
Antiparasitic Potential of Chromatographic Fractions of Nephrolepis biserrata and Liquid Chromatography-Quadrupole Time-of-Flight-Mass Spectrometry Analysis

 , and
, and 
Abstract
:
1. Introduction
2. Results
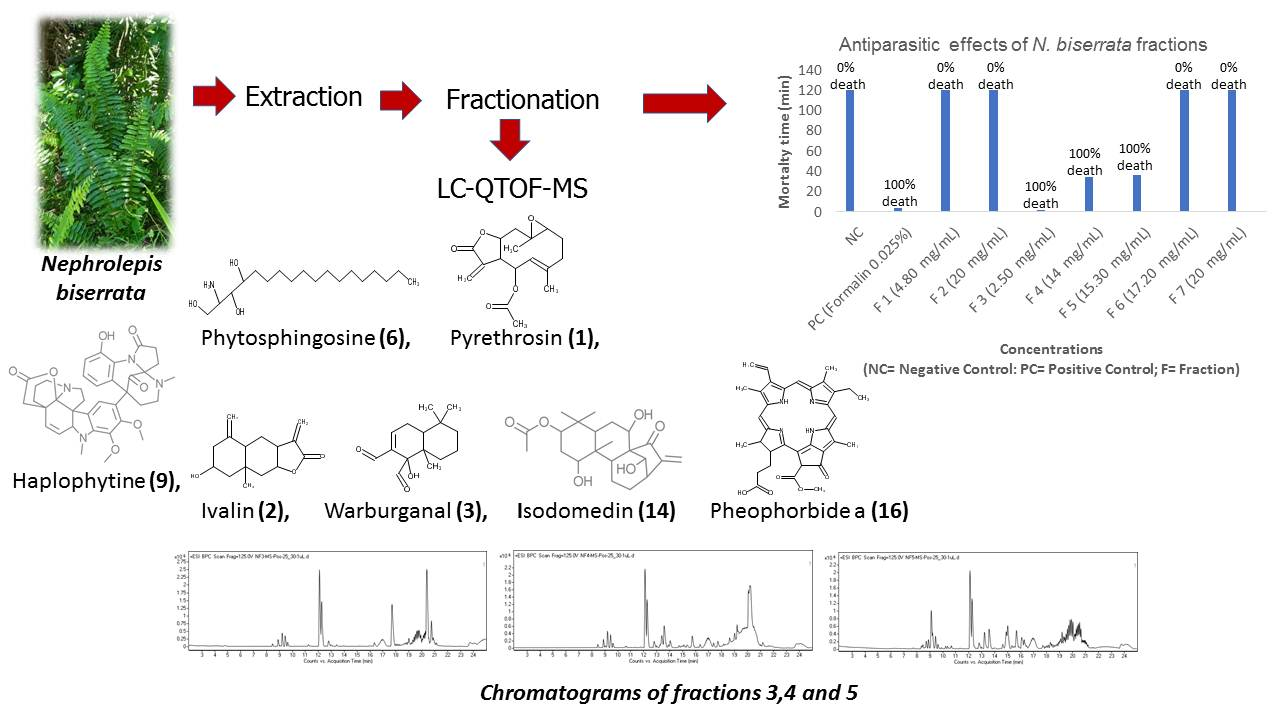
2.1. Antiparasitic Activity of the Chromatographic Fractions of N. biserrata Methanol Extract
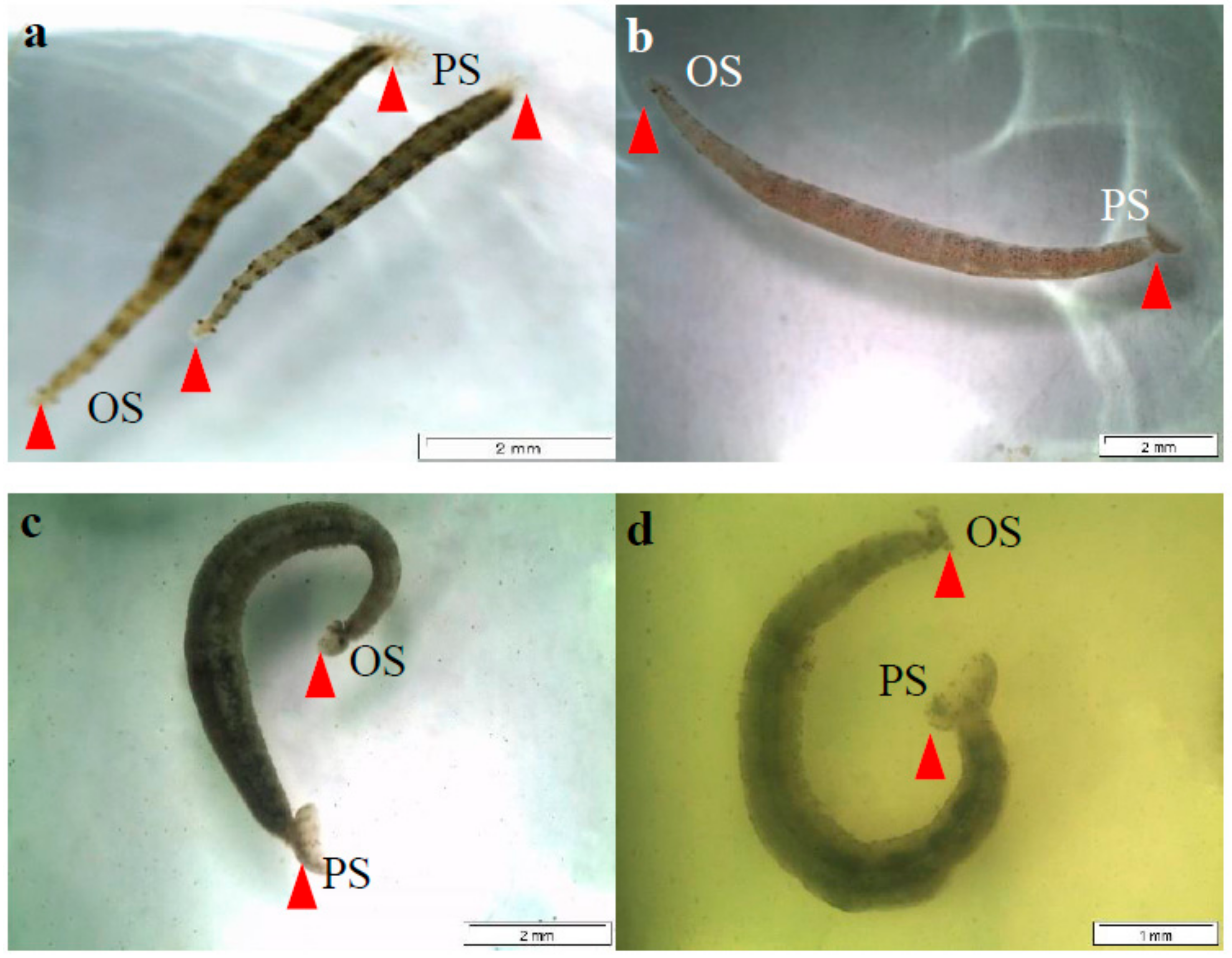
2.2. Behavioural Changes Induced by the Chromatographic Fractions of N. biserrata Methanol Extract
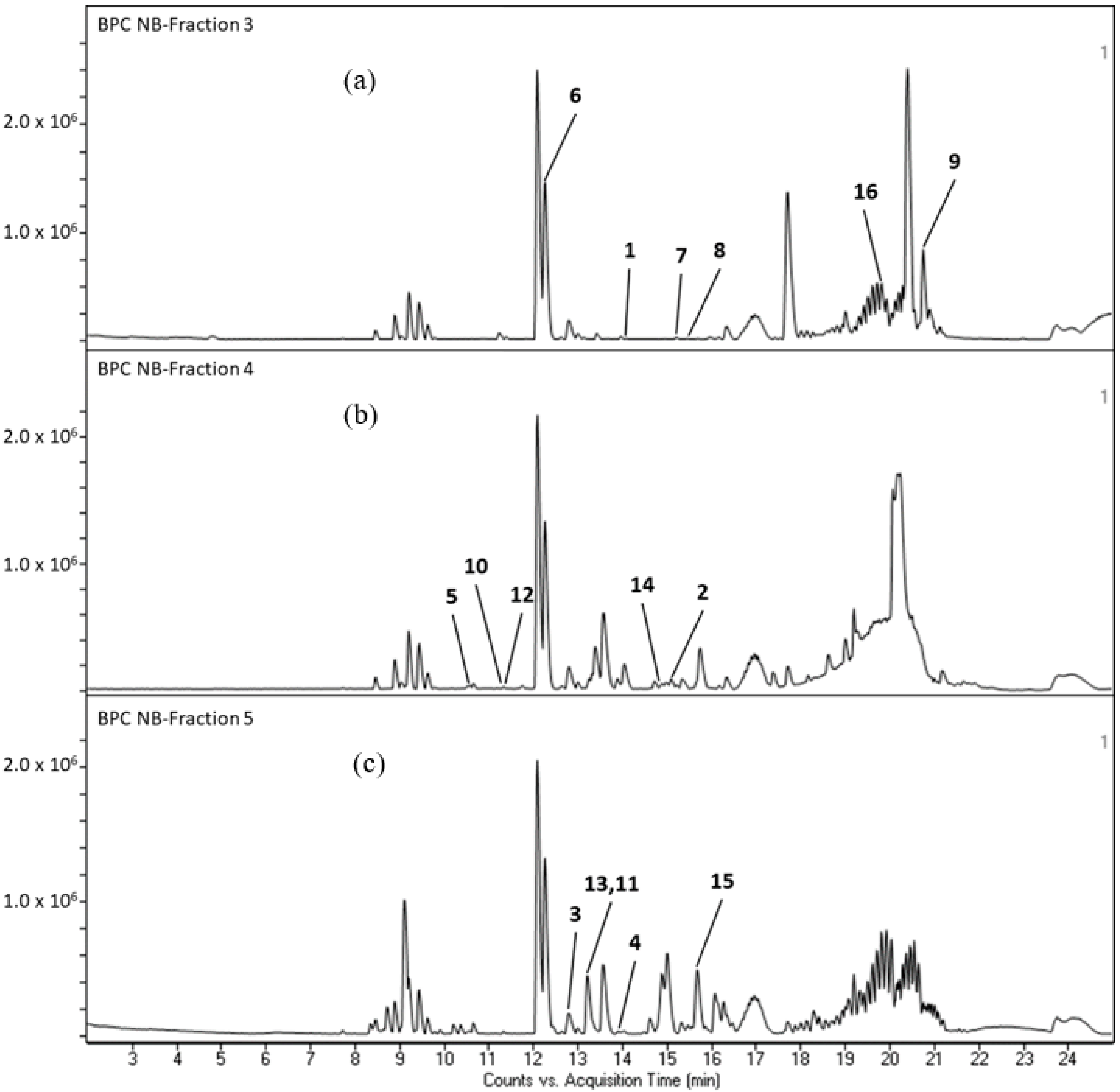
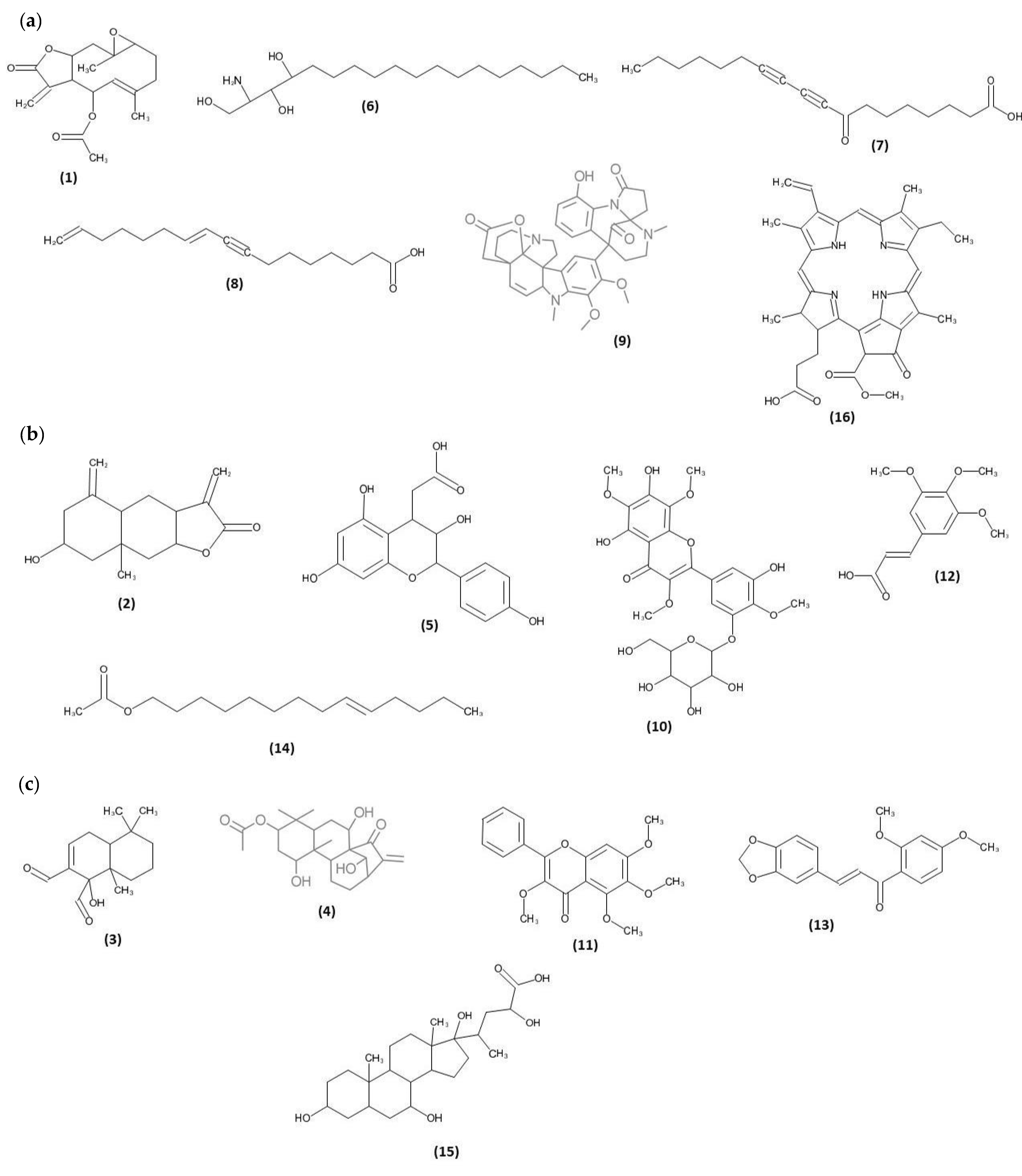
2.3. Phytochemical Compounds Detected in the Chromatographic Fractions of N. biserrata Methanol Extract
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Collection
4.3. Extraction and Fractionation
4.4. Antiparasitic Bioassay
4.5. Behavioural Observation
4.6. LC-QTOF MS Acquisition
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rosenberg, A.A. The price of lice. Nature 2008, 451, 23–24. [Google Scholar] [CrossRef] [PubMed]
- Othman, M.F.; Hashim, M.; Eim, Y.M.; Azmai, M.N.A.; Iksan, N.; Chong, H.G.; Merican, Z. Transforming the aquaculture industry in Malaysia. World Aquac. 2017, 48, 16–23. [Google Scholar]
- Ravi, R.; Yahaya, Z.S. Zeylanicobdella arugamensis, the marine leech from cultured crimson snapper (Lutjanus erythropterus), Jerejak Island, Penang, Malaysia. Asian Pac. J. Trop. Biomed. 2017, 7, 473–477. [Google Scholar] [CrossRef]
- Murwantoko, M.; Negoro, S.L.C.; Isnansetyo, A.; Zafran, Z. Short communication: Identification of marine leech and assessment of its prevalence and intensity on cultured hybrid groupers (Epinephelus sp.). Biodiversitas 2018, 19, 1798–1804. [Google Scholar] [CrossRef]
- Shinn, A.A.P.; Pratoomyot, J.; Bron, J.E.; Paladini, G.G.; Brooker, E.; Brooker, A.J. Economic impacts of aquatic parasites on global finfish production. Glob. Aquac. Advocate 2015, 2015, 82–84. [Google Scholar]
- Barber, I.; Hoare, D.; Krause, J. Effects of parasites on fish behaviour: A review and evolutionary perspective. Rev. Fish Biol. Fish. 2000, 10, 131–165. [Google Scholar] [CrossRef]
- Shah, M.D.; Venmathi Maran, B.A.; Haron, F.K.; Ching, F.F.; Shaleh, S.R.M.; Ransangan, J.; Yong, Y.-S.; Shapawi, R.; Ohtsuka, S. Antiparasitic potential of Nephrolepis biserrata methanol extract against the parasitic leech Zeylanicobdella arugamensis (Hirudinea) and LC-QTOF analysis. Sci. Rep. 2020, 10, 22091. [Google Scholar] [CrossRef]
- Khoa, T.N.D.; Mazelan, S.; Muda, S.; Shaharom-Harrison, F. Use of neem oil (Azadirachta indica) to control caligid copepod infestation on Asian seabass (Lates calcarifer). Aquac. Res. 2019, 50, 1885–1892. [Google Scholar] [CrossRef]
- De Oliveira Hashimoto, G.S.; Neto, F.M.; Ruiz, M.L.; Acchile, M.; Chagas, E.C.; Chaves, F.C.M.; Martins, M.L. Essential oils of Lippia sidoides and Mentha piperita against monogenean parasites and their influence on the hematology of Nile tilapia. Aquaculture 2016, 450, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Venmathi Maran, B.A.; Seng, L.T.; Ohtsuka, S.; Nagasawa, K. Records of Caligus (Crustacea: Copepoda: Caligidae) from marine fish cultured in floating cages in Malaysia with a redescription of the male of Caligus longipedis Bassett-Smith, 1898. Zool. Stud. 2009, 48, 797–807. [Google Scholar]
- Azmey, S.; Taruna, M.; Taha, H.; Arai, T. Prevalence and infestation intensity of a piscicolid leech, Zeylanicobdella arugamensis on cultured hybrid grouper in Brunei Darussalam. Vet. Parasitol. Reg. Stud. Rep. 2020, 20, 100398. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Lacierda, E.R.; Toledo, J.D.; Tan-Fermin, J.D.; Burreson, E.M. Marine leech (Zeylanicobdella arugamensis) infestation in cultured orange-spotted grouper, Epinephelus coioides. Aquaculture 2000, 185, 191–196. [Google Scholar] [CrossRef]
- Nagasawa, K.; Uyeno, D. Zeylanicobdella arugamensis (Hirudinea, Piscicolidae), a leech infesting brackish-water fishes, new to Japan. Biogeography 2009, 11, 125–130. [Google Scholar]
- Francis-Floyd, R. Use of formalin to control fish parasites. Coll. Vet. Med. Institue Food Agric. Sci. 1996, 77, 7–9. [Google Scholar]
- Fischer, M.H. The toxic effects of formaldehyde and formalin. J. Exp. Med. 1905, 6, 487–518. [Google Scholar] [CrossRef]
- MyBIS Nephrolepis biserrata | Nephrolepidaceae | Malaysia Biodiversity Information System. Available online: https://www.mybis.gov.my/sp/63552 (accessed on 24 September 2020).
- Kulip, J.; Fan, L.N.; Manshoor, N.; Julius, A.; Said, I.M.; Gisil, J.; Joseph, J.A.; Tukin, W.F. Medicinal plants in Maliau Basin, Sabah, Malaysia. J. Trop. Biol. Conserv. 2010, 6, 21–33. [Google Scholar]
- Shah, M.D.; Gnanaraj, C.; Haque, A.E.; Iqbal, M. Antioxidative and chemopreventive effects of Nephrolepis biserrata against carbon tetrachloride (CCl4)-induced oxidative stress and hepatic dysfunction in rats. Pharm. Biol. 2015, 53, 31–39. [Google Scholar] [CrossRef]
- Dantu, P.; Rani, D.; Khare, P. In vitro antibacterial and antifungal properties of aqueous and non-aqueous frond extracts of Psilotum nudum, Nephrolepis biserrata and Nephrolepis cordifolia. Indian J. Pharm. Sci. 2010, 72, 818. [Google Scholar] [CrossRef] [Green Version]
- Shariff, M.; Nagaraj, G.; Chua, F.H.; Wang, Y.G. The use of chemicals in aquaculture in Malaysia and Singapore. In Use of Chemicals in Aquaculture in Asia: Proceedings of the Meeting on the Use of Chemicals in Aquaculture in Asia 20–22 May 1996, Tigbauan, Iloilo, Philippines; Aquaculture Department, Southeast Asian Fisheries Development Center: Iloilo, Philippines, 2000; pp. 127–140. [Google Scholar]
- Valladão, G.M.R.; Gallani, S.U.; Pilarski, F. Phytotherapy as an alternative for treating fish disease. J. Vet. Pharmacol. Ther. 2015, 38, 417–428. [Google Scholar] [CrossRef]
- Shah, M.D.; Tani, K.; Venmathi Maran, B.A.; Yong, Y.S.; Fui Fui, C.; Shaleh, S.R.M.; Vairappan, C.S. High-resolution chemical profiling and antiparasitic potential of the tropical shrub Dillenia suffruticosa. Fish. Sci. 2020. [Google Scholar] [CrossRef]
- Kumar, A.; Raman, R.P.; Kumar, K.; Pandey, P.K.; Kumar, V.; Mohanty, S.; Kumar, S. Antiparasitic efficacy of piperine against Argulus spp. on Carassius auratus (Linn. 1758): In vitro and in vivo study. Parasitol. Res. 2012, 111, 2071–2076. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.F.; Zhu, B.; Wang, Y.; Lu, C.; Wang, G.X. In vivo evaluation of anthelmintic potential of medicinal plant extracts against Dactylogyrus intermedius (Monogenea) in goldfish (Carassius auratus). Parasitol. Res. 2011, 108, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wu, H.; Yu, X.; Zhang, X.; Lu, Y.; Fan, J.; Tang, L.; Wang, Z. A review of the phytochemistry and pharmacological activities of Magnoliae officinalis cortex. J. Ethnopharmacol. 2019, 236, 412–442. [Google Scholar] [CrossRef]
- Wang, R.; Deng, X.; Gao, Q.; Wu, X.; Han, L.; Gao, X.; Zhao, S.; Chen, W.; Zhou, R.; Li, Z.; et al. Sophora alopecuroides L.: An ethnopharmacological, phytochemical, and pharmacological review. J. Ethnopharmacol. 2020, 248, 112172. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.L.; Lu, C.; Hu, X.G.; Ling, F.; Wang, G.X. Antiprotozoal activity of medicinal plants against Ichthyophthirius multifiliis in goldfish (Carassius auratus). Parasitol. Res. 2012, 111, 1771–1778. [Google Scholar] [CrossRef]
- Pitten, F.A.; Kramer, A.; Herrmann, K.; Bremer, J.; Koch, S. Formaldehyde neurotoxicity in animal experiments. Pathol. Res. Pract. 2000, 196, 193–198. [Google Scholar] [CrossRef]
- Pavicic, T.; Wollenweber, U.; Farwick, M.; Korting, H.C. Anti-microbial and -inflammatory activity and efficacy of phytosphingosine: An in vitro and in vivo study addressing acne vulgaris. Int. J. Cosmet. Sci. 2007, 29, 181–190. [Google Scholar] [CrossRef]
- Nagahara, Y.; Shinomiya, T.; Kuroda, S.; Kaneko, N.; Nishio, R.; Ikekita, M. Phytosphingosine induced mitochondria-involved apoptosis. Cancer Sci. 2005, 96, 83–92. [Google Scholar] [CrossRef]
- Miranda, N.; Volpato, H.; da Silva Rodrigues, J.H.; Caetano, W.; Ueda-Nakamura, T.; de Oliveira Silva, S.; Nakamura, C.V. The photodynamic action of pheophorbide a induces cell death through oxidative stress in Leishmania amazonensis. J. Photochem. Photobiol. B Biol. 2017, 174, 342–354. [Google Scholar] [CrossRef]
- Ohta, S.; Ono, F.; Shiomi, Y.; Nakao, T.; Aozasa, O.; Nagate, T.; Kitamura, K.; Yamaguchi, S.; Nishi, M.; Miyata, H. Anti-herpes simplex virus substances produced by the marine green alga, Dunaliella primolecta. J. Appl. Phycol. 1998, 10, 349–356. [Google Scholar] [CrossRef]
- Ratnoglik, S.L.; Aoki, C.; Sudarmono, P.; Komoto, M.; Deng, L.; Shoji, I.; Fuchino, H.; Kawahara, N.; Hotta, H. Antiviral activity of extracts from Morinda citrifolia leaves and chlorophyll catabolites, pheophorbide a and pyropheophorbide a, against hepatitis C virus. Microbiol. Immunol. 2014, 58, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Lauritano, C.; Helland, K.; Riccio, G.; Andersen, J.H.; Ianora, A.; Hansen, E.H. Lysophosphatidylcholines and chlorophyll-derived molecules from the diatom Cylindrotheca closterium with anti-inflammatory activity. Mar. Drugs 2020, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saide, A.; Lauritano, C.; Ianora, A. Pheophorbide a: State of the Art. Mar. Drugs 2020, 18, 257. [Google Scholar] [CrossRef]
- Saxton, J.E. Chapter 18 Alkaloids of Haplophyton Cimicidum. In Alkaloids: Chemistry and Physiology; Academic Press: Cambridge, MA, USA, 1965; Volume 8, pp. 673–678. [Google Scholar]
- Schmidt, T.J.; Brun, R.; Willuhn, G.; Khalid, S.A. Anti-trypanosomal activity of Helenalin and some structurally related sesquiterpene lactones. Planta Med. 2002, 68, 750–751. [Google Scholar] [CrossRef]
- Liu, F.; Lin, S.; Zhang, C.; Ma, J.; Han, Z.; Jia, F.; Xie, W.; Li, X. The novel nature microtubule inhibitor ivalin induces G2/M arrest and apoptosis in human hepatocellular carcinoma SMMC-7721 cells in vitro. Medicina 2019, 55, 470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Kipanga, P.; Mai, A.H.; Dhondt, I.; Braeckman, B.P.; De Borggraeve, W.; Luyten, W. Bioassay-guided isolation of three antihelminthic compounds from Warburgia ugandensis Sprague subspecies ugandensis, and the mechanism of action of polygodial. Int. J. Parasitol. 2018, 48, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Kubo, I. Studies on Warburganal, Muzigadial and related compounds. Isr. J. Chem. 1977, 16, 28–31. [Google Scholar] [CrossRef]
- Kubo, I.; Miura, I.; Nakanishi, K.; Kamikawa, T.; Isobe, T.; Kubota, T. Structure of Isodomedin, a novel ent-kaurenoid diterpene. J. Chem. Soc. Chem. Commun. 1977, 555. [Google Scholar] [CrossRef]
- Srivastava, R.; Srivastava, P. Hepatotoxicity and the role of some herbal hepatoprotective plants in present scenario. Int. J. Dig. Dis. 2018, 4, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Shah, M.D.; Venmathi Maran, B.A.; Iqbal, M.; Ching, F.F.; Lal, M.T.M.; Othman, R.O.; Shapawi, R. Antiparasitic activity of the medicinal plant Dillenia suffruticosa against the marine leech Zeylanicobdella arugamensis (Hirudinea) and its phytochemical composition. Aquac. Res. 2020, 51, 215–221. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A technology platform for identifying knowns and unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Group | Death Time (min) Mean ± S.D. | Death Percentage |
|---|---|---|---|
| 1 | Negative Control | 0 00 ± 0.00 | 0 |
| 2 | Positive Control (Formalin 0.025%) | 3.90 ± 0.84 # | 100 |
| 3 | Fraction 1 (4.80 mg/mL) | 0 00 ± 0.00 * | 0 |
| 4 | Fraction 2 (20 mg/mL) | 0 00 ± 0.00 * | 0 |
| 5 | Fraction 3 (2.50 mg/mL) | 1.92 ± 0.59 # * ^ $ | 100 |
| 6 | Fraction 4 (14 mg/mL) | 34.57 ± 3.39 # * ^ $ & | 100 |
| 7 | Fraction 5 (15.30 mg/mL) | 36.82 ± 4.53 # * ^ $ & | 100 |
| 8 | Fraction 6 (17.20 mg/mL) | 0 00 ± 0.00 * & α ® | 00 |
| 9 | Fraction 7 (20 mg/mL) | 0 00 ± 0.00 * & β ® | 00 |
| Fractions | Retention Time (RT) | Mass to Charge Ratio, m/z | Formula | Mass Error, ppm | Matched Metabolites | Class |
|---|---|---|---|---|---|---|
| 1 | 13.144 | 203.1043 | C11 H16 O2 | −1.39 | 3-tert-Butyl-5-methylcatechol | Phenolic |
| 2 | 12.848 | 231.1377 | C15 H18 O2 | 0.30 | Isodehydrocostus lactone | Terpenoid |
| 13.112 | 203.1045 | C11 H16 O2 | −1.39 | 3-tert-Butyl-5-methylcatechol | Phenolic | |
| 13.443 | 311.1847 | C17 H26 O5 | −0.46 | methyl 8-[2-(2-formyl-vinyl)-3-hydroxy-5-oxo-cyclopentyl]-octanoate | Fatty acid methyl ester | |
| 16.533 | 280.1905 | C16 H25 N O3 | −0.37 | Serratine | Alkaloid | |
| 19.234 | 425.2905 | C24 H40 O6 | −1.51 | (3α,5β,6α,7α)-2,3,6,7-Tetrahydroxycholan-24-oic acid | Steroid | |
| 24.443 | 431.3522 | C28 H46 O3 | −0.03 | 1,25-Dihydroxy-ethylidene-19-norvitamin D3 | Steroid | |
| 3 | 12.220 | 318.3001 | C18 H39 N O3 | 0.93 | Phytosphingosine (6) | Sphingoid |
| 14.096 | 307.1541 | C17 H22 O5 | −0.38 | Pyrethrosin (1) | Terpenoid | |
| 15.081 | 291.1954 | C18 H26 O3 | −0.04 | 8-oxo-9,11-octadecadiynoic acid (7) | Oxo Fatty acid | |
| 15.343 | 263.2002 | C17 H26 O2 | 1.19 | 10,16-Heptadecadien-8-ynoic acid (8) | Fatty Acid | |
| 19.835 | 593.2766 | C35 H36 N4 O5 | −0.86 | Pheophorbide a (16) | Chlorophyll breakdown product | |
| 20.777 | 653.2969 | C37 H40 N4 O7 | 0.46 | Haplophytine (9) | Alkaloid | |
| 4 | 10.663 | 333.0965 | C17 H16 O7 | 1.05 | 3′-Deoxydryopteric acid (5) | Phenolic |
| 11.325 | 391.1018 | C19 H18 O9 | 1.34 | 5,2′,4′-Trihydroxy-3,7,8,5′-tetramethoxyflavone (10) | Flavonoid | |
| 11.404 | 239.0911 | C12 H14 O5 | 0.11 | 3,4,5-Trimethoxycinnamic acid (12) | Aromatic | |
| 14.927 | 277.2138 | C16 H30 O2 | 0.12 | 9-Tetradecenyl acetate (14) | Aliphatic acetate | |
| 15.120 | 249.1483 | C15 H20 O3 | −0.32 | Ivalin (2) | Terpenoid | |
| 5 | 12.796 | 251.1636 | C15 H22 O3 | 1.86 | Warburganal (3) | Terpenoid |
| 13.223 | 313.1070 | C18 H16 O5 | 0.75 | 3,4-Methylenedioxy-2′,4′-dimethoxychalcone (13) | Aromatic | |
| 13.245 | 343.1176 | C19 H18 O6 | 0.27 | 3,5,6,7-Tetramethoxyflavone (11) | Flavonoid | |
| 13.964 | 415.2085 | C22 H32 O6 | 1.05 | Isodomedin (4) | Terpenoid | |
| 15.651 | 447.2719 | C24 H40 O6 | −1.27 | 3,7,17,23-Tetrahydroxycholan-24-oic acid (15) | Steroid | |
| 6 | 8.965 | 465.1018 | C21 H20 O12 | 1.83 | Herbacetin 3-glucoside | Flavonoid |
| 10.052 | 611.1391 | C30 H26 O14 | 1.04 | Gallocatechin-(4alpha- > 8)-epigallocatechin | Flavonoid | |
| 12.306 | 290.269 | C16 H32 O3 | 0.26 | 14-hydroxy-hexadecanoic acid | Fatty acid | |
| 13.230 | 343.117 | C19 H18 O6 | 1.78 | 3,5,6,7-Tetramethoxyflavone | Flavonoid | |
| 13.983 | 415.2086 | C22 H32 O6 | 1.55 | Isodomedin | Terpenoid | |
| 21.127 | 391.2845 | C24 H38 O4 | −0.89 | 3,7-Dihydroxychol-22-en-24-oic acid | Steroid | |
| 7 | 2.053 | 328.1393 | C15 H18 O7 | −0.86 | Picrotin | Terpenoid |
| 7.065 | 163.0386 | C9 H6 O3 | 1.15 | 3-Hydroxycoumarin | Aromatic | |
| 7.309 | 355.1016 | C16 H18 O9 | 1.62 | Chlorogenic Acid | Phenolic | |
| 8.945 | 465.1020 | C21 H20 O12 | 1.89 | Herbacetin 3-glucoside | Flavonoid | |
| 9.008 | 291.0975 | C14 H14 N2 O5 | −0.40 | N2-Malonyl-D-tryptophan | Aromatic | |
| 9.333 | 303.0500 | C15 H10 O7 | −1.53 | Melanoxetin | Flavonoid | |
| 9.333 | 449.1077 | C21 H20 O11 | 0.25 | Kaempferol 5-glucoside | Flavonoid | |
| 10.034 | 404.1341 | C20 H18 O8 | −0.71 | Glucosyloxyanthraquinone | Aromatic | |
| 12.296 | 290.2687 | C16 H32 O3 | 1.23 | 14-hydroxy-hexadecanoic acid | Fatty acid | |
| 13.994 | 432.2381 | C24 H30 O6 | −0.39 | Magnoshinin | Aromatic | |
| 17.923 | 506.2541 | C30 H32 O6 | −1.45 | Rubraflavone D | Phenolic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, M.D.; Tani, K.; Yong, Y.S.; Ching, F.F.; Shaleh, S.R.M.; Vairappan, C.S.; Venmathi Maran, B.A. Antiparasitic Potential of Chromatographic Fractions of Nephrolepis biserrata and Liquid Chromatography-Quadrupole Time-of-Flight-Mass Spectrometry Analysis. Molecules 2021, 26, 499. https://doi.org/10.3390/molecules26020499
Shah MD, Tani K, Yong YS, Ching FF, Shaleh SRM, Vairappan CS, Venmathi Maran BA. Antiparasitic Potential of Chromatographic Fractions of Nephrolepis biserrata and Liquid Chromatography-Quadrupole Time-of-Flight-Mass Spectrometry Analysis. Molecules. 2021; 26(2):499. https://doi.org/10.3390/molecules26020499
Chicago/Turabian StyleShah, Muhammad Dawood, Kazuki Tani, Yoong Soon Yong, Fui Fui Ching, Sitti Raehanah Muhamad Shaleh, Charles S. Vairappan, and Balu Alagar Venmathi Maran. 2021. "Antiparasitic Potential of Chromatographic Fractions of Nephrolepis biserrata and Liquid Chromatography-Quadrupole Time-of-Flight-Mass Spectrometry Analysis" Molecules 26, no. 2: 499. https://doi.org/10.3390/molecules26020499