
Chloroquine and Hydroxychloroquine Interact Differently with ACE2 Domains Reported to Bind with the Coronavirus Spike Protein: Mediation by ACE2 Polymorphism




Abstract
:1. Introduction
2. Materials and Methods
2.1. Structure and Genetic ACE2 Polymorphism
2.2. In Silico Approach of ADME, Pharmacokinetics, and Docking Study
3. Results and Discussion
3.1. ACE2 Coding Variants
3.2. Molecular Binding and Interaction Results
3.3. Pharmacokinetics and ADME Findings of CQ and HCQ
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Jorge, A.; Ung, C.; Young, L.H.; Melles, R.B.; Choi, H.K. Hydroxychloroquine retinopathy—Implications of research advances for rheumatology care. Nat. Rev. Rheumatol. 2018, 14, 693–703. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Ji, Z.; Xu, C.; Zhu, J. The clinical value of using chloroquine or hydroxychloroquine as autophagy inhibitors in the treatment of cancers: A systematic review and meta-analysis. Medicine 2018, 97, e12912. [Google Scholar] [CrossRef] [PubMed]
- Lammers, A.; Brohet, R.; Theunissen, R.; Koster, C.; Rood, R.; Verhagen, D.; Brinkman, K.; Hassing, R.; Dofferhoff, A.; El Moussaoui, R.; et al. Early hydroxychloroquine but not chloroquine use reduces ICU admission in COVID-19 patients. Int. J. Infect. Dis. 2020, 101, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Jain, M.; Reddy, R.P.; Jain, R. Quinolines and structurally related heterocycles as antimalarials. Eur. J. Med. Chem. 2010, 45, 3245–3264. [Google Scholar] [CrossRef] [PubMed]
- Badraoui, R.; Alrashedi, M.M.; El-May, M.V.; Bardakci, F. Acute respiratory distress syndrome: A life threatening associated complication of SARS-CoV-2 infection inducing COVID-19. J. Biomol. Struct. Dyn. 2020, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.J.; Boyer, T.L.; Venner, E.; Beck, P.J.; Slamowitz, T.; Caste, T.; Hickman, A.; Raymond, M.H.; Costa-Pinheiro, P.; Jameson, M.J.; et al. Inhibition of Lysosomal Function Mitigates Protective Mitophagy and Augments Ceramide Nanoliposome–Induced Cell Death in Head and Neck Squamous Cell Carcinoma. Mol. Cancer Ther. 2020, 19, 2621–2633. [Google Scholar] [CrossRef]
- Liang, S.; Li, X.; Gao, C.; Zhang, L. microRNA-based autophagy inhibition as targeted therapy in pancreatic cancer. Biomed. Pharmacother. 2020, 132, 110799. [Google Scholar] [CrossRef]
- Pimentel, J.; Andersson, N. Chloroquine and its derivatives in the management of COVID-19: A scoping review. Biomedica 2020, 40, 80–95. [Google Scholar] [CrossRef]
- Ziegler, H.K.; Unanue, E.R. Decrease in macrophage antigen catabolism caused by ammonia and chloroquine is associated with inhibition of antigen presentation to T cells. Proc. Natl. Acad. Sci. USA 1982, 79, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Gies, V.; Bekaddour, N.; Dieudonné, Y.; Guffroy, A.; Frenger, Q.; Gros, F.; Rodero, M.P.; Herbeuval, J.-P.; Korganow, A.-S. Beyond anti-viral effects of chloroquine/hydroxychloroquine. Front. Immunol. 2020, 11, 1409. [Google Scholar] [CrossRef]
- Wang, N.; Han, S.; Liu, R.; Meng, L.; He, H.; Zhang, Y.; Wang, C.; Lv, Y.; Wang, J.; Li, X.; et al. Chloroquine and hydroxychloroquine as ACE2 blockers to inhibit viropexis of 2019-nCoV Spike pseudotyped virus. Phytomedicine 2020, 79, 153333. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci. China Life Sci. 2020, 63, 457–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of the SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhao, Z.; Wang, Y.; Zhou, Y.; Ma, Y.; Zuo, W. Single-cell RNA expression profiling of ACE2, the putative receptor of Wuhan 2019-nCov. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zou, X.; Chen, K.; Zou, J.; Han, P.; Hao, J.; Han, Z. Single-cell RNA-seq data analysis on the receptor ACE2 expression reveals the potential risk of different human organs vulnerable to 2019-nCoV infection. Front. Med. 2020, 14, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Donoghue, M.; Hsieh, F.; Baronas, E.; Godbout, K.; Gosselin, M.; Stagliano, S.; Donovan, M.; Woolf, B.; Robison, K.; Jeyaseelan, R.; et al. A novel angiotensin-converting enzyme-related carboxypeptidase (ACE2) converts angiotensin I to angiotensin 1–9. Circ. Res. 2000, 87, E1–E9. [Google Scholar] [CrossRef]
- Delanghe, J.R.; Speeckaert, M.M.; De Buyzere, M.L. COVID-19 infections are also affected by human ACE1 D/I polymorphism. Clin. Chem. Lab. Med. 2020, 58, 1125–1126. [Google Scholar] [CrossRef] [Green Version]
- Delanghe, J.R.; Speeckaert, M.M.; De Buyzere, M.L. The host’s angiotensin-converting enzyme polymorphism may explain epidemiological findings in COVID-19 infections. Clin. Chim. Acta 2020, 505, 192–193. [Google Scholar] [CrossRef]
- Knoll, M.D.; Wonodi, C. Oxford-AstraZeneca COVID-19 vaccine efficacy. Lancet 2021, 397, 72–74. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Zhu, H.; Lu, H. The development of a quarantine strategy is an important path to a normalized response to COVID-19. Biosci. Trends 2020, 14, 396–398. [Google Scholar] [CrossRef] [PubMed]
- Abdelli, I.; Hassani, F.; Brikci, S.B.; Ghalem, S. In silico study the inhibition of angiotensin converting enzyme 2 receptor of COVID-19 by Ammoides verticillata components harvested from Western Algeria. J. Biomol. Struct. Dyn. 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.J.; Danciu, C.; Ashraf, S.A.; Moin, A.; Singh, R.; Alreshidi, M.; Patel, M.; Jahan, S.; Kumar, S.; Alkhinjar, M.I.M.; et al. Plants-derived biomolecules as potent antiviral phytomedicines: New insights on ethnobotanical evidences against coronaviruses. Plants 2020, 9, 1244. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.E.; McLaren, W.; Gil, L.; Thormann, A.; Schuilenburg, H.; Sheppard, D.; Parton, A.; Armean, I.M.; Trevanion, S.J.; Flicek, P.; et al. Ensembl variation resources. Database 2018, 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Newman, V.; Moore, B.; Sparrow, H.; Perry, E. The Ensembl Genome Browser: Strategies for Accessing Eukaryotic Genome Data. Methods Mol. Biol. 2018, 1757, 115–139. [Google Scholar] [CrossRef] [Green Version]
- Lek, M.; Karczewski, K.J.; Exome Aggregation Consortium; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Hussain, M.; Jabeen, N.; Raza, F.; Shabbir, S.; Baig, A.A.; Amanullah, A.; Aziz, B. Structural variations in human ACE2 may influence its binding with SARS-CoV-2 spike protein. J. Med. Virol. 2020, 92, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Goodsell, D.S.; Zardecki, C.; Di Costanzo, L.; Duarte, J.M.; Hudson, B.P.; Persikova, I.; Segura, J.; Shao, C.; Voigt, M.; Westbrook, J.D.; et al. RCSB Protein Data Bank: Enabling biomedical research and drug discovery. Protein Sci. 2020, 29, 52–65. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.D.; Achuthan, P.; Akanni, W.; Allen, J.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; et al. Ensembl 2020. Nucleic Acids Res. 2020, 48, D682–D688. [Google Scholar] [CrossRef] [PubMed]
- Paladugu, S.; Donato, A.A. Remdesivir improved time to recovery in adults hospitalized with COVID-19 and lower respiratory tract involvement. Ann. Intern. Med. 2020, 173, JC4. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, C.; Sui, J.; Kuhn, J.H.; Moore, M.J.; Luo, S.; Wong, S.-K.; Huang, I.-C.; Xu, K.; Vasilieva, N.; et al. Receptor and viral determinants of SARScoronavirus adaptation to human ACE2. EMBO J. 2005, 24, 1634–1643. [Google Scholar] [CrossRef] [Green Version]
- Mudgal, R.; Nehul, S.; Tomar, S. Prospects for mucosal vaccine: Shutting the door on SARS-CoV-2. Hum. Vaccin. Immunother. 2020, 16, 2921–2931. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.E.; Barbitoff, Y.A.; Glotov, A.S.; Danilova, M.M.; Tonyan, Z.N.; Nasykhova, Y.A.; Mikhailova, A.A.; Bespalova, O.N.; Kalinin, R.S.; Mirzorustamova, A.M.; et al. Analysis of the Spectrum of ACE2 Variation Suggests a Possible Influence of Rare and Common Variants on Susceptibility to COVID-19 and Severity of Outcome. Front. Genet. 2020, 29, 551220. [Google Scholar] [CrossRef]
- Ali, F.; Elserafy, M.; Alkordi, M.H.; Amin, M. ACE2 coding variants in different populations and their potential impact on SARS-CoV-2 binding affinity. Biochem. Biophys. Rep. 2020, 24, 100798. [Google Scholar] [CrossRef]
- Novelli, A.; Biancolella, M.; Borgiani, P.; Cocciadiferro, D.; Colona, V.L.; D’Apice, M.R.; Rogliani, P.; Zaffina, S.; Leonardis, F.; Campana, A.; et al. Analysis of ACE2 genetic variants in 131 Italian SARS-CoV-2-positive patients. Hum. Genom. 2020, 14, 29. [Google Scholar] [CrossRef]
- Sinha, M.; Dola, V.R.; Soni, A.; Agarwal, P.; Srivastava, K.; Haq, W.; Puri, S.K.; Katti, S.B. Synthesis of chiral chloroquine and its analogues as antimalarial agents. Bioorganic Med. Chem. 2014, 22, 5950–5960. [Google Scholar] [CrossRef]
- Skariyachan, S.; Gopal, D.; Chakrabarti, S.; Kempanna, P.; Uttarkar, A.; Muddebihalkar, A.G.; Niranjan, V. Structural and molecular basis of the interaction mechanism of selected drugs towards multiple targets of SARS-CoV-2 by molecular docking and dynamic simulation studies- deciphering the scope of repurposed drugs. Comput. Biol. Med. 2020, 126, 104054. [Google Scholar] [CrossRef]
- Fantini, J.; Di Scala, C.; Chahinian, H.; Yahi, N. Structural and molecular modelling studies reveal a new mechanism of action of chloroquine and hydroxychloroquine against SARS-CoV-2 infection. Int. J. Antimicrob. Agent 2020, 55, 105960. [Google Scholar] [CrossRef]
- Akacha, A.; Badraoui, R.; Rebai, T.; Zourgui, L. Effect of Opuntia ficus indica extract on methotrexate-induced testicular injury: A biochemical, docking and histological study. J. Biomol. Struct. Dyn. 2020, 1856187. [Google Scholar] [CrossRef]
- Hwang, D.; Ramsey, J.D.; Makita, N.; Sachse, C.; Jordan, R.; Sokolsky-Papkov, M.; Kabanov, A.V. Novel poly(2-oxazoline) block copolymer with aromatic heterocyclic side chains as a drug delivery platform. J. Control. Release 2019, 307, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Saab, Y.B.; Gard, P.; Overall, A. The geographic distribution of the ACE II genotype: A novel finding. Genet. Res. 2007, 89, 259–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badraoui, R.; Rebai, T.; Elkahoui, S.; Alreshidi, M.; Veettil, V.N.; Noumi, E.; Al-Motair, K.A.; Aouadi, K.; Kadri, A.; De Feo, V.; et al. Allium subhirsutum L. as a Potential Source of Antioxidant and Anticancer Bioactive Molecules: HR-LCMS Phytochemical Profiling, In Vitro and In Vivo Pharmacological Study. Antioxidants 2020, 9, 1003. [Google Scholar] [CrossRef]
- Chatre, C.; Roubille, F.; Vernhet, H.; Jorgensen, C.; Pers, Y.M. Cardiac complications attributed to chloroquine and hydroxychloroquine: A systematic review of the literature. Drug Saf. 2018, 41, 919–931. [Google Scholar] [CrossRef]
- Babayeva, M.; Loewy, Z. Repurposing Drugs for COVID-19: Pharmacokinetics and Pharmacogenomics of Chloroquine and Hydroxychloroquine. Pharmgenomics Pers. Med. 2020, 13, 531–542. [Google Scholar] [CrossRef]
- Curreli, F.; Victor, S.M.B.; Ahmed, S.; Drelich, A.; Tong, X.; Tseng, C.-T.K.; Hillyer, C.D.; Debnath, A.K. Stapled Peptides Based on Human Angiotensin-Converting Enzyme 2 (ACE2) Potently Inhibit SARS-CoV-2 Infection In Vitro. mBio 2020, 11. [Google Scholar] [CrossRef]
- Flores-Holguín, N.; Frau, J.; Glossman-Mitnik, D. Conceptual DFT-Based Computational Peptidology of Marine Natural Compounds: Discodermins A–H. Molecules 2020, 25, 4158. [Google Scholar] [CrossRef]
- Guo, X.; Chen, Z.; Xia, Y.; Lin, W.; Li, H. Investigation of the genetic variation in ACE2 on the structural recognition by the novel coronavirus (SARS-CoV-2). J. Transl. Med. 2020, 18, 321. [Google Scholar] [CrossRef]
- Della Porta, A.; Bornstein, K.; Coye, A.; Montrief, T.; Long, B.; Parris, M.A. Acute chloroquine and hydroxychloroquine toxicity: A review for emergency clinicians. Am. J. Emerg. Med. 2020, 38, 2209–2217. [Google Scholar] [CrossRef]



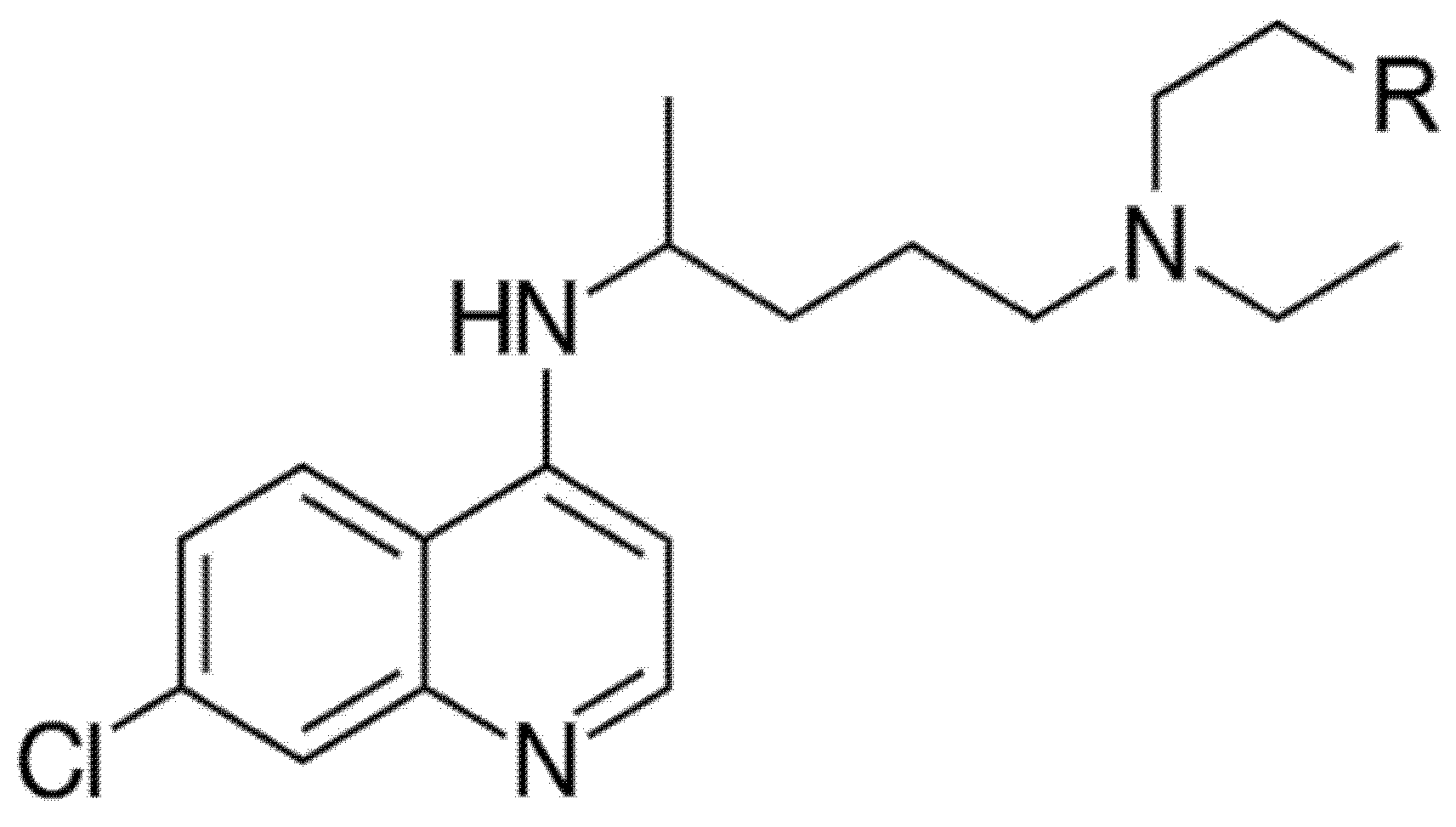{kind=link}
{kind=link}
| Variant No. | Genetic Variant | Amino Acid Change | Allele Frequency |
|---|---|---|---|
| 1 | rs4646116 | K26R | 3.88 × 10−3 |
| 2 | rs73635825 | S19P | 3.13 × 10−4 |
| 3 | rs146676783 | E37K | 3.9 × 10−5 |
| 4 | rs762890235 | P389H | 3.83 × 10−5 |
| 5 | rs143936283 | E329G | 3.44 × 10−5 |
| 6 | rs766996587 | M82I | 2.44 × 10−5 |
| 7 | rs1348114695 | E35K | 1.64 × 10−5 |
| 8 | rs961360700 | D355N | 1.17 × 10−5 |
| 9 | rs755691167 | K68E | 1.09 × 10−5 |
| 10 | rs1316056737 | D427Y | 1.09 × 10−5 |
| 11 | rs781255386 | T27A | 1.09 × 10−5 |
| 12 | rs1299103394 | K26E | 5.45 × 10−6 |
| 13 | rs759134032 | P84T | 5.47 × 10−6 |
| 14 | rs1238146879 | P426A | 5.47 × 10−6 |
| 15 | rs778500138 | E35D | N/A |
| 16 | rs1396769231 | M383T | N/A |
| 17 | rs1016777825 | R559S | N/A |
| No. | Genetic Variant | Chloroquine (CQ) | Hydroxychloroquine (HCQ) | ||||
|---|---|---|---|---|---|---|---|
| Affinity (Kcal/Mol) | Conventional H-Bonds | Number of Closest Interacting Residues | Affinity (Kcal/mol) | Conventional H-Bonds | Number of Closest Interacting Residues | ||
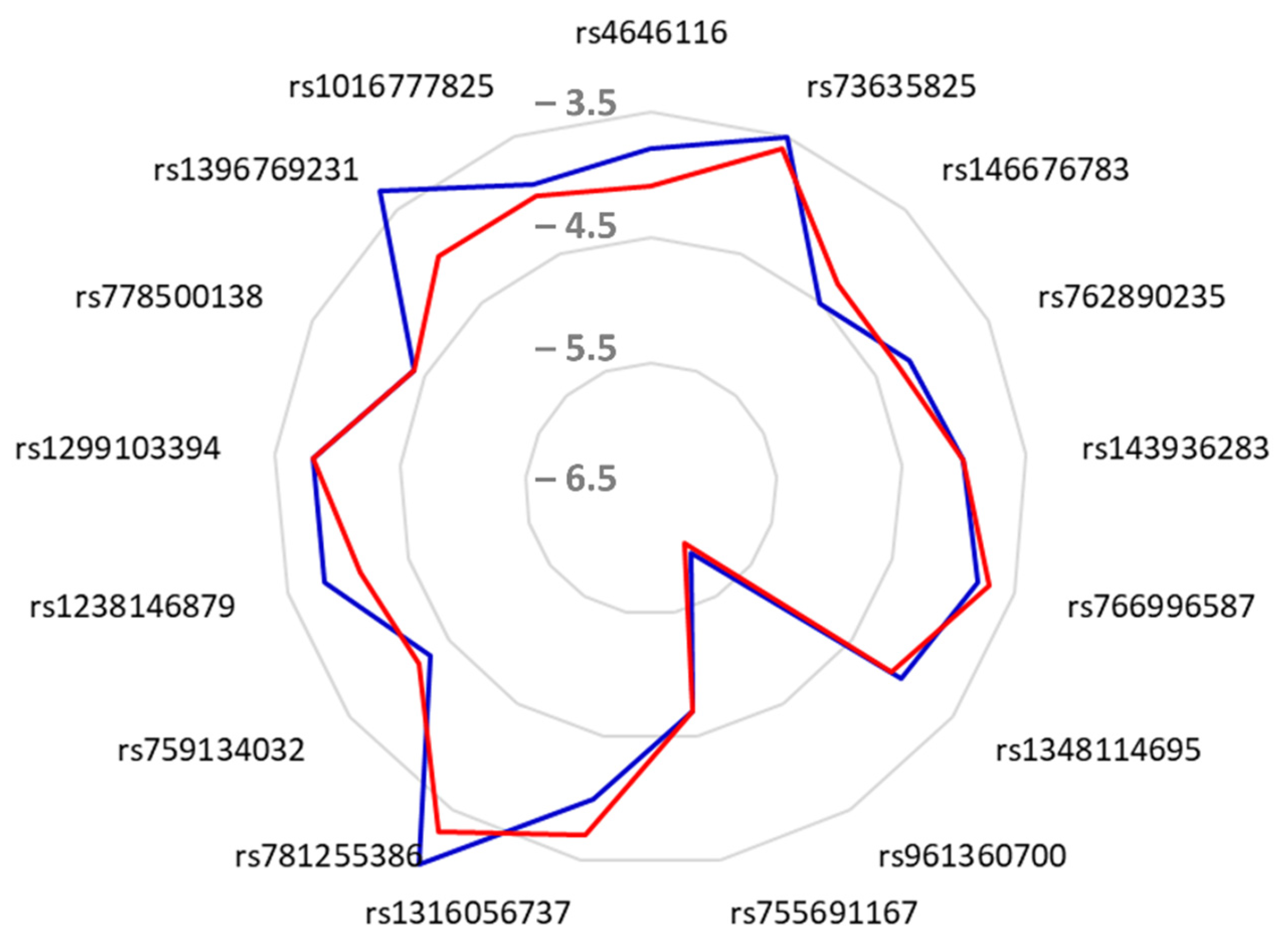
| 1 | rs4646116 | −3.8 | 2 | 4 | −4.1 | 2 | 4 |
| 2 | rs73635825 | −3.5 | 3 | 3 | −3.6 | 3 | 3 |
| 3 | rs146676783 | −4.5 | 2 | 4 | −4.3 | 2 | 4 |
| 4 | rs762890235 | −4.2 | 1 | 7 | −4.3 | 3 | 3 |
| 5 | rs143936283 | −4.0 | 1 | 6 | −4.0 | 2 | 5 |
| 6 | rs766996587 | −3.8 | 4 | 3 | −3.7 | 3 | 3 |
| 7 | rs1348114695 | −4.0 | 1 | 5 | −4.1 | 2 | 5 |
| 8 | rs961360700 | −5.9 | 2 | 6 | −6.0 | 4 | 7 |
| 9 | rs755691167 | −4.7 | 2 | 4 | −4.7 | 2 | 6 |
| 10 | rs1316056737 | −4.0 | 2 | 3 | −3.7 | 3 | 7 |
| 11 | rs781255386 | −3.0 | 1 | 4 | −3.3 | 3 | 3 |
| 12 | rs1299103394 | −3.8 | 4 | 2 | −3.8 | 4 | 4 |
| 13 | rs759134032 | −4.3 | 2 | 6 | −4.2 | 2 | 5 |
| 14 | rs1238146879 | −3.8 | 2 | 5 | −4.1 | 3 | 6 |
| 15 | rs778500138 | −4.4 | 2 | 4 | −4.4 | 3 | 2 |
| 16 | rs1396769231 | −3.3 | 3 | 6 | −4.0 | 5 | 6 |
| 17 | rs1016777825 | −3.9 | 2 | 3 | −4.0 | 2 | 4 |
| Receptor-Ligand Interactions, Distance in Angstroms and 2D Interactions Diagrams | Receptor-Ligand 3D interaction Microphotographs | |
|---|---|---|
| Chloroquine (CQ) | rs1299103394 (K26E)—CQ (CYS16)---(CQ) Conventional hydrogen bond: 3.702 Å; (ILE27)---(CQ) Conventional hydrogen bond: 2.269 Å; (SER10)---(CQ) Conventional hydrogen bond: 2.849 Å; (PHE15)---(CQ) Conventional hydrogen bond: 3.572 Å; (ALA18)---(CQ) Alkyl interaction: 4.047 Å; (CYS16)---(CQ) Alkyl interaction: 4.802 Å; (TRP37)---(CQ) Pi-Alkyl interaction: 5.195 Å.  |  |
| rs766996587 (M82I)—CQ (TYR33)---(CQ) Conventional hydrogen bond: 2.227 Å; (GLN51)---(CQ) Conventional hydrogen bond: 1.983 Å; (TYR33)---(CQ) Conventional hydrogen bond: 3.385 Å; (TYR33)---(CQ) Conventional hydrogen bond: 3.361 Å; (PRO34)---(CQ) Pi-Sigma interaction: 3.437 Å; (TYR33,PRO34)---(CQ) Amide-Pi Stacked interaction: 4.757 Å; (PRO34)---(CQ) Pi-Alkyl interaction: 5.295 Å.  |  | |
| Hydroxychloroquine (HCQ) | rs1396769231 (M383T)—HCQ (GLY28)---(HCQ) Conventional hydrogen bond: 2.900 Å; (SER48)---(HCQ) Conventional hydrogen bond: 2.271 Å; (SER48)---(HCQ) Conventional hydrogen bond: 3.177 Å; (GLY28)---(HCQ) Conventional hydrogen bond: 3.296 Å; (SER46)---(HCQ) Conventional hydrogen bond: 3.399 Å; (PRO53)---(HCQ) Alkyl interaction: 5.246 Å; (LYS27)---(HCQ) Alkyl interaction: 4.494 Å; (ARG49)---(HCQ) Pi-Alkyl interaction: 5.201 Å; (PRO53)---(HCQ) Pi-Alkyl interaction: 5.246 Å; (PRO53)---(HCQ) Pi-Alkyl interaction: 3.918 Å.  |  |
| rs961360700 (D355N)—HCQ (THR26)---(HCQ) Conventional hydrogen bond: 2.344 Å; (THR26)---(HCQ) Conventional hydrogen bond: 2.537 Å; (HIS24)---(HCQ) Conventional hydrogen bond: 3.377 Å; (THR26)---(HCQ) Conventional hydrogen bond: 3.270 Å; (MET11)---(HCQ) Carbon hydrogen bond: 3.494 Å; (VAL22)---(HCQ) Carbon hydrogen bond: 3.455 Å; (TRP28)---(HCQ) Pi-Hydrogen bond: 3.018 Å; (ALA21)---(HCQ) Alkyl interaction: 3.885 Å; (MET11)---(HCQ) Alkyl interaction: 4.566 Å; (LEU30)---(HCQ) Pi-Alkyl interaction: 5.174 Å.  |  |
| Entry | Chloroquine (CQ) | Hydroxychloroquine (HCQ) |
|---|---|---|
| Physicochemical Properties, Lipophilicity and Drug-Likeness | ||
| Molecular weight (g/mol) | 319.87 | 335.87 |
| No. heavy atoms | 22 | 23 |
| No. arom. heavy atoms | 10 | 10 |
| Fraction Csp3 | 0.50 | 0.50 |
| No. rotatable bonds | 8 | 9 |
| No. H-bond acceptors | 2 | 3 |
| No. H-bond donors | 1 | 2 |
| Molar Refractivity | 97.41 | 98.57 |
| TPSA (Å2) | 28.16 | 48.39 |
| Consensus Log Po/w | 4.15 | 3.29 |
| Lipinski’s Rule | Yes | Yes |
| Bioavailability Score | 0.55 | 0.55 |
| PAINS | 0 alert | 0 alert |
| Pharmacokinetics | ||
| Gastrointestinal absorption | High | High |
| BBB permeant | Yes | Yes |
| P-gp substrate | No | No |
| CYP1A2 inhibitor | Yes | Yes |
| CYP2C19 inhibitor | No | No |
| CYP2C9 inhibitor | No | No |
| CYP2D6 inhibitor | Yes | Yes |
| CYP3A4 inhibitor | Yes | Yes |
| Log Kp (cm/s) | −4.96 | −5.81 |
 |  | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badraoui, R.; Adnan, M.; Bardakci, F.; Alreshidi, M.M. Chloroquine and Hydroxychloroquine Interact Differently with ACE2 Domains Reported to Bind with the Coronavirus Spike Protein: Mediation by ACE2 Polymorphism. Molecules 2021, 26, 673. https://doi.org/10.3390/molecules26030673
Badraoui R, Adnan M, Bardakci F, Alreshidi MM. Chloroquine and Hydroxychloroquine Interact Differently with ACE2 Domains Reported to Bind with the Coronavirus Spike Protein: Mediation by ACE2 Polymorphism. Molecules. 2021; 26(3):673. https://doi.org/10.3390/molecules26030673
Chicago/Turabian StyleBadraoui, Riadh, Mohd Adnan, Fevzi Bardakci, and Mousa M. Alreshidi. 2021. "Chloroquine and Hydroxychloroquine Interact Differently with ACE2 Domains Reported to Bind with the Coronavirus Spike Protein: Mediation by ACE2 Polymorphism" Molecules 26, no. 3: 673. https://doi.org/10.3390/molecules26030673