
Carnosine to Combat Novel Coronavirus (nCoV): Molecular Docking and Modeling to Cocrystallized Host Angiotensin-Converting Enzyme 2 (ACE2) and Viral Spike Protein


Abstract
:1. Introduction
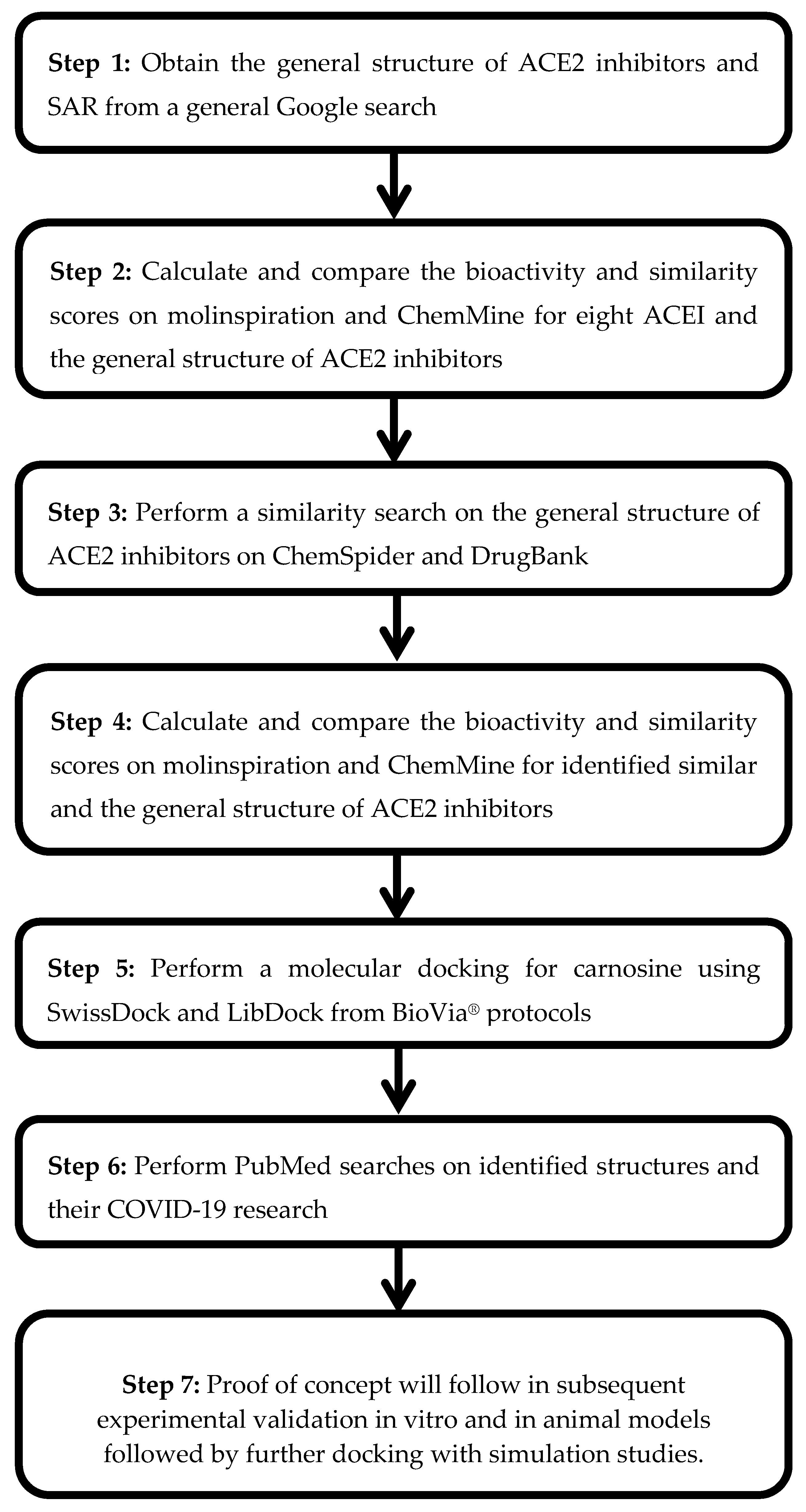
2. Aims
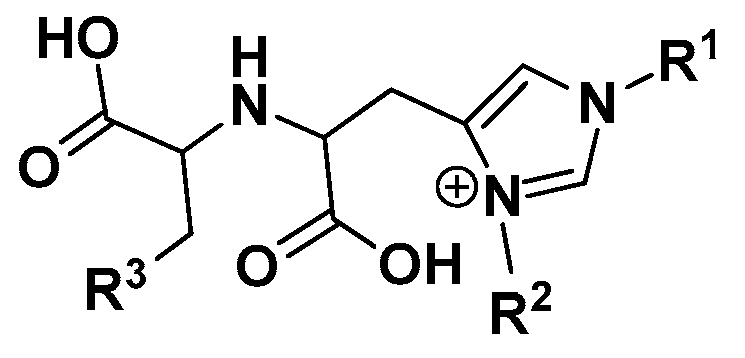
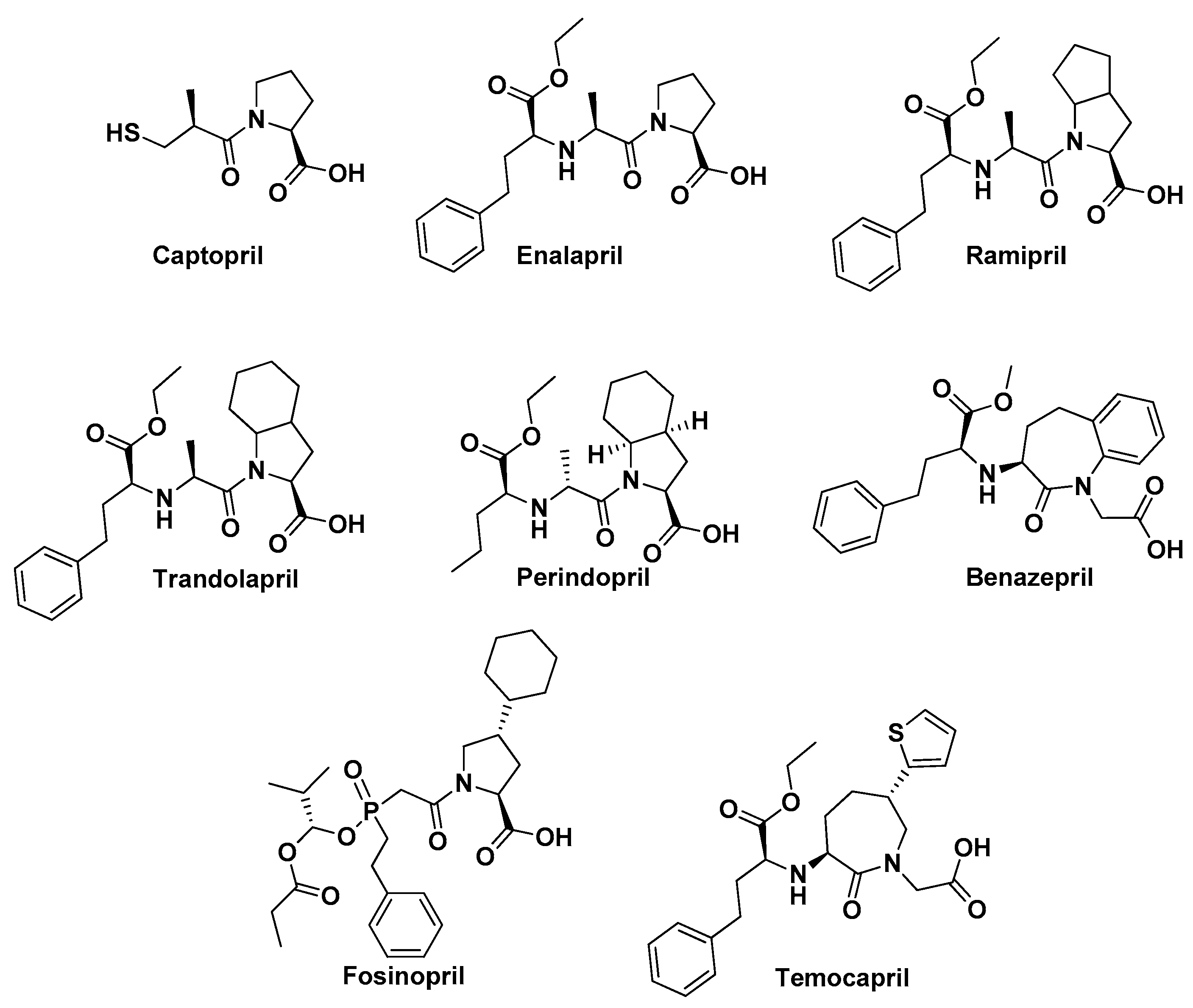
3. Methods
Statistical Analysis
4. Results and Discussion
4.1. ACE Inhibitors and COVID-19
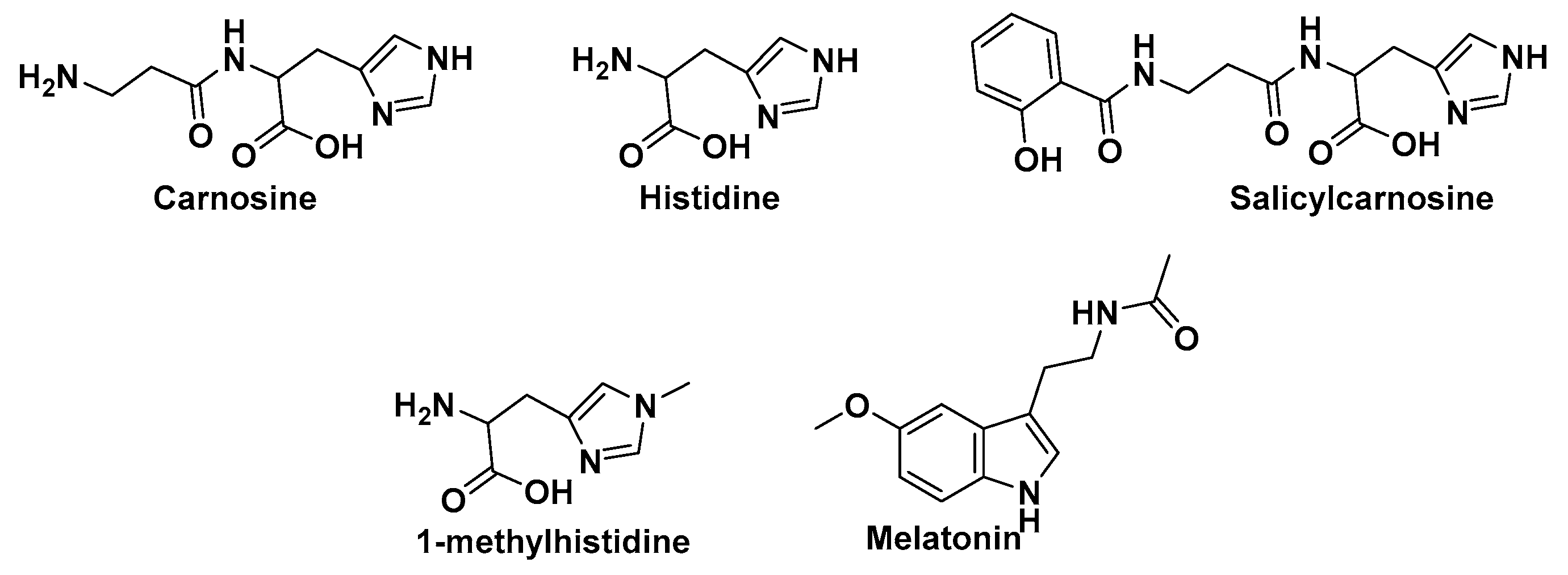
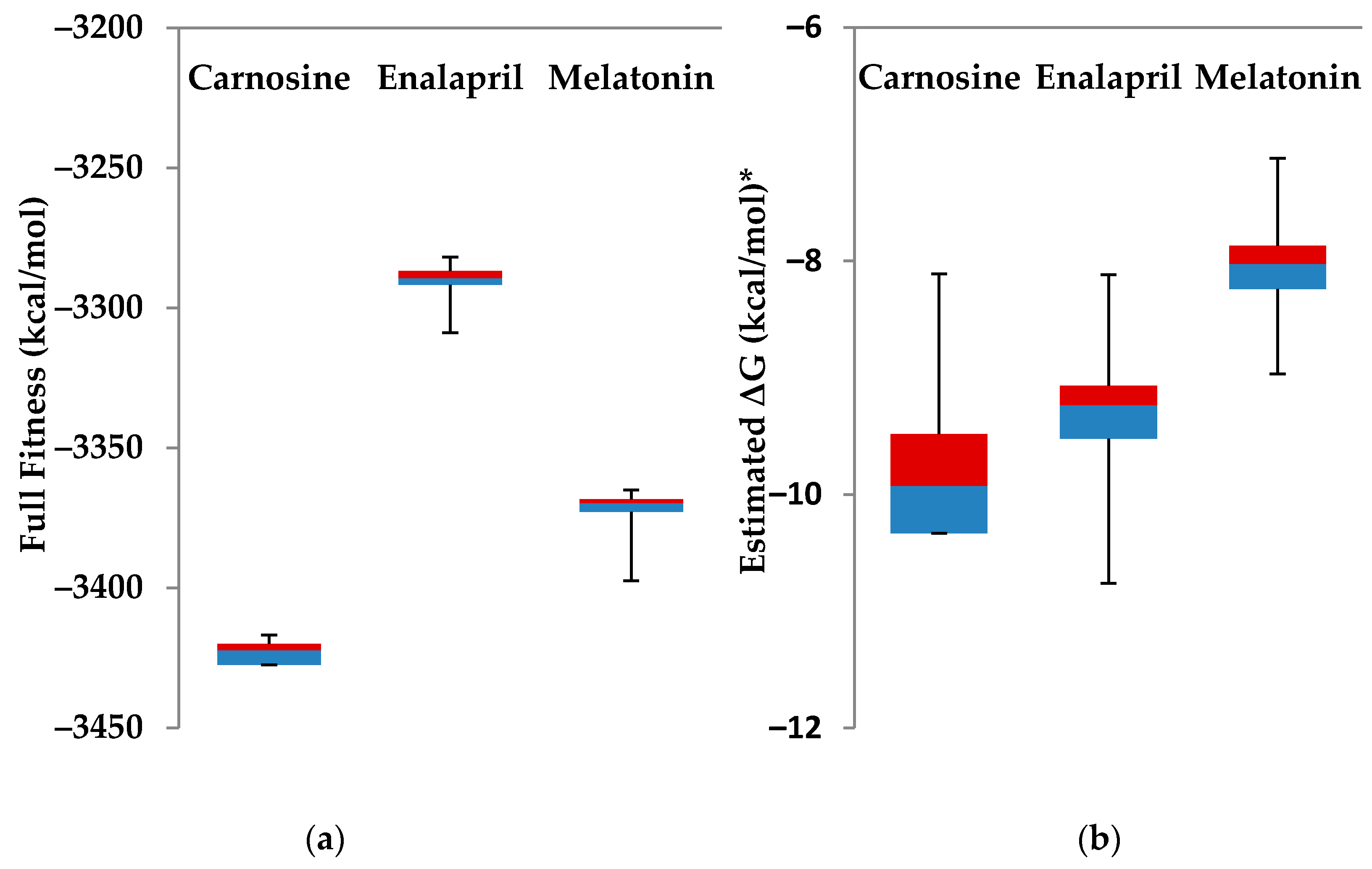
4.2. Carnosine Preliminary Molecular Docking
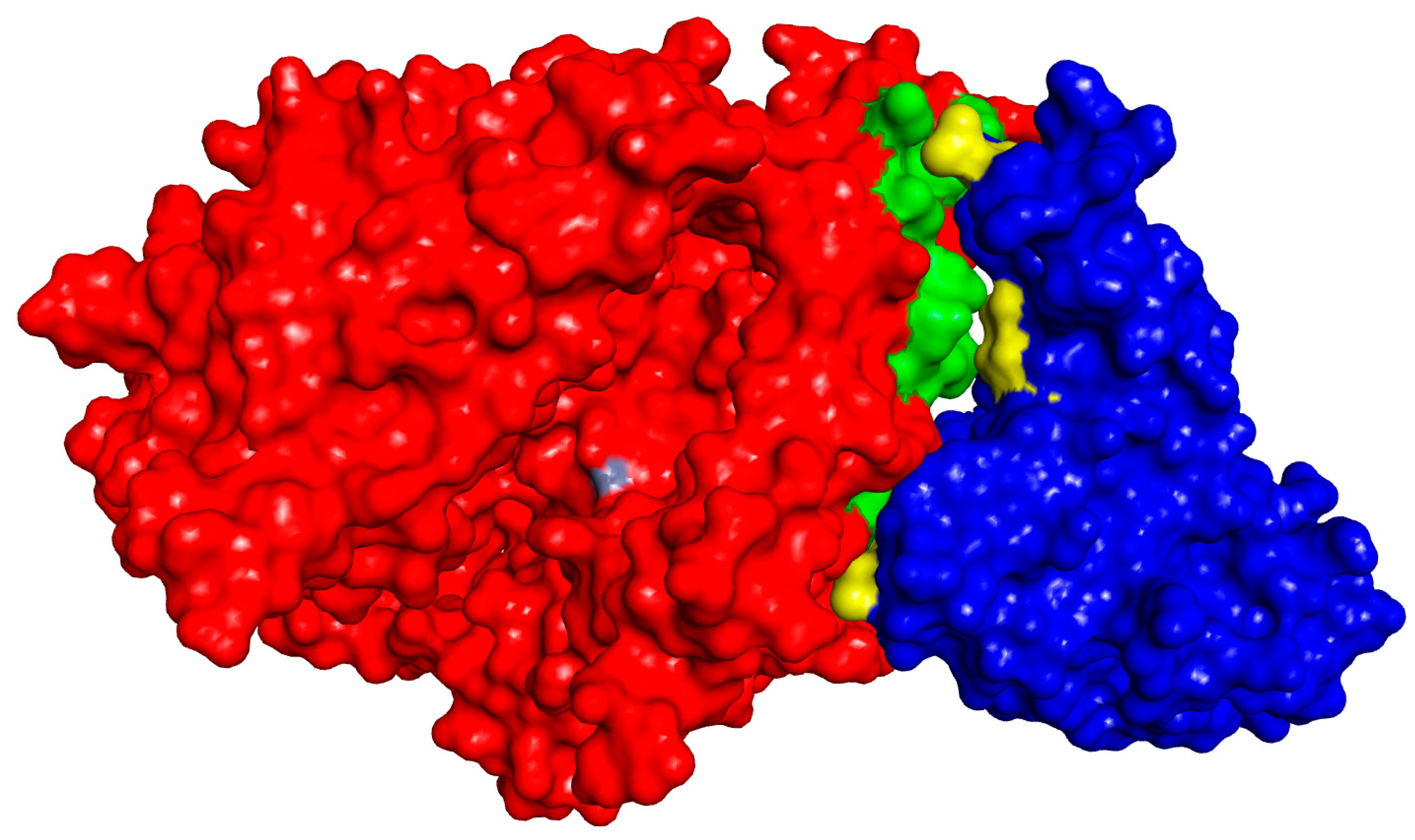
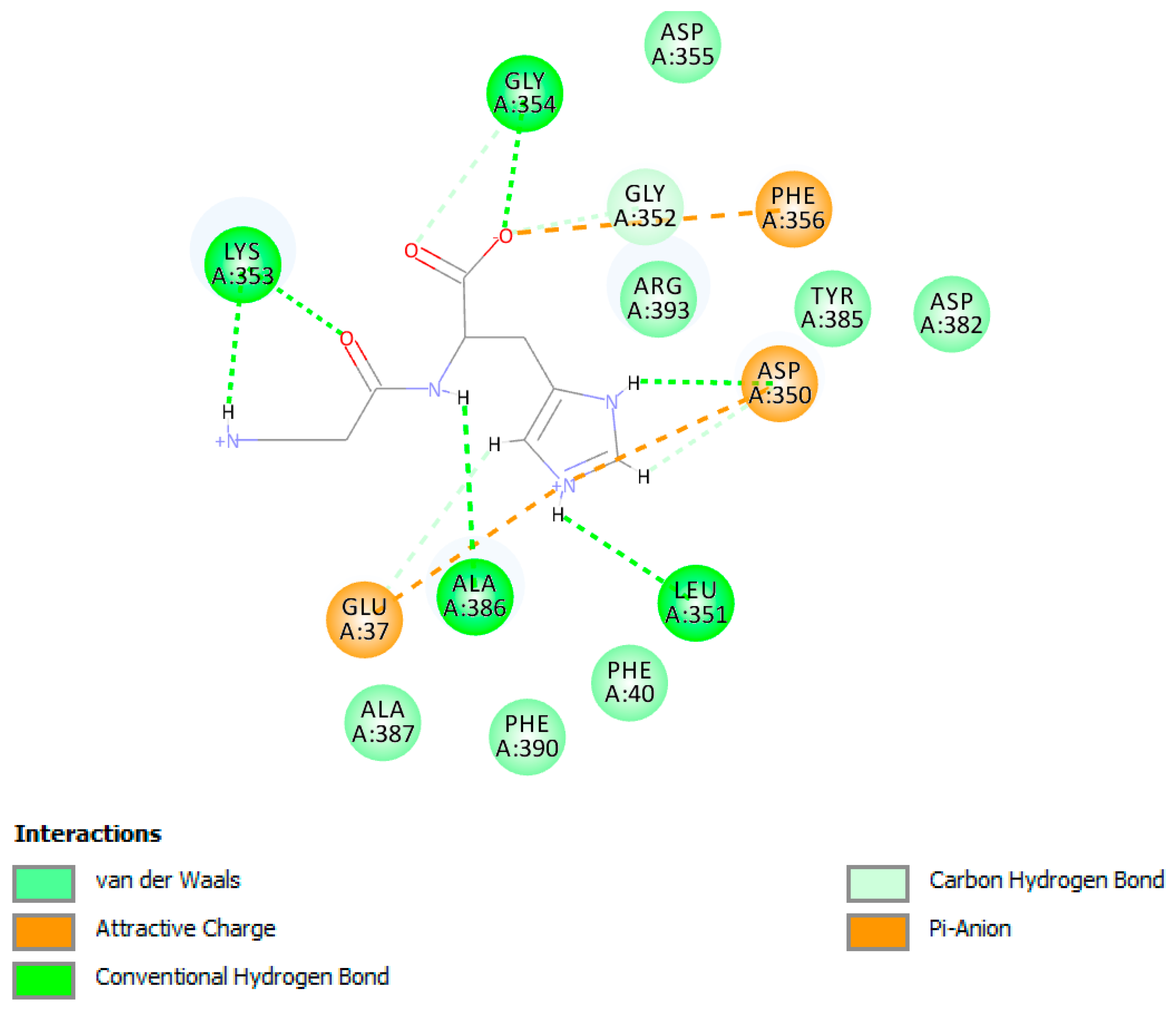
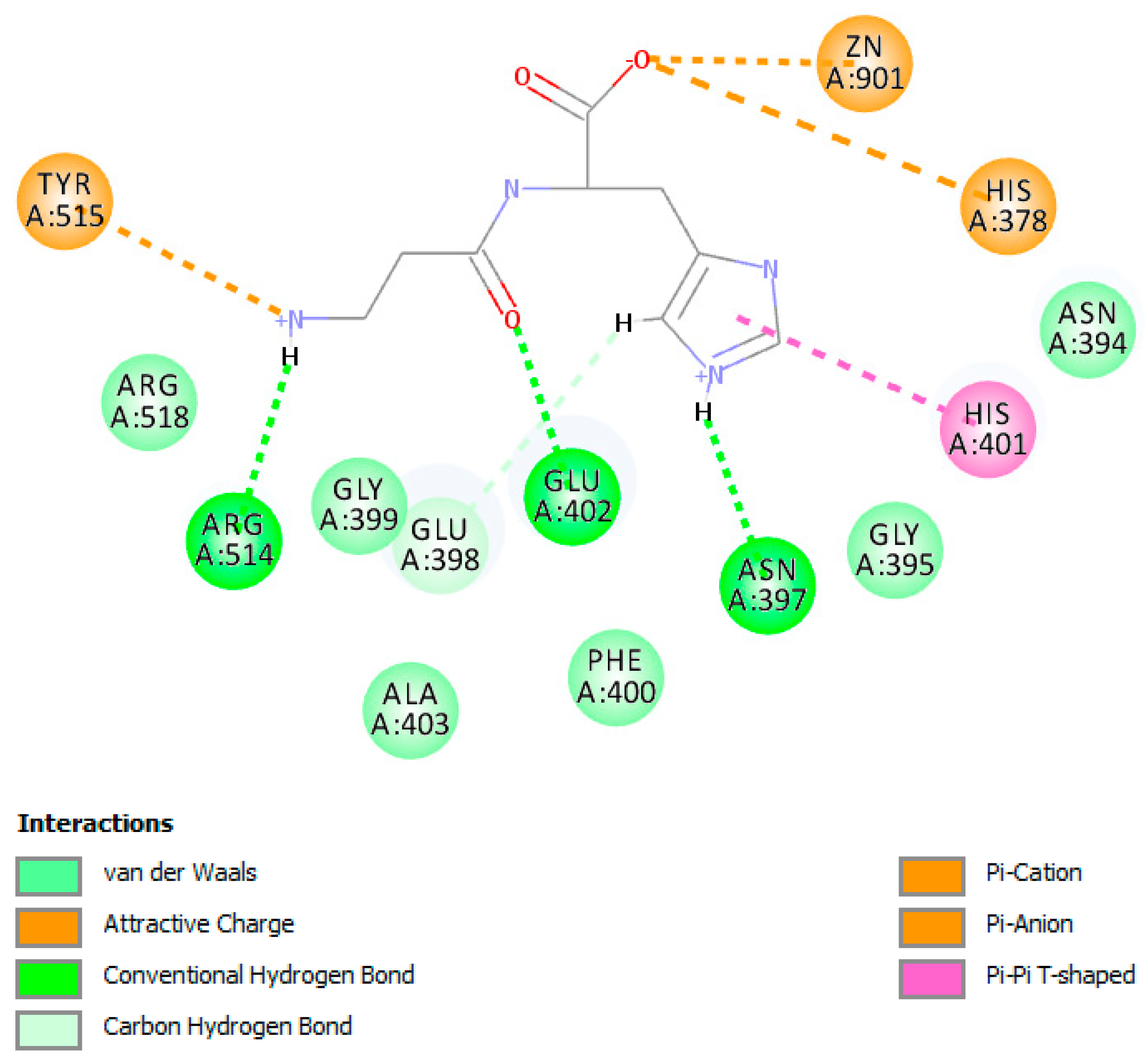
4.3. Detailed Molecular Docking and Modeling
4.4. Literature Evaluation
4.5. Bioavailability and Physiological Role of Carnosine and Salicylcarnosine
4.6. Limitations
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stepensky, D. Prediction of Drug Disposition on the Basis of its Chemical Structure. Clin. Pharm. 2013, 52, 415–431. [Google Scholar] [CrossRef] [PubMed]
- Al-Taweel, D.; Al-Haqan, A.; Bajis, D.; Al-Bader, J.; Al-Taweel, A.M.; Al-Awadhi, A.; Al-Awadhi, F. Multidisciplinary academic perspectives during the COVID-19 pandemic. Int. J. Health Plann. Manag. 2020. [Google Scholar] [CrossRef]
- Amawi, H.; Abu Deiab, G.I.; Aljabali, A.A.A.; Dua, K.; Tambuwala, M.M. COVID-19 pandemic: An overview of epidemiology, pathogenesis, diagnostics and potential vaccines and therapeutics. Ther. Deliv. 2020, 11, 245–268. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Rolling Updates on Coronavirus Disease (COVID-19). Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/events-as-they-happen (accessed on 25 July 2020).
- Worldometer. COVID-19 Coronavirus Pandemic. Available online: https://www.worldometers.info/coronavirus/ (accessed on 21 September 2020).
- Sahebnasagh, A.; Avan, R.; Saghafi, F.; Mojtahedzadeh, M.; Sadremomtaz, A.; Arasteh, O.; Tanzifi, A.; Faramarzi, F.; Negarandeh, R.; Safdari, M.; et al. Pharmacological treatments of COVID-19. Pharmacol. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Travanty, E.A.; Oko, L.; Edeen, K.; Berglund, A.; Wang, J.; Ito, Y.; Holmes, K.V.; Mason, R.J. Innate immune response of human alveolar type II cells infected with severe acute respiratory syndrome-coronavirus. Am. J. Respir. Cell Mol. Biol. 2013, 48, 742–748. [Google Scholar] [CrossRef] [Green Version]
- Trindade, G.G.; Caxito, S.M.C.; Xavier, A.R.W.O.; Xavier, M.A.S.; BrandÃo, F. COVID-19: Therapeutic approaches description and discussion. An. Acad. Bras. Cienc. 2020, 92, e20200466. [Google Scholar] [CrossRef]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [Green Version]
- Benton, D.J.; Wrobel, A.G.; Xu, P.; Roustan, C.; Martin, S.R.; Rosenthal, P.B.; Skehel, J.J.; Gamblin, S.J. Receptor binding and priming of the spike protein of SARS-CoV-2 for membrane fusion. Nature 2020. [Google Scholar] [CrossRef]
- Peter, E.K.; Schug, A. The Inhibitory Effect of a Coronavirus Spike Protein Fragment with ACE2. Biophys. J. 2020. [Google Scholar] [CrossRef]
- Pollard, C.A.; Morran, M.P.; Nestor-Kalinoski, A.L. The COVID-19 Pandemic: A Global Health Crisis. Physiol. Genom. 2020. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Abhinand, C.S.; Nair, A.S.; Krishnamurthy, A.; Oommen, O.V.; Sudhakaran, P.R. Potential protease inhibitors and their combinations to block SARS-CoV-2. J. Biomol. Struct. Dyn. 2020, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.E.; Baldiris, R.; Vivas-Reyes, R. Design of Angiotensin-converting Enzyme 2 (ACE2) Inhibitors by Virtual Lead Optimization and Screening. J. Chin. Chem. Soc. 2012, 59, 1394–1400. [Google Scholar] [CrossRef] [PubMed]
- Fara, D.C.; Oprea, T.I. Section III: Cheminformatics—Basics: Molecular Similarity (or Diversity). University of New Mexico. Available online: http://datascience.unm.edu/biomed505/Course/Cheminformatics/basic/similarity_diversity/similarity_diversity.htm (accessed on 25 July 2020).
- Lee, J.K. Statistical Bioinformatics: A Guide for Life and Biomedical Science Researchers; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar]
- Baldi, P.; Benz, R.W. BLASTing small molecules--statistics and extreme statistics of chemical similarity scores. Bioinformatics 2008, 24, i357–i365. [Google Scholar] [CrossRef] [Green Version]
- Dales, N.; Gould, A.; Brown, J.; Calderwood, E.; Guan, B.; Minor, C.; Gavin, J.; Hales, P.; Kaushik, V.; Stewart, M.; et al. Substrate-Based Design of the First Class of Angiotensin-Converting Enzyme-Related Carboxypeptidase (ACE2) Inhibitors. J. Am. Chem. Soc. 2002, 124, 11852. [Google Scholar] [CrossRef]
- Jarrahpour, A.; Fathi, J.; Mostafa, M.; Hadda, T.; Sheikh, J.; Chohan, Z.H.; Ali, P. Petra, Osiris and Molinspiration (POM) Together as a Successful Support in Drug Design: Antibacterial Activity and Biopharmaceutical Characterization of Some Azo Schiff Bases. Med. Chem. Res. 2012, 21, 1984–1990. [Google Scholar] [CrossRef]
- Backman, T.W.H.; Cao, Y.; Girke, T. ChemMine tools: An online service for analyzing and clustering small molecules. Nucleic Acids Res. 2011, 39 (Suppl. 2), W486–W491. [Google Scholar] [CrossRef]
- Pence, H.E.; Williams, A. ChemSpider: An Online Chemical Information Resource. J. Chem. Educ. 2010, 87, 1123–1124. [Google Scholar] [CrossRef]
- Wishart, D.S.; Wu, A. Using DrugBank for in silico drug exploration and discovery. Curr. Protoc. Bioinform. 2016, 54, 14.4.1–14.4.31. [Google Scholar] [CrossRef]
- Kong, Q.; Wu, Y.; Gu, Y.; Lv, Q.; Qi, F.; Gong, S.; Chen, X. Analysis of the molecular mechanism of Pudilan (PDL) treatment for COVID-19 by network pharmacology tools. Biomed. Pharmacother. 2020, 128, 110316. [Google Scholar] [CrossRef]
- Feitosa, E.L.; Tiago Dos, S.S.; Júnior, F.; De O Nery Neto, J.A.; Matos, L.F.L.; De S Moura, M.H.; Rosales, T.O.; De Freitas, G.B.L. COVID-19: Rational discovery of the therapeutic potential of Melatonin as a SARS-CoV-2 main Protease Inhibitor. Int. J. Med. Sci. 2020, 17, 2133–2146. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fosbøl, E.L.; Butt, J.H.; Østergaard, L.; Andersson, C.; Selmer, C.; Kragholm, K.; Schou, M.; Phelps, M.; Gislason, G.H.; Gerds, T.A.; et al. Association of Angiotensin-Converting Enzyme Inhibitor or Angiotensin Receptor Blocker Use with COVID-19 Diagnosis and Mortality. JAMA 2020, 324, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Menon, K.; Mousa, A.; de Courten, B. Effects of supplementation with carnosine and other histidine-containing dipeptides on chronic disease risk factors and outcomes: Protocol for a systematic review of randomised controlled trials. BMJ Open 2018, 8, e020623. [Google Scholar] [CrossRef]
- Beltrán-García, J.; Osca-Verdegal, R.; Pallardó, F.V.; Ferreres, J.; Rodríguez, M.; Mulet, S.; Sanchis-Gomar, F.; Carbonell, N.; García-Giménez, J.L. Oxidative Stress and Inflammation in COVID-19-Associated Sepsis: The Potential Role of Anti-Oxidant Therapy in Avoiding Disease Progression. Antioxidants 2020, 9, 936. [Google Scholar] [CrossRef] [PubMed]
- Scuto, M.; Salinaro, A.T.; Modafferi, S.; Polimeni, A.; Pfeffer, T.; Weigand, T.; Calabrese, V.; Schmitt, C.P.; Peters, V. Carnosine Activates Cellular Stress Response in Podocytes and Reduces Glycative and Lipoperoxidative Stress. Biomedicines 2020, 8, 177. [Google Scholar] [CrossRef] [PubMed]
- Ooi, T.C.; Chan, K.M.; Sharif, R. Zinc L-Carnosine Protects CCD-18co Cells from L-Buthionine Sulfoximine-Induced Oxidative Stress via the Induction of Metallothionein and Superoxide Dismutase 1 Expression. Biol. Trace Elem Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, O.I.; Stvolinsky, S.L.; Migulin, V.A.; Andreeva, L.A.; Nagaev, I.Y.; Lopacheva, O.M.; Kulichenkova, K.N.; Lopachev, A.V.; Trubitsina, I.E.; Fedorova, T.N. A new derivative of acetylsalicylic acid and carnosine: Synthesis, physical and chemical properties, biological activity. Daru 2020, 28, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Bermúdez, M.L.; Skelton, M.R.; Genter, M.B. Intranasal carnosine attenuates transcriptomic alterations and improves mitochondrial function in the Thy1-aSyn mouse model of Parkinson’s disease. Mol. Genet Metab. 2018, 125, 305–313. [Google Scholar] [CrossRef]
- Johansen, M.D.; Irving, A.; Montagutelli, X.; Tate, M.D.; Rudloff, I.; Nold, M.F.; Hansbro, N.G.; Kim, R.Y.; Donovan, C.; Liu, G.; et al. Animal and translational models of SARS-CoV-2 infection and COVID-19. Mucosal Immunol. 2020, 13, 877–891. [Google Scholar] [CrossRef]
- Khelfaoui, H.; Harkati, D.; Saleh, B.A. Molecular docking, molecular dynamics simulations and reactivity, studies on approved drugs library targeting ACE2 and SARS-CoV-2 binding with ACE2. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chikhale, R.V.; Gurav, S.S.; Patil, R.B.; Sinha, S.K.; Prasad, S.K.; Shakya, A.; Shrivastava, S.K.; Gurav, N.S.; Prasad, R.S. Sars-cov-2 host entry and replication inhibitors from Indian ginseng: An in-silico approach. J. Biomol. Struct. Dyn. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jindal, C.; Kumar, S.; Sharma, S.; Choi, Y.M.; Efird, J.T. The Prevention and Management of COVID-19: Seeking a Practical and Timely Solution. Int. J. Environ. Res. Public Health 2020, 17, 3986. [Google Scholar] [CrossRef] [PubMed]
- Lopachev, A.V.; Kazanskaya, R.B.; Khutorova, A.V.; Fedorova, T.N. An overview of the pathogenic mechanisms involved in severe cases of COVID-19 infection, and the proposal of salicyl-carnosine as a potential drug for its treatment. Eur. J. Pharmacol. 2020, 886, 173457. [Google Scholar] [CrossRef]
- Hipkiss, A.R. COVID-19 and Senotherapeutics: Any Role for the Naturally-occurring Dipeptide Carnosine? Aging Dis. 2020, 11, 737–741. [Google Scholar] [CrossRef]
- Rothan, H.A.; Abdulrahman, A.Y.; Khazali, A.S.; Nor Rashid, N.; Chong, T.T.; Yusof, R. Carnosine exhibits significant antiviral activity against Dengue and Zika virus. J. Pept. Sci. 2019, 25, e3196. [Google Scholar] [CrossRef]
- Xu, T.; Wang, C.; Zhang, R.; Xu, M.; Liu, B.; Wei, D.; Wang, G.; Tian, S. Carnosine markedly ameliorates H9N2 swine influenza virus-induced acute lung injury. J. Gen Virol. 2015, 96, 2939–2950. [Google Scholar] [CrossRef]
- Vistoli, G.; Carini, M.; Aldini, G. Transforming dietary peptides in promising lead compounds: The case of bioavailable carnosine analogs. Amino Acids 2012, 43, 111–126. [Google Scholar] [CrossRef]
- Soto, M.; di Zerega, G.; Rodgers, K.E. Countermeasure and therapeutic: A(1-7) to treat acute respiratory distress syndrome due to COVID-19 infection. J. Renin Angiotensin Aldosterone Syst. 2020, 21. [Google Scholar] [CrossRef]
- Huh, T.; Larouche-Lebel, E.; Loughran, K.A.; Oyama, M.A. Effect of angiotensin receptor blockers and angiotensin-converting enzyme 2 on plasma equilibrium angiotensin peptide concentrations in cats with heart disease. J. Vet. Intern. Med. 2020. [Google Scholar] [CrossRef]
- Saponaro, F.; Rutigliano, G.; Sestito, S.; Bandini, L.; Storti, B.; Bizzarri, R.; Zucchi, R. ACE2 in the Era of SARS-CoV-2: Controversies and Novel Perspectives. Front. Mol. Biosci. 2020, 7, 588618. [Google Scholar] [CrossRef] [PubMed]
- Gul, R.; Kim, U.; Alfadda, A.A. Renin-angiotensin system at the interface of COVID-19 infection. Eur. J. Pharmacol. 2020, 173656. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.I.; Kawahara, M. Carnosine and Lung Disease. Curr. Med. Chem. 2020, 27, 1714–1725. [Google Scholar] [CrossRef] [PubMed]
- Sale, C.; Saunders, B.; Harris, R.C. Effect of beta-alanine supplementation on muscle carnosine concentrations and exercise performance. Amino Acids 2010, 39, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.N.; Greaney, A.I.; Hilton, S.K.; Crawford, K.H.D.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; Veesler, D.; Bloom, J.D. Deep mutational scanning of SARS-CoV-2 receptor binding domain reveals constraints on folding and ACE2 binding. Cell 2020, 182, 1295–1310. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Boorla, V.S.; Maranas, C.D. Computational biophysical characterization of the SARS-CoV-2 spike protein binding with the ACE2 receptor and implications for infectivity. Comput. Struct. Biotechnol. J. 2020, 18, 2573–2582. [Google Scholar] [CrossRef]
- Chan, K.K.; Dorosky, D.; Sharma, P.; Abbasi, S.A.; Dye, P.M.; Kranz, D.M.; Herbert, A.S.; Procko, E. Engineering human ACE2 to optimize binding to the spike protein of SARS coronavirus 2. Science 2020, 369, 1261–1265. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | 1 * | 2 | 3 | 4 | 5 | 6 | Total Difference ** |
|---|---|---|---|---|---|---|---|
| General scaffold | 0.46 | 0.47 | −0.15 | −1.25 | 0.52 | 0.58 | 0 |
| Captopril | −0.14 | −0.08 | −0.98 | −0.55 | 0.97 | 0.50 | 3.21 |
| Enalapril | 0.36 | 0.16 | −0.30 | −0.08 | 0.70 | 0.18 | 2.31 |
| Ramipril | 0.36 | 0.08 | −0.36 | −0.12 | 0.78 | 0.23 | 2.44 |
| Trandolapril | 0.36 | 0.05 | −0.44 | −0.16 | 0.76 | 0.17 | 2.55 |
| Perindopril | 0.36 | 0.02 | −0.52 | −0.23 | 0.83 | 0.20 | 2.63 |
| Benazepril | 0.22 | 0.09 | −0.35 | 0.07 | 0.43 | 0.10 | 2.71 |
| Fosinopril | 0.44 | 0.07 | −0.31 | −0.11 | 1.03 | 0.41 | 2.40 |
| Temocapril | 0.10 | −0.13 | −0.45 | −0.21 | 0.49 | 0.03 | 2.88 |
| Carnosine | 0.61 | 0.48 | −0.06 | −1.2 | 0.65 | 0.73 | 0.58 |
| Salicyl-carnosine | 0.61 | 0.26 | 0.08 | −0.58 | 0.63 | 0.49 | 1.46 |
| Drug | AP Tanimoto | MCS Tanimoto | MCS Size | MCS Min | MCS Max |
|---|---|---|---|---|---|
| Scaffold | - | - | - | - | - |
| Captopril | 0.128 | 0.429 | 9 | 0.643 | 0.563 |
| Enalapril | 0.169 | 0.344 | 11 | 0.688 | 0.407 |
| Ramipril | 0.147 | 0.314 | 11 | 0.688 | 0.367 |
| Trandolapril | 0.138 | 0.306 | 11 | 0.688 | 0.355 |
| Perindopril | 0.138 | 0.355 | 11 | 0.688 | 0.423 |
| Benzapril | 0.114 | 0.306 | 11 | 0.688 | 0.355 |
| Fosinopril | 0.051 | 0.196 | 9 | 0,.563 | 0.231 |
| Temocapril | 0.110 | 0.297 | 11 | 0.688 | 0.344 |
| Carnosine | 0.364 | 0.684 | 13 | 0.813 | 0.813 |
| Salicyl-carnosine | 0.214 | 0.464 | 13 | 0.813 | 0.520 |
Sample Availability: Samples of the compounds are available from the authors.. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saadah, L.M.; Deiab, G.I.A.; Al-Balas, Q.; Basheti, I.A. Carnosine to Combat Novel Coronavirus (nCoV): Molecular Docking and Modeling to Cocrystallized Host Angiotensin-Converting Enzyme 2 (ACE2) and Viral Spike Protein. Molecules 2020, 25, 5605. https://doi.org/10.3390/molecules25235605
Saadah LM, Deiab GIA, Al-Balas Q, Basheti IA. Carnosine to Combat Novel Coronavirus (nCoV): Molecular Docking and Modeling to Cocrystallized Host Angiotensin-Converting Enzyme 2 (ACE2) and Viral Spike Protein. Molecules. 2020; 25(23):5605. https://doi.org/10.3390/molecules25235605
Chicago/Turabian StyleSaadah, Loai M., Ghina’a I. Abu Deiab, Qosay Al-Balas, and Iman A. Basheti. 2020. "Carnosine to Combat Novel Coronavirus (nCoV): Molecular Docking and Modeling to Cocrystallized Host Angiotensin-Converting Enzyme 2 (ACE2) and Viral Spike Protein" Molecules 25, no. 23: 5605. https://doi.org/10.3390/molecules25235605