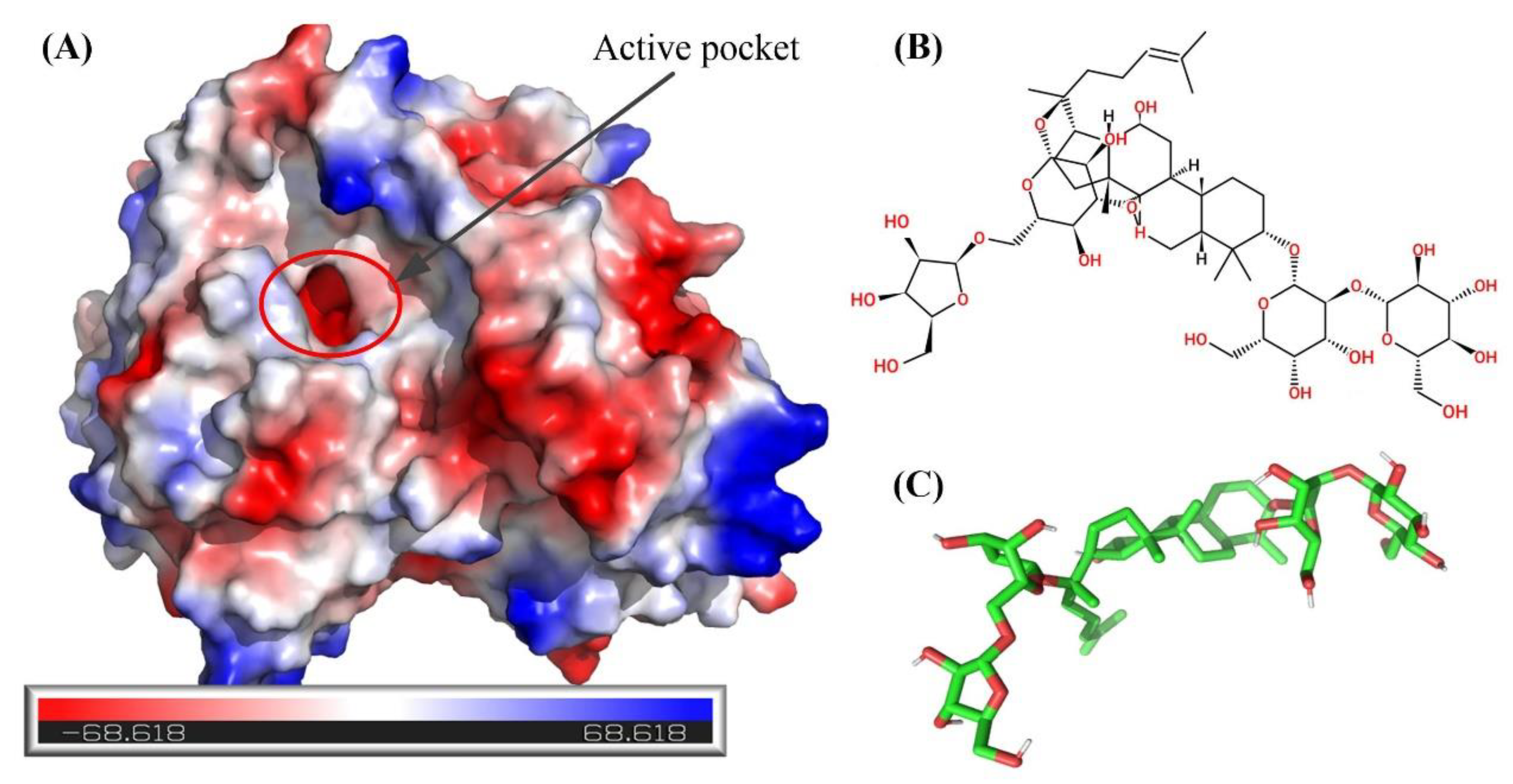
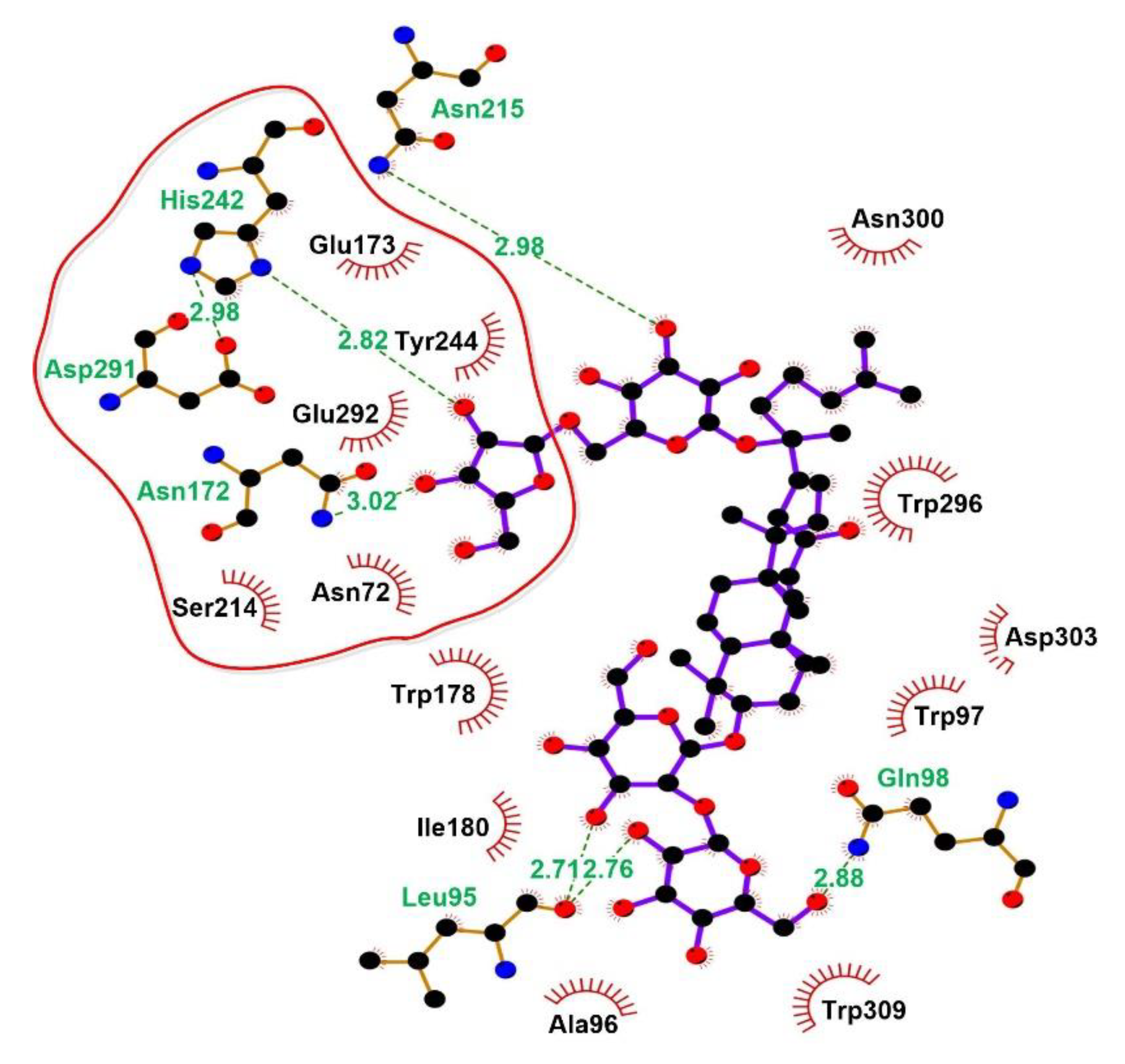
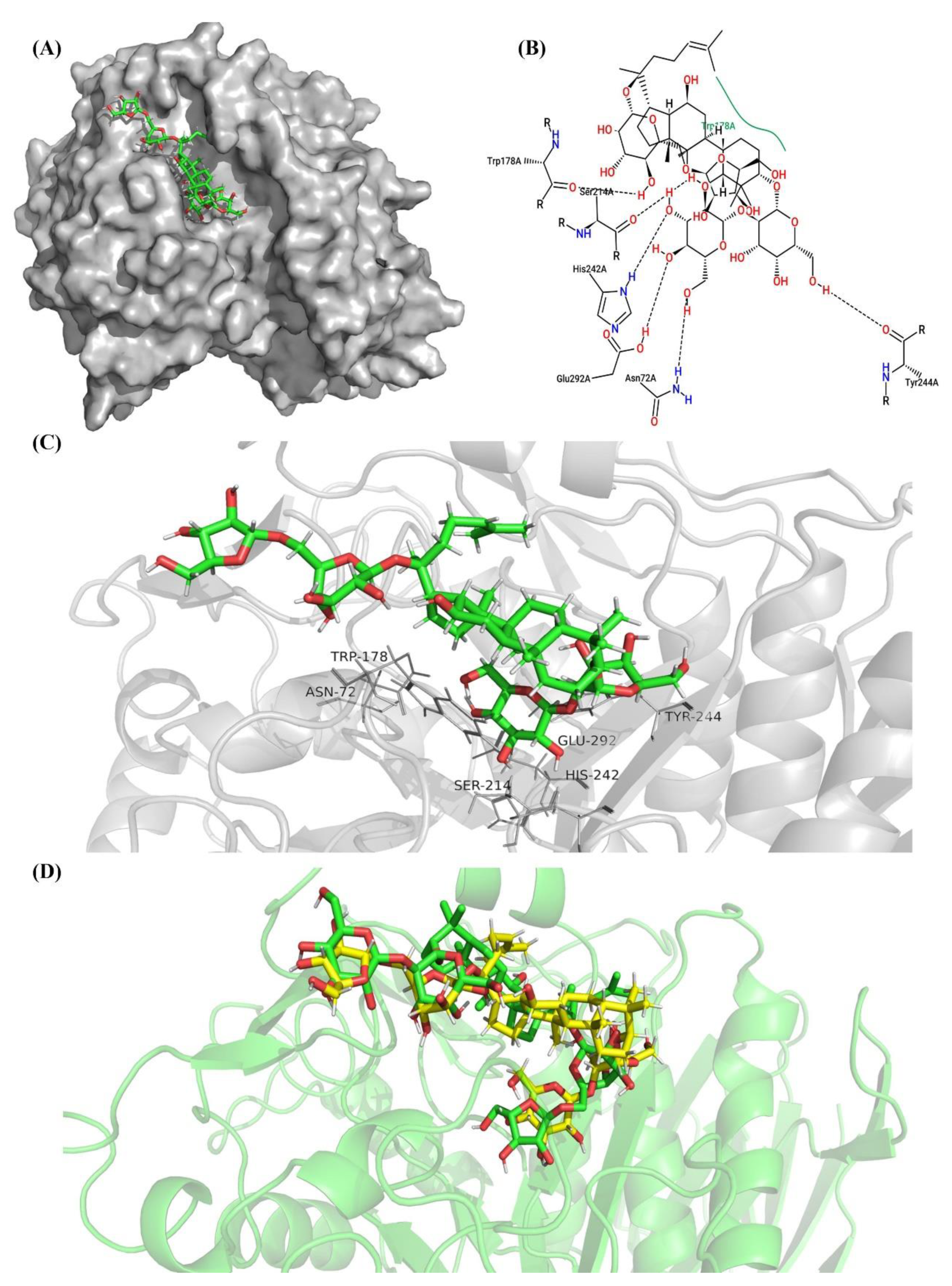
2.7. Molecular Docking and Examination of BsAbfA Active Site
3. Materials and Methods
3.1. Bacterial Strains, Plasmids, and Chemicals
3.2. Cloning, Site-directed Mutagenesis, Heterologous Expression, and Protein Purification
3.3. Determination of Kinetic Parameters and Substrate Specificity
3.4. Effects of pH, Temperature, Metal Ions, and Chemicals on Stability
3.5. Biotransformation of Ginsenoside Rc
3.6. Homology Modeling and Molecular Docking
3.6.1. Homology Modeling
3.6.2. Molecular Docking
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Kim, J.H.; Oh, J.M.; Chun, S.; Park, H.Y.; Im, W.T. Enzymatic biotransformation of ginsenoside Rb2 into Rd by recombinant alpha-l-arabinopyranosidase from Blastococcus saxobsidens. J. Microbiol. Biotechnol. 2020, 30, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Zhang, Z.; Su, H.; Xu, P.; Qi, H.; Zhao, D.; Li, X. Panax ginseng clinical trials: Current status and future perspectives. Biomed. Pharmacother. 2020, 132, 110832. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Choi, E.; Shin, K.K.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Hossain, M.A.; Kim, H.S.; Yi, Y.S.; Kim, D.; et al. Compound K, a ginsenoside metabolite, plays an antiinflammatory role in macrophages by targeting the AKT1-mediated signaling pathway. J. Ginseng Res. 2019, 43, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Xue, Q.; He, N.; Wang, Z.; Fu, X.; Aung, L.H.H.; Liu, Y.; Li, M.; Cho, J.Y.; Yang, Y.; Yu, T. Functional roles and mechanisms of ginsenosides from Panax ginseng in atherosclerosis. J. Ginseng Res. 2021, 45, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Shin, B.K.; Kwon, S.W.; Park, J.H. Chemical diversity of ginseng saponins from Panax ginseng. J. Ginseng Res. 2015, 39, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Balan, P.; Popovich, D.G. Ginsenosides analysis of New Zealand-grown forest Panax ginseng by LC-QTOF-MS/MS. J. Ginseng Res. 2020, 44, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.S.; Gu, L.J.; Fang, Z.M.; Wang, C.Y.; Wang, Z.; Lee, M.R.; Li, Z.; Li, J.J.; Sung, C.K. Simultaneous quantification of 19 ginsenosides in black ginseng developed from Panax ginseng by HPLC-ELSD. J. Pharm. Biomed. Anal. 2009, 50, 15–22. [Google Scholar] [CrossRef]
- Zhang, F.; Tang, S.; Zhao, L.; Yang, X.; Yao, Y.; Hou, Z.; Xue, P. Stem-leaves of Panax as a rich and sustainable source of less-polar ginsenosides: Comparison of ginsenosides from Panax ginseng, American ginseng and Panax notoginseng prepared by heating and acid treatment. J. Ginseng Res. 2021, 45, 163–175. [Google Scholar] [CrossRef]
- Liu, G.M.; Lu, T.C.; Sun, M.L.; Jia, W.Y.; Ji, X.; Luo, Y.G. Ginsenoside Rd inhibits glioblastoma cell proliferation by up-regulating the expression of miR-144-5p. Biol. Pharm. Bull. 2020, 43, 1534–1541. [Google Scholar] [CrossRef]
- Wang, P.; Du, X.; Xiong, M.; Cui, J.; Yang, Q.; Wang, W.; Chen, Y.; Zhang, T. Ginsenoside Rd attenuates breast cancer metastasis implicating derepressing microRNA-18a-regulated Smad2 expression. Sci. Rep. 2016, 6, 33709. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Liang, G.; Lin, J.; Zhang, S.; Lin, Q.; Ji, X.; Chen, H.; Li, N.; Jin, S. Ginsenoside Rd therapy improves histological and functional recovery in a rat model of inflammatory bowel disease. Phytother. Res. 2020, 34, 3019–3028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, X.; Hu, G.; Zhang, G.; Zhao, G.; Shi, M. Ginsenoside Rd attenuates blood-brain barrier damage by suppressing proteasome-mediated signaling after transient forebrain ischemia. Neuroreport 2020, 31, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Zhang, X.; Kong, X.; Han, J.; Yang, Q.; Zhang, Y.; Chen, Y.; Li, P.; Liu, J.; Shi, M.; et al. Ginsenoside Rd attenuates mitochondrial dysfunction and sequential apoptosis after transient focal ischemia. Neuroscience 2011, 178, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Anufriev, V.P.; Malinovskaya, G.V.; Denisenko, V.A.; Uvarova, N.I.; Elyakov, G.B.; Kim, S.I.; Baek, N.I. Synthesis of ginsenoside Rg3, a minor constituent of Ginseng radix. Carbohydr. Res. 1997, 304, 179–182. [Google Scholar] [CrossRef]
- An, D.S.; Cui, C.H.; Sung, B.H.; Yang, H.C.; Kim, S.C.; Lee, S.T.; Im, W.T.; Kim, S.G. Characterization of a novel ginsenoside-hydrolyzing alpha-l-arabinofuranosidase, AbfA, from Rhodanobacter ginsenosidimutans Gsoil 3054T. Appl. Microbiol. Biotechnol. 2012, 94, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.M.; Jung, H.M.; Cui, C.H.; Sung, B.H.; Kim, J.K.; Kim, S.G.; Lee, S.T.; Kim, S.C.; Im, W.T. Bioconversion of ginsenoside Rc into Rd by a novel alpha-l-arabinofuranosidase, Abf22-3 from Leuconostoc sp. 22-3: Cloning, expression, and enzyme characterization. Antonie Leeuwenhoek 2013, 103, 747–754. [Google Scholar]
- Renchinkhand, G.; Cho, S.H.; Park, Y.W.; Song, G.Y.; Nam, M.S. Biotransformation of major ginsenoside Rb1 to Rd by Dekkera anomala YAE-1 from Mongolian fermented milk (Airag). J. Microbiol. Biotechnol. 2020, 30, 1536–1542. [Google Scholar] [CrossRef]
- Zhong, F.L.; Ma, R.; Jiang, M.; Dong, W.W.; Jiang, J.; Wu, S.; Li, D.; Quan, L.H. Cloning and characterization of ginsenoside-hydrolyzing beta-glucosidase from Lactobacillus brevis that transforms ginsenosides Rb1 and F2 into ginsenoside Rd and compound K. J. Microbiol. Biotechnol. 2016, 26, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Zhang, B.; Song, W.X.; Wang, A.; Ni, M.; Luo, X.; Aung, H.H.; Xie, J.T.; Tong, R.; He, T.C.; et al. Steamed American ginseng berry: Ginsenoside analyses and anticancer activities. J. Agric. Food Chem. 2006, 54, 9936–9942. [Google Scholar] [CrossRef]
- Kim, T.H.; Yang, E.J.; Shin, K.C.; Hwang, K.H.; Park, J.S.; Oh, D.K. Enhanced production of β-D-glycosidase and α-l-arabinofuranosidase in recombinant Escherichia coli in fed-batch culture for the biotransformation of ginseng leaf extract to ginsenoside compound K. Biotechnol. Bioproc. E 2018, 23, 183–193. [Google Scholar] [CrossRef]
- Taylor, E.J.; Smith, N.L.; Turkenburg, J.P.; D’Souza, S.; Gilbert, H.J.; Davies, G.J. Structural insight into the ligand specificity of a thermostable family 51 arabinofuranosidase, Araf51, from Clostridium thermocellum. Biochem. J. 2006, 395, 31–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debeche, T.; Bliard, C.; Debeire, P.; O’Donohue, M.J. Probing the catalytically essential residues of the alpha-l-arabinofuranosidase from Thermobacillus xylanilyticus. Protein Eng. 2002, 15, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.Y.; Park, S.Y.; Sung, J.H.; Kim, D.H. Purification and characterization of alpha-l-arabinopyranosidase and alpha-l-arabinofuranosidase from Bifidobacterium breve K-110, a human intestinal anaerobic bacterium metabolizing ginsenoside Rb2 and Rc. Appl. Environ. Microbiol. 2003, 69, 7116–7123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, K.C.; Lee, G.W.; Oh, D.K. Production of ginsenoside Rd from ginsenoside Rc by alpha-l-arabinofuranosidase from Caldicellulosiruptor saccharolyticus. J. Microbiol. Biotechnol. 2013, 23, 483–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.H.; Shin, K.C.; Oh, D.K. An L213A variant of beta-glycosidase from Sulfolobus solfataricus with increased alpha-l-arabinofuranosidase activity converts ginsenoside Rc to compound K. PLoS ONE 2018, 13, e0191018. [Google Scholar]
- Xie, J.; Zhao, D.; Zhao, L.; Pei, J.; Xiao, W.; Ding, G.; Wang, Z.; Xu, J. Characterization of a novel arabinose-tolerant alpha-l-arabinofuranosidase with high ginsenoside Rc to ginsenoside Rd bioconversion productivity. J. Appl. Microbiol. 2016, 120, 647–660. [Google Scholar] [CrossRef]
- Canakci, S.; Belduz, A.O.; Saha, B.C.; Yasar, A.; Ayaz, F.A.; Yayli, N. Purification and characterization of a highly thermostable alpha-l-arabinofuranosidase from Geobacillus caldoxylolyticus TK4. Appl. Microbiol. Biotechnol. 2007, 75, 813–820. [Google Scholar] [CrossRef]
- Pitson, S.M.; Voragen, A.G.; Beldman, G. Stereochemical course of hydrolysis catalyzed by arabinofuranosyl hydrolases. FEBS Lett. 1996, 398, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Pazos, F.; Heredia, P.; Valencia, A.; de las Rivas, J. Threading structural model of the manganese-stabilizing protein PsbO reveals presence of two possible beta-sandwich domains. Protein Struct. Funct. Genet. 2001, 45, 372–381. [Google Scholar] [CrossRef]
- Shallom, D.; Belakhov, V.; Solomon, D.; Shoham, G.; Baasov, T.; Shoham, Y. Detailed kinetic analysis and identification of the nucleophile in alpha-l-arabinofuranosidase from Geobacillus stearothermophilus T-6, a family 51 glycoside hydrolase. J. Biol. Chem. 2002, 277, 43667–43673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shallom, D.; Belakhov, V.; Solomon, D.; Gilead-Gropper, S.; Baasov, T.; Shoham, G.; Shoham, Y. The identification of the acid-base catalyst of alpha-arabinofuranosidase from Geobacillus stearothermophilus T-6, a family 51 glycoside hydrolase. FEBS Lett. 2002, 514, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.R.; Yoon, R.Y.; Seo, E.S.; Kim, Y.S.; Park, C.S.; Oh, D.K. Hydrolytic properties of a thermostable alpha-l-arabinofuranosidase from Caldicellulosiruptor saccharolyticus. J. Appl. Microbiol. 2010, 109, 1188–1197. [Google Scholar] [CrossRef]
- Zuo, S.S.; Wang, Y.C.; Zhu, L.; Zhao, J.Y.; Li, M.G.; Han, X.L.; Wen, M.L. Cloning and characterization of a ginsenoside-hydrolyzing alpha-l-arabinofuranosidase, CaAraf51, from Cellulosimicrobium aquatile Lyp51. Curr. Microbiol. 2020, 77, 2783–2791. [Google Scholar] [CrossRef]
- Zverlov, V.V.; Liebl, W.; Bachleitner, M.; Schwarz, W.H. Nucleotide sequence of arfB of Clostridium stercorarium, and prediction of catalytic residues of alpha-l-arabinofuranosidases based on local similarity with several families of glycosyl hydrolases. FEMS Microbiol. Lett. 1998, 164, 337–343. [Google Scholar]
- Quan, L.H.; Piao, J.Y.; Min, J.W.; Kim, H.B.; Kim, S.R.; Yang, D.U.; Yang, D.C. Biotransformation of ginsenoside Rb1 to prosapogenins, gypenoside XVII, ginsenoside Rd, ginsenoside F2, and compound K by Leuconostoc mesenteroides DC102. J. Ginseng Res. 2011, 35, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Zhong, F.L.; Dong, W.W.; Wu, S.; Jiang, J.; Yang, D.C.; Li, D.; Quan, L.H. Biotransformation of gypenoside XVII to compound K by a recombinant beta-glucosidase. Biotechnol. Lett. 2016, 38, 1187–1193. [Google Scholar] [CrossRef]
- Shin, H.Y.; Lee, J.H.; Lee, J.Y.; Han, Y.O.; Han, M.J.; Kim, D.H. Purification and characterization of ginsenoside Ra-hydrolyzing beta-d-xylosidase from Bifidobacterium breve K-110, a human intestinal anaerobic bacterium. Biol. Pharm. Bull. 2003, 26, 1170–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zechel, D.L.; Withers, S.G. Glycosidase mechanisms: Anatomy of a finely tuned catalyst. Acc. Chem. Res. 2000, 33, 11–18. [Google Scholar] [PubMed]
- Moracci, M.; Capalbo, L.; Ciaramella, M.; Rossi, M. Identification of two glutamic acid residues essential for catalysis in the beta-glycosidase from the thermoacidophilic archaeon Sulfolobus solfataricus. Protein Eng. 1996, 9, 1191–1195. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Trimbur, D.; Graham, R.; Warren, R.A.; Withers, S.G. Identification of the acid/base catalyst in Agrobacterium faecalis beta-glucosidase by kinetic analysis of mutants. Biochemistry (U. S.) 1995, 34, 14554–14562. [Google Scholar] [CrossRef] [PubMed]
- Moracci, M.; Trincone, A.; Perugino, G.; Ciaramella, M.; Rossi, M. Restoration of the activity of active-site mutants of the hyperthermophilic beta-glycosidase from Sulfolobus solfataricus: Dependence of the mechanism on the action of external nucleophiles. Biochemistry (U. S.) 1998, 37, 17262–17270. [Google Scholar] [CrossRef]
- Sun, J.; Wang, W.; Yao, C.; Dai, F.; Zhu, X.; Liu, J.; Hao, J. Overexpression and characterization of a novel cold-adapted and salt-tolerant GH1 beta-glucosidase from the marine bacterium Alteromonas sp. L82. J. Microbiol. 2018, 56, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Noh, K.H.; Oh, D.K. Production of the rare ginsenosides compound K, compound Y, and compound Mc by a thermostable beta-glycosidase from Sulfolobus acidocaldarius. Biol. Pharm. Bull. 2009, 32, 1830–1835. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Nuruzzaman, M.; Xiu, H.; Huang, J.; Wu, K.; Chen, X.; Li, J.; Wang, L.; Jeong, J.H.; Park, S.J.; et al. Transcriptome analysis of methyl jasmonate-elicited Panax ginseng adventitious roots to discover putative ginsenoside biosynthesis and transport genes. Int. J. Mol. Sci. 2015, 16, 3035–3057. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, J.; Yoon, M.S.; Kim, M.J.; Ryu, N.S.; Song, Y.E.; Kim, Y.H.; Kim, M.K. Purification and characterization of a novel ginsenoside Rc-hydrolyzing beta-glucosidase from Armillaria mellea mycelia. AMB Express. 2016, 6, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Quintana, L.; Beltran, D.; Mendez-Yanez, A.; Valenzuela-Riffo, F.; Herrera, R.; Moya-Leon, M.A. Characterization of FcXTH2, a novel xyloglucan endotransglycosylase/hydrolase enzyme of Chilean strawberry with hydrolase activity. Int. J. Mol. Sci. 2020, 21, 3380. [Google Scholar] [CrossRef]
- Zhang, R.; Huang, X.M.; Yan, H.J.; Liu, X.Y.; Zhou, Q.; Luo, Z.Y.; Tan, X.N.; Zhang, B.L. Highly selective production of compound K from ginsenoside Rd by hydrolyzing glucose at C-3 glycoside using beta-glucosidase of Bifidobacterium breve ATCC 15700. J. Microbiol. Biotechn. 2019, 29, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Ji, Q.C.; Harkey, M.R.; Henderson, G.L.; Gershwin, M.E.; Stern, J.S.; Hackman, R.M. Quantitative determination of ginsenosides by high-performance liquid chromatography-tandem mass spectrometry. Phytochem. Anal. 2001, 12, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Wang, S.; Yao, L.; Wang, J.; Gao, W. Quality evaluation of Panax ginseng adventitious roots based on ginsenoside constituents, functional genes, and ferric-reducing antioxidant power. J. Food Biochem. 2019, 43, e12901. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository-new features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30 (Suppl. S1), S162–S173. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019, 47, D1102–D1109. [Google Scholar] [CrossRef] [Green Version]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Substrates a | Relative Activity (%) b |
|---|---|---|
| BsAbfA | pNP-α-Af | 100.1 ± 6.2 |
| BsAbfA | Ginsenoside Rc | 121.6 ± 4.3 |
| E173A | Ginsenoside Rc | ND c |
| E173D | Ginsenoside Rc | 32.3 ± 2.9 |
| E173Q | Ginsenoside Rc | 65.6 ± 2.7 |
| E292A | Ginsenoside Rc | ND |
| E292D | Ginsenoside Rc | 28.7 ± 3.8 |
| E292Q | Ginsenoside Rc | 53.4 ± 2.2 |
| Enzymes | Km (mM) | Kcat (s−1) | Kcat/Km (s−1 mM−1) |
|---|---|---|---|
| BsAbfA | 0.6 ± 0.08 | 108.9 ± 8.6 | 181.5 ± 6.9 |
| E173A | ND b | ND | ND |
| E173D | 0.2 ± 0.05 | 0.5 ± 3.1 | 2.5 ± 0.06 |
| E173Q | 0.4 ± 0.07 | 2.5 ± 2.1 | 6.3 ± 0.09 |
| E292A | ND | ND | ND |
| E292D | 0.7 ± 0.06 | 0.9 ± 0.05 | 1.3 ± 0.03 |
| E292Q | 0.7 ± 0.09 | 1.6 ± 0.04 | 2.3 ± 0.05 |
| Metal Ions or Chemicals | Relative Activity ± SD (%) a | |
|---|---|---|
| 1 mM | 5 mM | |
| Na+ | 100.4 ± 2.1 | 98.3 ± 1.7 |
| K+ | 99.8 ± 1.4 | 93.6 ± 1.5 |
| Ca2+ | 98.4 ± 2.7 | 93.1 ± 1.9 |
| Mg2+ | 99.6 ± 1.8 | 91.5 ± 2.3 |
| Fe2+ | 105.7 ± 2.2 | 95.7 ± 3.1 |
| Mn2+ | 119.7 ± 2.4 | 109.4 ± 2.5 |
| Zn2+ | 87.1 ± 1.9 | 77.8 ± 2.4 |
| Ni2+ | 98.8 ± 2.8 | 91.8 ± 2.5 |
| Cu2+ | 36.8 ± 0.9 | 24.3 ± 1.1 |
| Hg2+ | 20.3 ± 1.7 | 5.3 ± 1.1 |
| EDTA | 100.2 ± 2.3 | 99.7 ± 3.1 |
| DTT | 98.6 ± 2.7 | 97.4 ± 2.8 |
| SDS | 98.2 ± 2.5 | 81.2 ± 3.2 |
| Control | 100 ± 1.9 | 100 ± 2.6 |
| Substrates a | Relative Activity (%) b |
|---|---|
| pNP-α-Af | 100 ± 3.9 |
| pNP-α-Ap | ND c |
| pNP-α-Rp | ND |
| pNP-β-Glc | ND |
| Ginsenoside Rb1 | ND |
| Ginsenoside Rb2 | ND |
| Ginsenoside Rc | 120.6 ± 2.9 |
| Ginsenoside Rd | ND |
| Ginsenoside Re | ND |
| Ginsenoside Rg1 | ND |
| Ginsenoside F2 | ND |
| C-K | ND |
| C-Mc1 | 106.2 ± 3.8 |
| C-Mc | 108.9 ± 3.5 |
| Gentiobiose | ND |
| Sophorose | ND |
| Cluster | Members | Energy | Dissociation Constant (pM) |
|---|---|---|---|
| 1 | 3 | −9.824 | 6.29 × 104 |
| 2 | 2 | −9.158 | 2.19 × 105 |
| 3 | 2 | −9.086 | 2.49 × 105 |
| 4 | 2 | −8.991 | 2.64 × 105 |
| 5 | 1 | −8.927 | 2.86 × 105 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Tan, S.Q.; Zhang, B.L.; Guo, Z.Y.; Tian, L.Y.; Weng, P.; Luo, Z.Y. Two Key Amino Acids Variant of α-l-arabinofuranosidase from Bacillus subtilis Str. 168 with Altered Activity for Selective Conversion Ginsenoside Rc to Rd. Molecules 2021, 26, 1733. https://doi.org/10.3390/molecules26061733
Zhang R, Tan SQ, Zhang BL, Guo ZY, Tian LY, Weng P, Luo ZY. Two Key Amino Acids Variant of α-l-arabinofuranosidase from Bacillus subtilis Str. 168 with Altered Activity for Selective Conversion Ginsenoside Rc to Rd. Molecules. 2021; 26(6):1733. https://doi.org/10.3390/molecules26061733
Chicago/Turabian StyleZhang, Ru, Shi Quan Tan, Bian Ling Zhang, Zi Yu Guo, Liang Yu Tian, Pei Weng, and Zhi Yong Luo. 2021. "Two Key Amino Acids Variant of α-l-arabinofuranosidase from Bacillus subtilis Str. 168 with Altered Activity for Selective Conversion Ginsenoside Rc to Rd" Molecules 26, no. 6: 1733. https://doi.org/10.3390/molecules26061733