
Antioxidant and Anti-Inflammatory Effects of White Mulberry (Morus alba L.) Fruits on Lipopolysaccharide-Stimulated RAW 264.7 Macrophages




Abstract
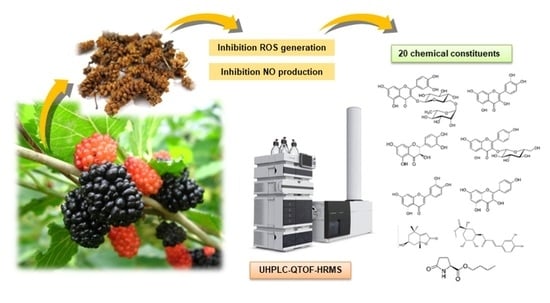
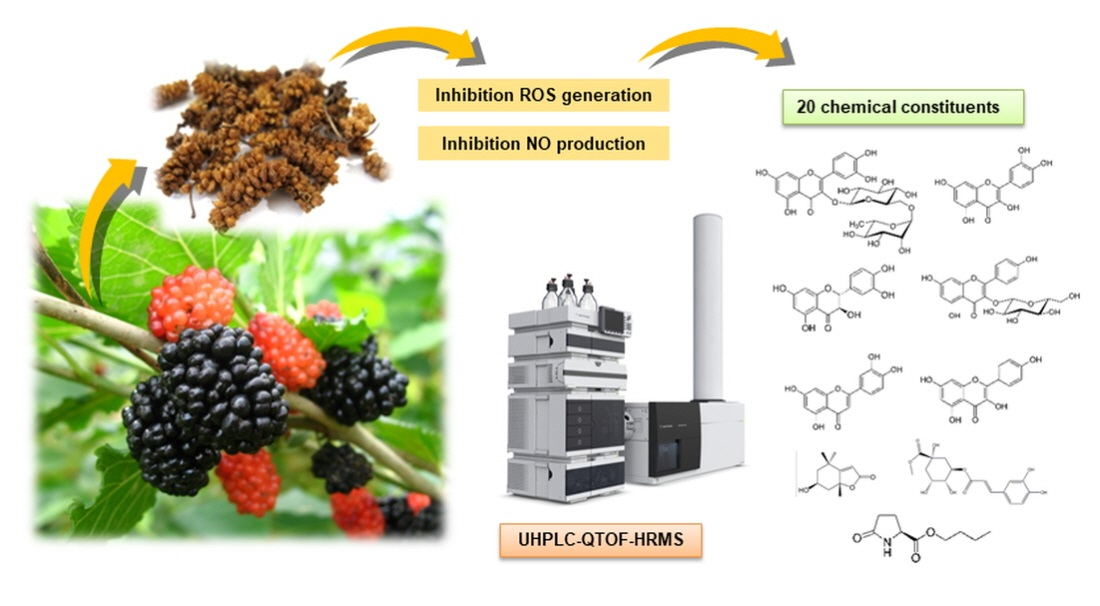
:
1. Introduction
2. Results
2.1. Total Phenolic and Flavonoid Contents
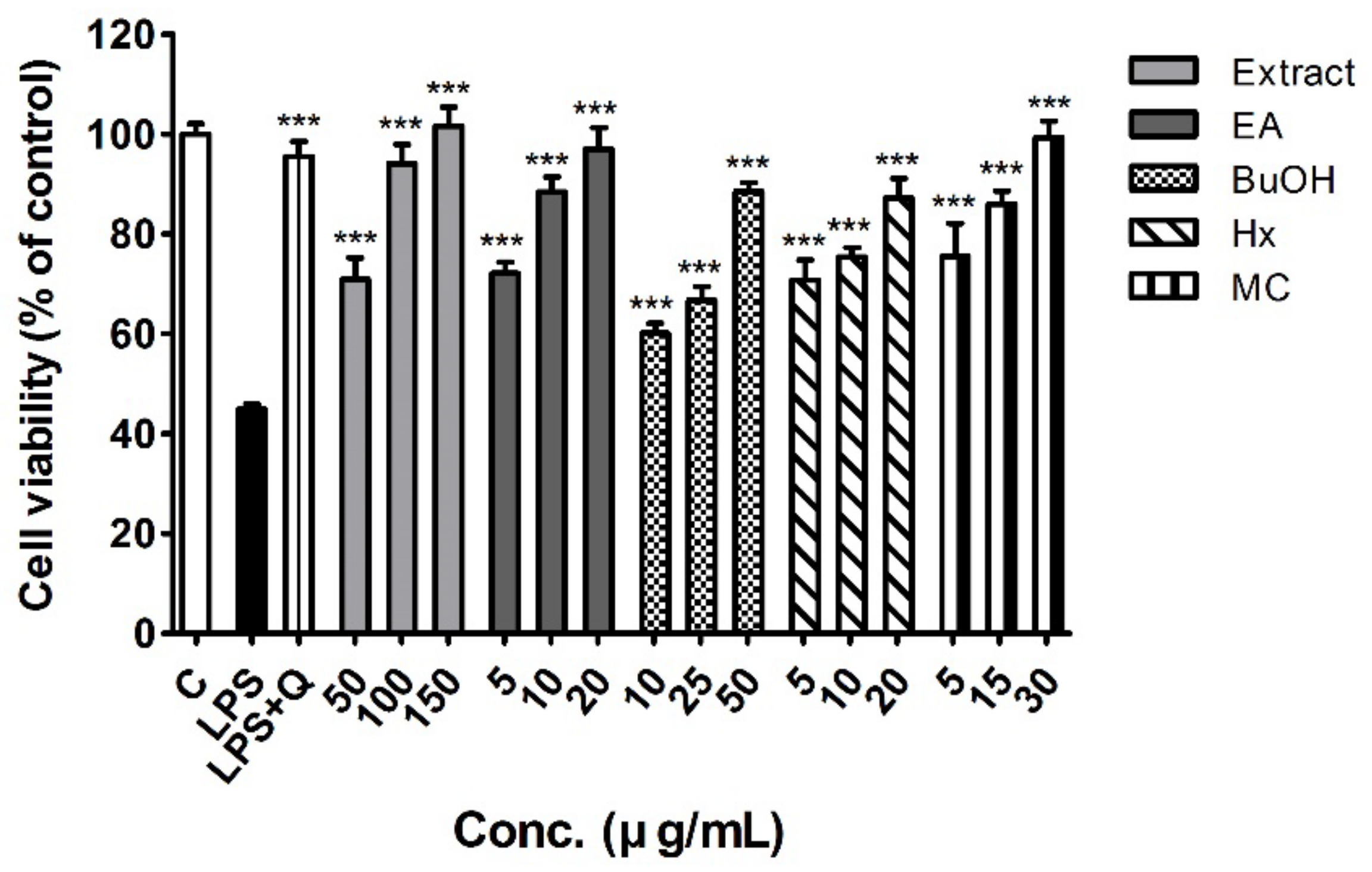
2.2. Cell Viability
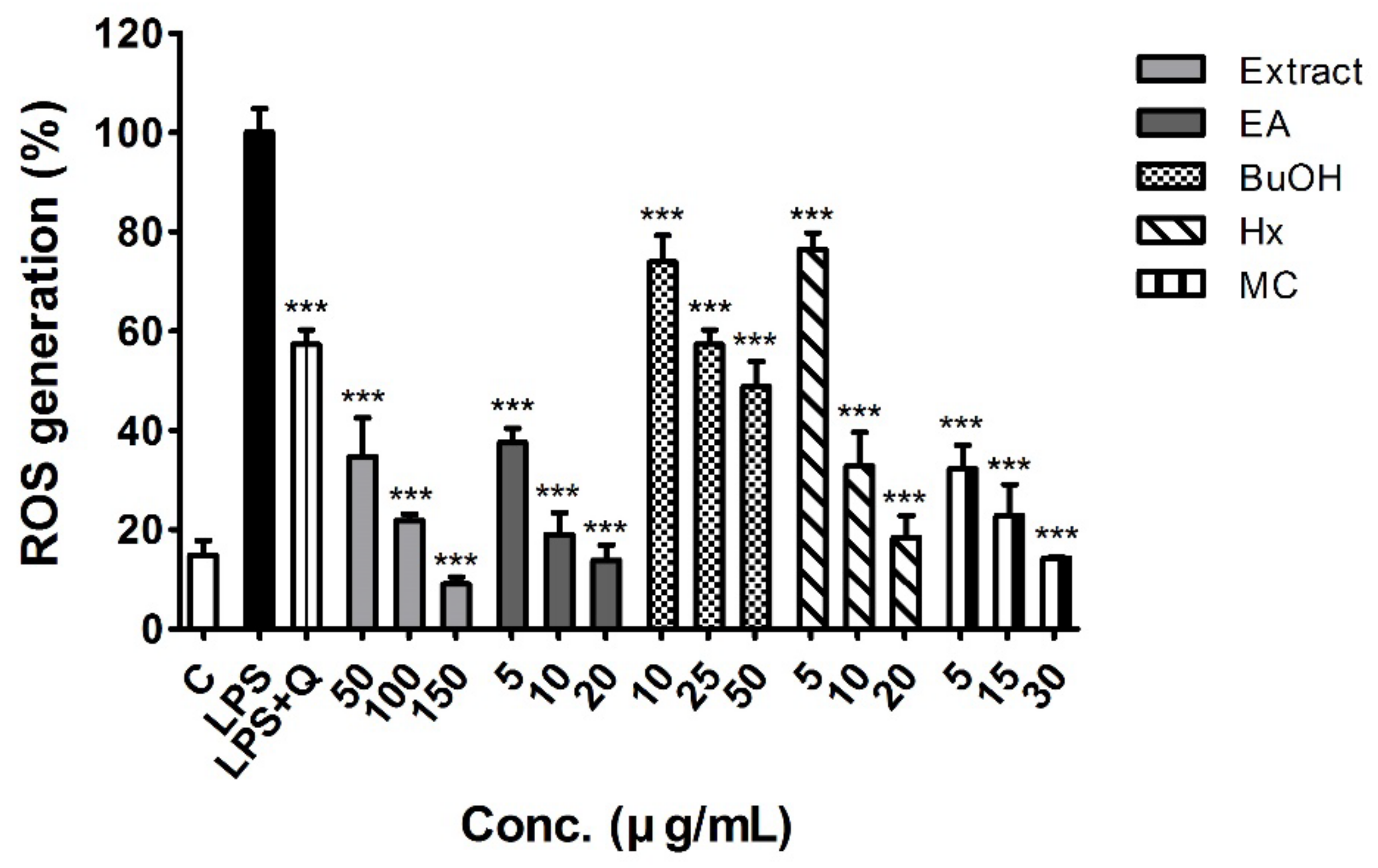
2.3. Antioxidant Activity
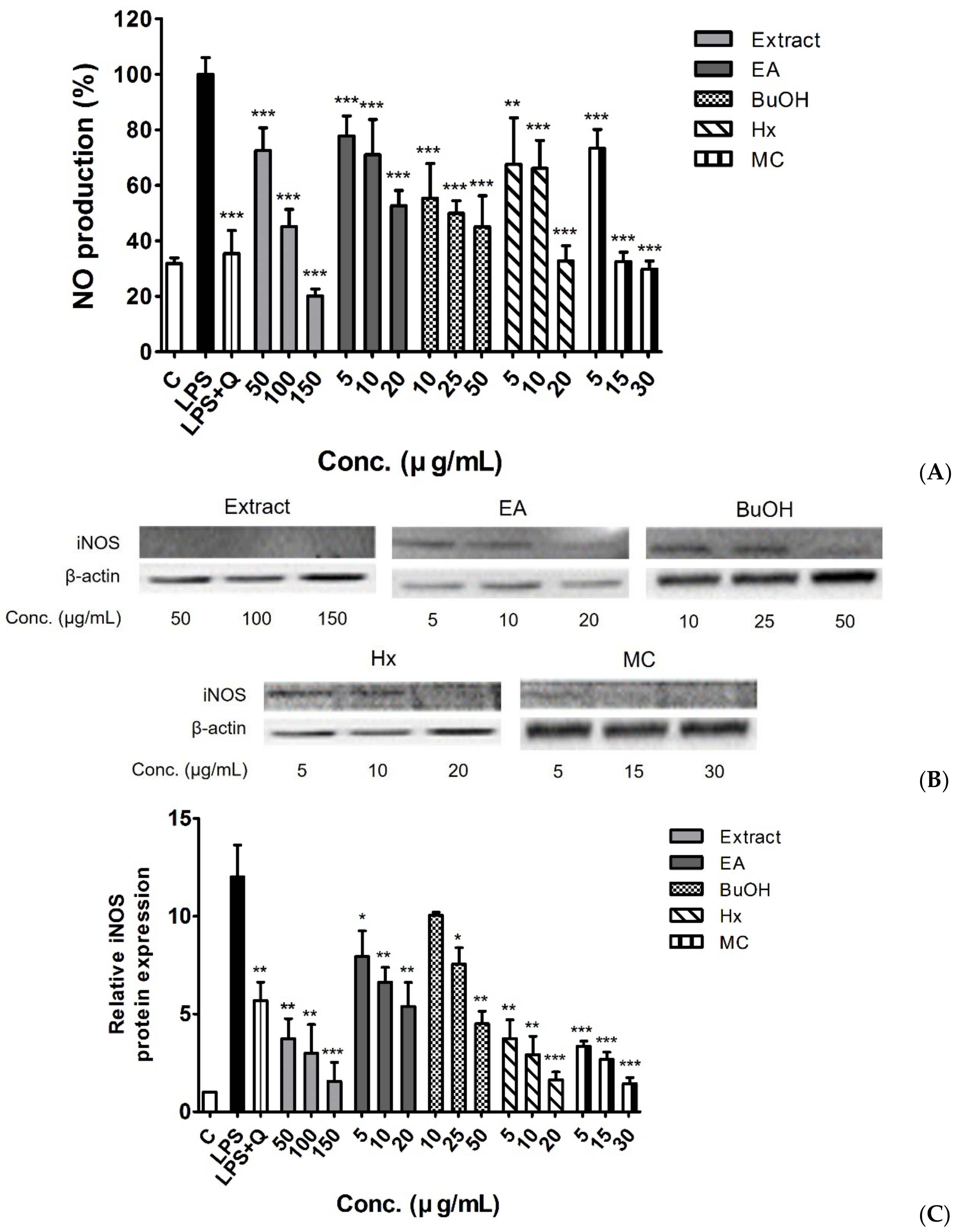
2.4. Anti-Inflammatory Activity
2.5. Mass Spectral Identification and Qualitative Analysis of Extracts
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Plant Material, Extraction, and Preparation of Fractions
4.3. Determination of Total Phenolic Content
4.4. Determination of Flavonoid Content
4.5. Cell Culture
4.6. Cell Viability
4.7. Antioxidant Activity
4.7.1. DPPH Radical Scavenging Assay
4.7.2. ABTS Radical Scavenging Assay
4.7.3. FRAP Assay
4.7.4. Measurement of Intracellular ROS Levels
4.7.5. Antioxidant Enzyme Capacity Assays
4.8. Anti-Inflammatory Activity
4.8.1. Measurement of Intracellular NO levels
4.8.2. Measurement of iNOS Protein Expression
4.9. Chemical Profiling and Qualitative UHPLC-QTOF-MS Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Sample Availability
References
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fialkow, L.; Wang, Y.; Downey, G.P. Reactive oxygen and nitrogen species as signaling molecules regulating neutrophil function. Free Radic. Biol. Med. 2007, 42, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, H.; Nair, J. Chronic inflammation and oxidative stress in the genesis and perpetuation of cancer: role of lipid peroxidation, DNA damage, and repair. Langenbecks Arch. Surg. 2006, 391, 499–510. [Google Scholar] [CrossRef]
- Prabhakar, O. Cerebroprotective effect of resveratrol through antioxidant and anti-inflammatory effects in diabetic rats. Naunyn. Schmiedebergs Arch. Pharmacol. 2013, 386, 705–710. [Google Scholar] [CrossRef]
- Kang, H.-J.; Jeong, J.-S.; Park, N.-J.; Go, G.-B.; Kim, S.O.; Park, C.; Kim, B.W.; Hong, S.-H.; Choi, Y.H. An ethanol extract of Aster yomena (Kitam.) Honda inhibits lipopolysaccharide-induced inflammatory responses in murine RAW 264.7 macrophages. Biosci. Trends 2017, 11, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afrin, S.; Gasparrini, M.; Forbes-Hernández, T.Y.; Cianciosi, D.; Reboredo-Rodriguez, P.; Manna, P.P.; Battino, M.; Giampieri, F. Protective effects of Manuka honey on LPS-treated RAW 264.7 macrophages. Part 1: Enhancement of cellular viability, regulation of cellular apoptosis and improvement of mitochondrial functionality. Food Chem. Toxicol. 2018, 121, 203–213. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, S.; Wang, L.; Yu, L.; Zhao, X.; Ni, H.; Wang, Y.; Wang, J.; Shan, C.; Fu, Y. Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells. Molecules 2020, 25, 1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Ji, D.; Turgeon, R.; Chen, J.; Lin, T.; Huang, J.; Luo, J.; Zhu, Y.; Zhang, C.; Lv, Z. Physiological and Proteomic Responses of Mulberry Trees (Morus alba. L.) to Combined Salt and Drought Stress. Int. J. Mol. Sci. 2019, 20, 2486. [Google Scholar] [CrossRef] [Green Version]
- Ramappa, V.K.; Srivastava, D.; Singh, P.; Kumar, U.; Kumar, D.; Gosipatala, S.B.; Saha, S.; Kumar, D.; Raj, R. Mulberries: A Promising Fruit for Phytochemicals, Nutraceuticals, and Biological Activities. Int. J. Fruit Sci. 2020, 1–26. [Google Scholar] [CrossRef]
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass spectrometry-based metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef]
- Yu, J.S.; Lee, D.; Lee, S.R.; Lee, J.W.; Choi, C.-I.; Jang, T.S.; Kang, K.S.; Kim, K.H. Chemical characterization of cytotoxic indole acetic acid derivative from mulberry fruit (Morus alba L.) against human cervical cancer. Bioorganic Chem. 2018, 76, 28–36. [Google Scholar] [CrossRef]
- Lee, S.R.; Park, J.Y.; Yu, J.S.; Lee, S.O.; Ryu, J.-Y.; Choi, S.-Z.; Kang, K.S.; Yamabe, N.; Kim, K.H. Odisolane, a novel oxolane derivative, and antiangiogenic constituents from the fruits of mulberry (Morus alba L.). J. Agric. Food Chem. 2016, 64, 3804–3809. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Lee, M.-S.; Choi, A.-J.; Kim, C.-T.; Kim, Y. Anti-Inflammatory effects of high hydrostatic pressure extract of mulberry (Morus alba) fruit on LPS-stimulated RAW264. 7 cells. Molecules 2019, 24, 1425. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-O.; Yu, M.-H.; Lee, Y.-J.; Leem, H.-H.; Kim, S.; Kang, D.-H.; Choi, S.-W. Comparison of functional constituents and biological activity of the seed extracts from two mulberry fruits. Prev. Nutr. Food Sci. 2010, 15, 98–104. [Google Scholar] [CrossRef]
- Kim, S.B.; Chang, B.Y.; Hwang, B.Y.; Kim, S.Y.; Lee, M.K. Pyrrole alkaloids from the fruits of Morus alba. Bioorg. Med. Chem. Lett. 2014, 24, 5656–5659. [Google Scholar] [CrossRef]
- Chu, Q.; Lin, M.; Tian, X.; Ye, J. Study on capillary electrophoresis–amperometric detection profiles of different parts of Morus alba L. J. Chromatogr. A 2006, 1116, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Perez, J.L.; Jayaprakasha, G.K.; Patil, B.S. Metabolite profiling and in vitro biological activities of two commercial bitter melon (Momordica charantia Linn.) cultivars. Food Chem. 2019, 288, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-J.; Park, E.-J. Feature analysis of different in vitro antioxidant capacity assays and their application to fruit and vegetable samples. J. Korean Soc. Food Sci. Nutr. 2011, 40, 1053–1062. [Google Scholar] [CrossRef] [Green Version]
- Molyneux, P. The use of the stable free radical diphenylpicrylhydrazyl (DPPH) for estimating antioxidant activity. Songklanakarin. J. Sci. Technol. 2004, 26, 211–219. [Google Scholar]
- Awika, J.M.; Rooney, L.W.; Wu, X.; Prior, R.L.; Cisneros-Zevallos, L. Screening methods to measure antioxidant activity of sorghum (Sorghum bicolor) and sorghum products. J. Agric. Food Chem. 2003, 51, 6657–6662. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xiang, L.; Wang, C.; Tang, C.; He, X. Antidiabetic and Antioxidant Effects and Phytochemicals of Mulberry Fruit (Morus Alba L.) Polyphenol Enhanced Extract. PLoS ONE 2013, 8, e71144. [Google Scholar] [CrossRef]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, M.L.; Liu, J.; Wang, Y.; Hu, J.H.; Wang, M.-H. Sonchus asper extract inhibits LPS-induced oxidative stress and pro-inflammatory cytokine production in RAW264. 7 macrophages. Nutr. Res. Pract. 2015, 9, 579–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, Y.-J.; Jeong, M.; Lee, K.-T.; Jang, D.S.; Choi, J.-H. Isocyperol, isolated from the rhizomes of Cyperus rotundus, inhibits LPS-induced inflammatory responses via suppression of the NF-κB and STAT3 pathways and ROS stress in LPS-stimulated RAW 264.7 cells. Int. Immunopharmacol. 2016, 38, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, M.; Forbes-Hernandez, T.Y.; Giampieri, F.; Afrin, S.; Alvarez-Suarez, J.M.; Mazzoni, L.; Mezzetti, B.; Quiles, J.L.; Battino, M. Anti-inflammatory effect of strawberry extract against LPS-induced stress in RAW 264.7 macrophages. Food Chem. Toxicol. 2017, 102, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Magar, T.B.T.; Pun, N.T.; Lee, Y.; Kim, E.H.; Lee, E.-S.; Park, P.-H. YJI-7 Suppresses ROS Production and Expression of Inflammatory Mediators via Modulation of p38MAPK and JNK Signaling in RAW 264.7 Macrophages. Biomol. Ther. 2018, 26, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, P.; Wadhwani, A. Antioxidant enzymes and human health. Antioxid. Enzyme 2012, 1, 3–18. [Google Scholar]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.E.; Ju, E.M.; Kim, J.H. Antioxidant activity of extracts from Euryale ferox seed. Exp. Mol. Med. 2002, 34, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Moncada, S.; Higgs, E.A. Molecular mechanisms and therapeutic strategies related to nitric oxide. FASEB J. 1995, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Kiss, J.P.; Vizi, E.S. Nitric oxide: a novel link between synaptic and nonsynaptic transmission. Trends Neurosci. 2001, 24, 211–215. [Google Scholar] [CrossRef]
- Förstermann, U.; Nakane, M.; Tracey, W.R.; Pollock, J.S. Isoforms of nitric oxide synthase: functions in the cardiovascular system. Eur. Heart J. 1993, 14, 10. [Google Scholar]
- Boscá, L.; Zeini, M.; Través, P.G.; Hortelano, S. Nitric oxide and cell viability in inflammatory cells: a role for NO in macrophage function and fate. Toxicology 2005, 208, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Joo, T.; Sowndhararajan, K.; Hong, S.; Lee, J.; Park, S.-Y.; Kim, S.; Jhoo, J.-W. Inhibition of nitric oxide production in LPS-stimulated RAW 264.7 cells by stem bark of Ulmus pumila L. Saudi J. Biol. Sci. 2014, 21, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Lakhanpal, P.; Rai, D.K. Quercetin: A versatile flavonoid. Internet J. Med. Update 2007, 2, 22–37. [Google Scholar] [CrossRef] [Green Version]
- López-Lázaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef]
- Calderon-Montano, J.M.; Burgos-Morón, E.; Pérez-Guerrero, C.; López-Lázaro, M. A review on the dietary flavonoid kaempferol. Mini Rev. Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef]
- Riaz, A.; Rasul, A.; Hussain, G.; Zahoor, M.K.; Jabeen, F.; Subhani, Z.; Younis, T.; Ali, M.; Sarfraz, I.; Selamoglu, Z. Astragalin: A bioactive phytochemical with potential therapeutic activities. Adv. Pharmacol. Sci. 2018, 2018. [Google Scholar] [CrossRef]
- Sunil, C.; Xu, B. An insight into the health-promoting effects of taxifolin (dihydroquercetin). Phytochemistry 2019, 166, 112066. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Kumar, P.; Pathak, D. Biological importance of the indole nucleus in recent years: a comprehensive review. J. Heterocycl. Chem. 2010, 47, 491–502. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Huang, K.-J.; Wang, S.-K.; Wen, Z.-H.; Chen, P.-W.; Duh, C.-Y. Antiviral and anti-inflammatory metabolites from the soft coral Sinularia capillosa. J. Nat. Prod. 2010, 73, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Kang, M.-C.; Lee, K.-W.; Kang, S.-M.; Lee, W.-W.; Jeon, Y.-J. Antioxidant activity and cell protective effect of loliolide isolated from Sargassum ringgoldianum subsp. coreanum. Algae 2011, 26, 201. [Google Scholar] [CrossRef]
- Machado, F.B.; Yamamoto, R.E.; Zanoli, K.; Nocchi, S.R.; Novello, C.R.; Schuquel, I.T.A.; Sakuragui, C.M.; Luftmann, H.; Ueda-Nakamura, T.; Nakamura, C.V. Evaluation of the antiproliferative activity of the leaves from Arctium lappa by a bioassay-guided fractionation. Molecules 2012, 17, 1852–1859. [Google Scholar] [CrossRef]
- Park, S.H.; Choi, E.; Kim, S.; Kim, D.S.; Kim, J.H.; Chang, S.; Choi, J.S.; Park, K.J.; Roh, K.-B.; Lee, J. Oxidative stress-protective and anti-melanogenic effects of loliolide and ethanol extract from fresh water green algae, Prasiola japonica. Int. J. Mol. Sci. 2018, 19, 2825. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Kim, D.S.; Kim, S.; Lorz, L.R.; Choi, E.; Lim, H.Y.; Hossain, M.A.; Jang, S.; Choi, Y.I.; Park, K.J. Loliolide presents antiapoptosis and antiscratching effects in human keratinocytes. Int. J. Mol. Sci. 2019, 20, 651. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-H.; Kwak, C.-H.; Lee, S.-K.; Ha, S.-H.; Park, J.; Chung, T.-W.; Ha, K.-T.; Suh, S.-J.; Chang, Y.-C.; Chang, H.W. Anti-inflammatory effect of ascochlorin in LPS-stimulated RAW 264.7 macrophage cells is accompanied with the down-regulation of iNOS, COX-2 and proinflammatory cytokines through NF-κB, ERK1/2, and p38 signaling pathway. J. Cell. Biochem. 2016, 117, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-W.; West, X.Z.; Byzova, T.V. Inflammation and oxidative stress in angiogenesis and vascular disease. J. Mol. Med. 2013, 91, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T.; Fong, C. The resolution of inflammation: Anti-inflammatory roles for NF-κB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef]
- Hong, S.C.; Jeong, J.B.; Koo, J.S. Inhibitory effect of essential oil from Agastache rugosa against nitric oxide (NO) production induced by inducible nitric oxide synthase (iNOS) over-expression through NF-B and mitogen-activated protein kinase (MAPK) activation in lipopolysaccharide (LPS)-stimulated RAW264. 7 cells. J. Med. Plants Res. 2012, 6, 4494–4500. [Google Scholar]
- Li, J.; Xu, L.; Sang, R.; Yu, Y.; Ge, B.; Zhang, X. Immunomodulatory and anti-inflammatory effects of total flavonoids of Astragalus by regulating NF-KB and MAPK signalling pathways in RAW 264.7 macrophages. Pharm. Int. J. Pharm. Sci. 2018, 73, 589–593. [Google Scholar]
- Monmai, C.; Go, S.H.; Shin, I.; You, S.; Lee, H.; Kang, S.; Park, W.J. Immune Enhancement Effect of Asterias amurensis Fatty Acids through NF-κB and MAPK Pathways on RAW 264.7 Cells. J. Microbiol. Biotechnol. 2018, 28, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Yang, E.-J.; Hyun, K.H.; Kim, H.; Kim, M.-J.; Lee, N.H.; Hyun, C.-G. Acanthopanax koreanum roots inhibit the expression of pro-inflammatory cytokines, inducible nitric oxide synthase, and cyclooxygenase-2 in RAW 264.7 macrophages. Orient. J. Chem. 2016, 32, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Ryoo, R.; Choi, J.H.; Kim, J.-H.; Kim, S.-H.; Kim, K.H. Trichothecene and tremulane sesquiterpenes from a hallucinogenic mushroom Gymnopilus junonius and their cytotoxicity. Arch. Pharm. Res. 2020, 43, 214–223. [Google Scholar] [CrossRef]
- Trinh, T.A.; Park, E.J.; Lee, D.; Song, J.H.; Lee, H.L.; Kim, K.H.; Kim, Y.; Kung, K.; Kang, K.S.; Yoo, J.E. Estrogenic Activity of Sanguiin H-6 through Activation of Estrogen Receptor α Coactivator-binding Site. Nat. Prod. Sci. 2019, 25, 28–33. [Google Scholar] [CrossRef]
- Lee, S.R.; Kang, H.; Yoo, M.J.; Yu, J.S.; Lee, S.; Yi, S.A.; Beemelmanns, C.; Lee, J.; Kim, K.H. Anti-adipogenic pregnane steroid from a Hydractinia-associated fungus, Cladosporium sphaerospermum SW67. Nat. Prod. Sci. 2020, 26, 230–235. [Google Scholar]
- Lee, K.; Seo, Y.-J.; Song, J.-H.; Chei, S.; Lee, B.-Y. Ginsenoside Rg1 promotes browning by inducing UCP1 expression and mitochondrial activity in 3T3-L1 and subcutaneous white adipocytes. J. Ginseng Res. 2019, 43, 589–599. [Google Scholar] [CrossRef]
- Blois, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Arts, M.J.; Haenen, G.R.; Voss, H.-P.; Bast, A. Antioxidant capacity of reaction products limits the applicability of the Trolox Equivalent Antioxidant Capacity (TEAC) assay. Food Chem. Toxicol. 2004, 42, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Sittisart, P.; Chitsomboon, B. Intracellular ROS Scavenging Activity and Downregulation of Inflammatory Mediators in RAW264.7 Macrophage by Fresh Leaf Extracts of Pseuderanthemum palatiferum. Evid. Based Complement. Alternat. Med. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jo, M.S.; Lee, S.; Yu, J.S.; Baek, S.C.; Cho, Y.-C.; Kim, K.H. Megastigmane Derivatives from the Cladodes of Opuntia humifusa and Their Nitric Oxide Inhibitory Activities in Macrophages. J. Nat. Prod. 2020, 83, 684–692. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Phenolic (mg GAE/g) | Flavonoid (mg CAE/g) | DPPH (IC50, μg/mL) | ABTS (IC50, μg/mL) | FRAP (mmol Fe2+/g) | |
|---|---|---|---|---|---|
| EtOH extract | 173.1 ± 3.9 | 68.5 ± 8.2 | > 1000 | 890.6 ± 56.6 | 0.770 ± 0.019 |
| EA fraction | 177.9 ± 4.7 | 74.9 ± 8.6 | 133.6 ± 4.7 | 216.6 ± 28.8 | 3.727 ± 0.055 |
| BuOH fraction | 204.3 ± 4.7 | 70.6 ± 6.9 | 471.1 ± 35.3 | 793.2 ± 8.4 | 1.259 ± 0.026 |
| HX fraction | 102.0 ± 1.5 | 55.1 ± 9.6 | > 1000 | > 1000 | 0.505 ± 0.017 |
| MC fraction | 124.7 ± 0.6 | 64.3 ± 2.8 | 475.0 ± 18.4 | 218.1 ± 22.6 | 1.649 ± 0.023 |
| SOD (% of Control) | GPx (nmol/min) | CAT (unit/mL) | |
|---|---|---|---|
| Control (0.5% DMSO) | 100.0 | 2.674 ± 0.029 | 0.147 ± 0.007 |
| LPS 2 μg/mL | 64.6 ± 0.2 ### | 2.292 ± 0.092 ## | 0.120 ± 0.007 ## |
| LPS + Quercetin (10 μM) | 93.7 ± 5.1 *** | 2.547 ± 0.039 ** | 0.163 ± 0.005 ** |
| LPS + EtOH Extract (50 μg/mL) | 78.7 ± 3.2 ** | 2.776 ± 0.025 *** | 0.156 ± 0.011 * |
| LPS + EA fraction (5 μg/mL) | 91.1 ± 5.7 ** | 2.674 ± 0.039 ** | 0.142 ± 0.004 * |
| LPS + BuOH fraction (10 μg/mL) | 85.7 ± 5.1 ** | 2.521 ± 0.044 ** | 0.157 ± 0.010 ** |
| LPS + HX fraction (5 μg/mL) | 85.6 ± 2.0 *** | 2.471 ± 0.015 * | 0.162 ± 0.011 ** |
| LPS + MC fraction (5 μg/mL) | 89.1 ± 1.9 *** | 2.700 ± 0.044 ** | 0.146 ± 0.012 * |
| Fractions | Compound | Formula | Theoretical (m/z) | Experimental (m/z) | Adduct | Δ (ppm) a |
|---|---|---|---|---|---|---|
| Hexane | Ar-Turmerone | C15H20O | 217.1592 | 217.1585 | M + H | −3.22 |
| (2S,4aR,4bR,6aR,8S,10aR,10bR,12aS)-8-(Acetyloxy)hexadecahydro-2,4a,4b,7,7,10a-hexamethyl-2-(3-oxobutyl)-1(2H)-chrysenone | C30H48O4 | 473.3631 | 473.3621 | M + H | −2.11 | |
| Stigmasterol | C29H48O | 413.3783 | 413.3779 | M + H | −9.68 | |
| Methylene chloride | Odisolane | C8H14O4 | 175.097 | 175.0979 | M + H | 5.14 |
| 3-Benzofurancarboxyaldehyde | C9H6O2 | 147.0446 | 147.0437 | M + H | −6.12 | |
| Loliolide | C11H16O3 | 197.1172 | 197.1178 | M + H | 3.04 | |
| (R)-5-Hydroxypyrrolidin-2-one | C4H7NO2 | 102.0547 | 102.0555 | M + H | 7.84 | |
| Methyl R-pyroglutamate | C6H9NO3 | 144.0661 | 144.0655 | M + H | −4.16 | |
| Indole | C8H7N | 118.0657 | 118.0646 | M + H | −9.32 | |
| Ethyl acetate | Quercetin | C15H10O7 | 303.0505 | 303.0499 | M + H | −1.98 |
| Kaempferol | C15H10O6 | 287.0556 | 287.0549 | M + H | −2.44 | |
| Luteolin | C15H10O6 | 287.0556 | 287.0546 | M + H | −3.48 | |
| Astragalin | C21H20O11 | 449.1084 | 449.1082 | M + H | −4.45 | |
| Taxifolin | C15H12O7 | 305.0661 | 305.0656 | M + H | −1.64 | |
| Morrole A | C14H21NO5 | 284.1498 | 284.1489 | M + H | −3.17 | |
| Methyl chlorogenate | C17H20O9 | 369.1186 | 369.1181 | M + H | −1.35 | |
| n-Butanol | Quercetin 3-O-β-glucoside | C21H20O12 | 465.1033 | 465.1028 | M + H | −1.08 |
| Kaempferol 3-O-β-rutinoside | C27H30O15 | 595.1663 | 595.1659 | M + H | −6.72 | |
| Rutin | C27H30O16 | 611.1612 | 611.1609 | M + H | −4.91 | |
| Butyl L-pyroglutamate | C9H15NO3 | 186.113 | 186.1127 | M + H | −1.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.S.; Lim, S.H.; Lee, S.R.; Choi, C.-I.; Kim, K.H. Antioxidant and Anti-Inflammatory Effects of White Mulberry (Morus alba L.) Fruits on Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. Molecules 2021, 26, 920. https://doi.org/10.3390/molecules26040920
Yu JS, Lim SH, Lee SR, Choi C-I, Kim KH. Antioxidant and Anti-Inflammatory Effects of White Mulberry (Morus alba L.) Fruits on Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. Molecules. 2021; 26(4):920. https://doi.org/10.3390/molecules26040920
Chicago/Turabian StyleYu, Jae Sik, Sung Ho Lim, Seoung Rak Lee, Chang-Ik Choi, and Ki Hyun Kim. 2021. "Antioxidant and Anti-Inflammatory Effects of White Mulberry (Morus alba L.) Fruits on Lipopolysaccharide-Stimulated RAW 264.7 Macrophages" Molecules 26, no. 4: 920. https://doi.org/10.3390/molecules26040920