Effects of Length and Loop Composition on Structural Diversity and Similarity of (G3TG3NmG3TG3) G-Quadruplexes


Abstract
:1. Introduction
2. Results
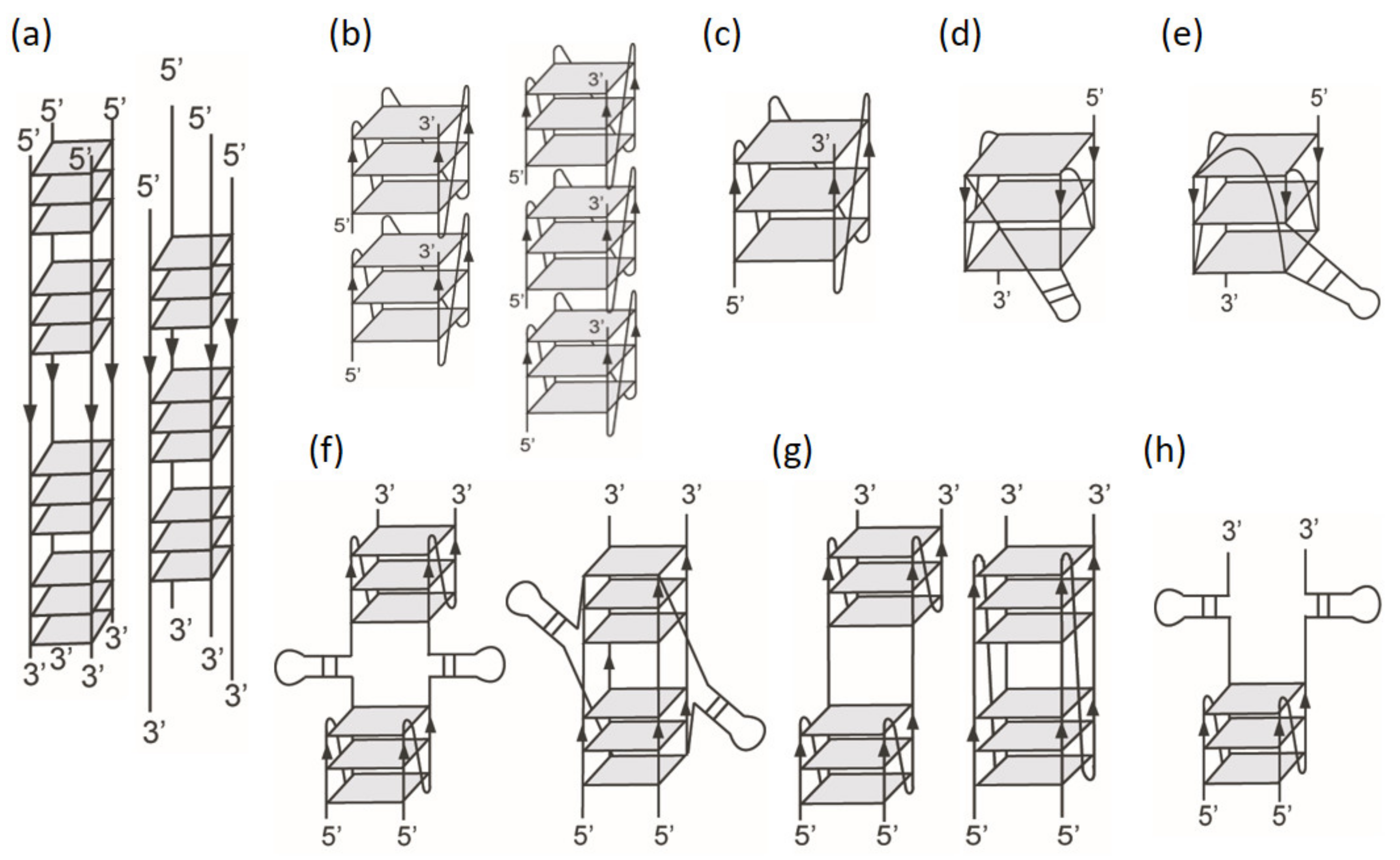
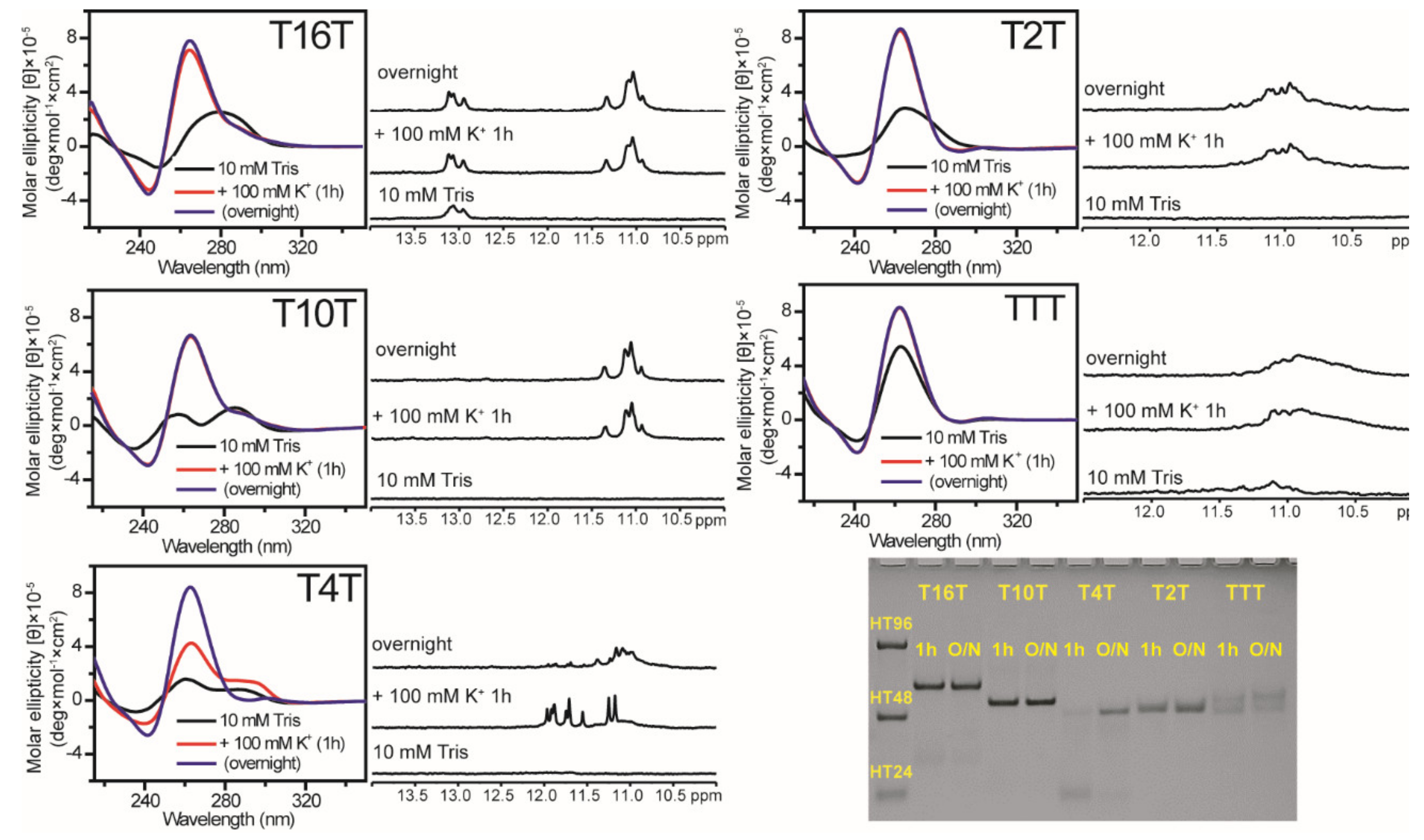
2.1. Effect of Loop Length on G4 Formation of (G3TG3NmG3TG3) Sequences
2.2. Effect of Base Composition in the Middle Loop on G4 Formation of (G3TG3N4G3TG3) Sequences
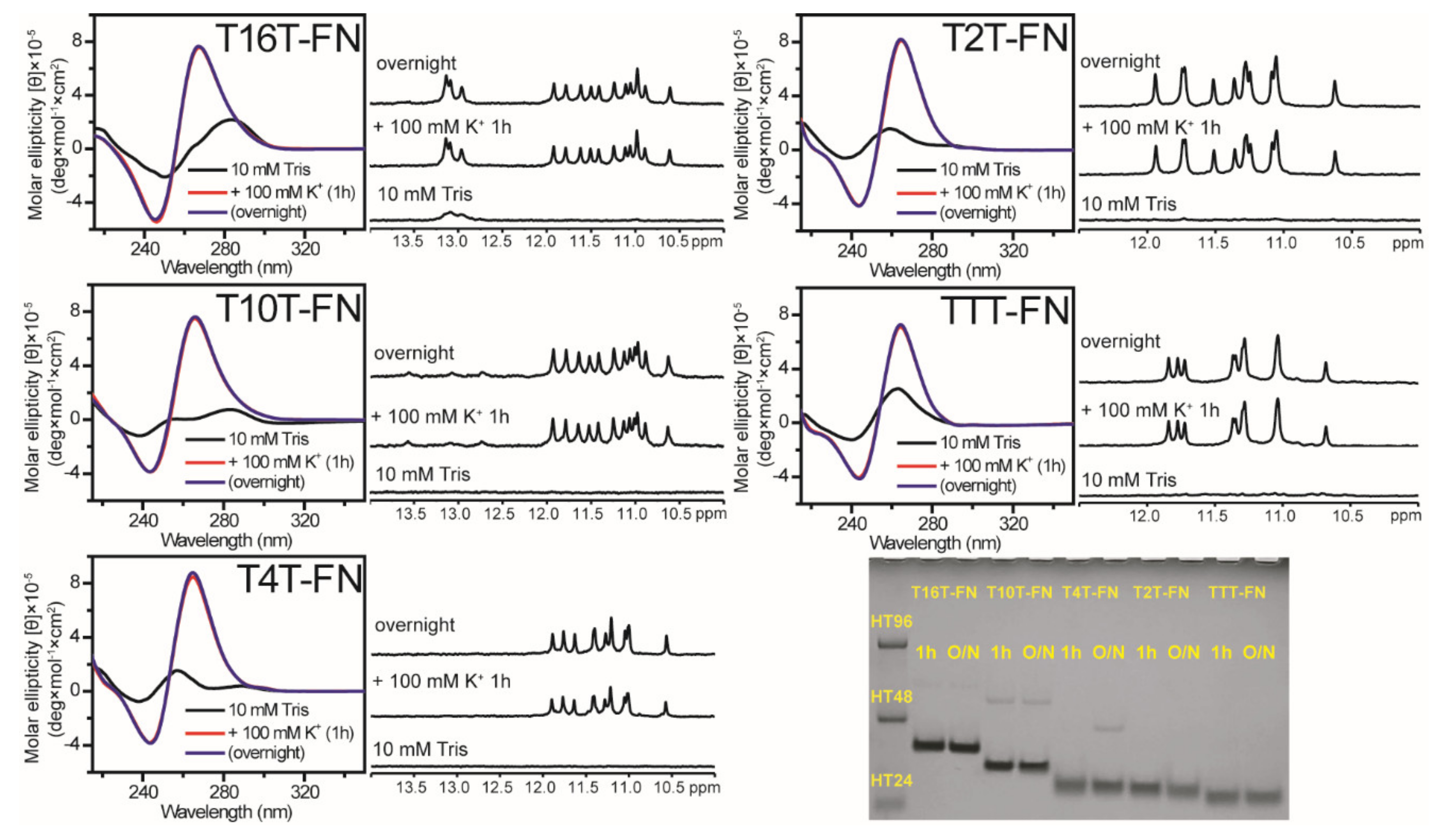
2.3. Effect of Flanking Nucleotides on G4 Formation of (G3TG3NmG3TG3)-FN Sequences
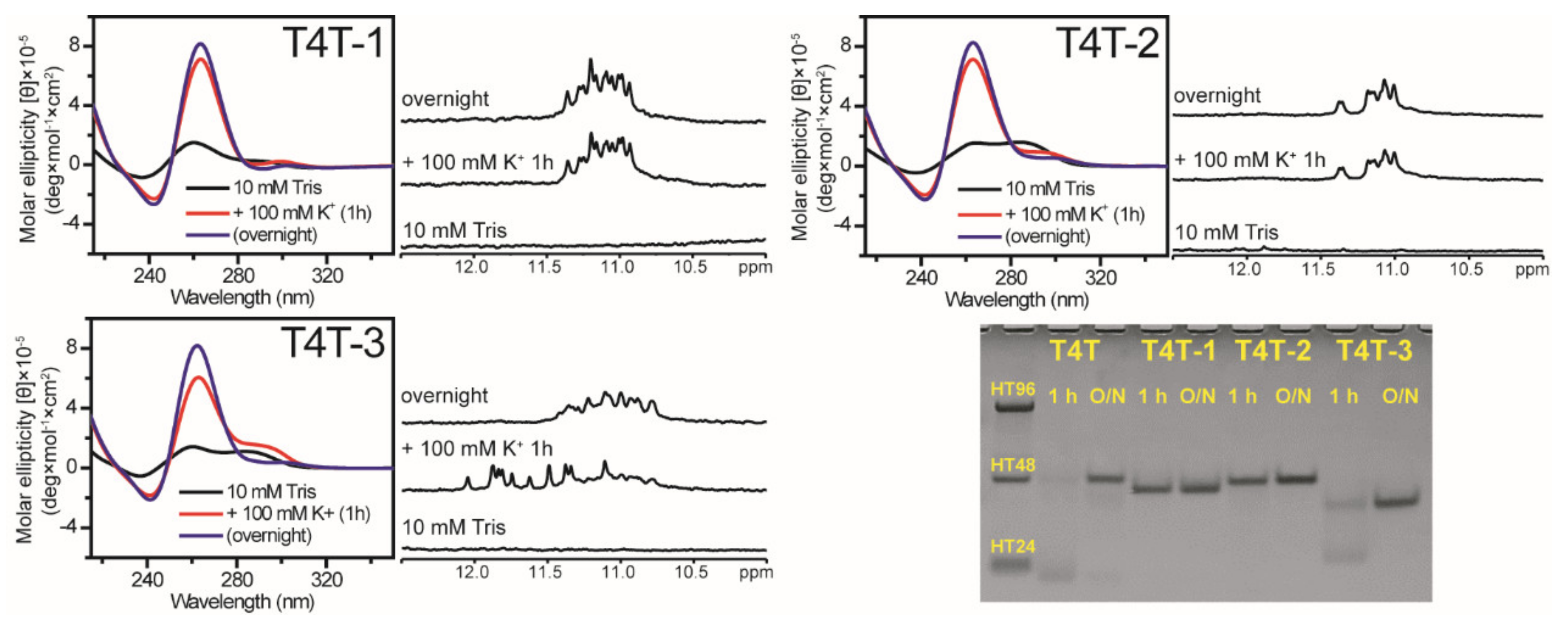
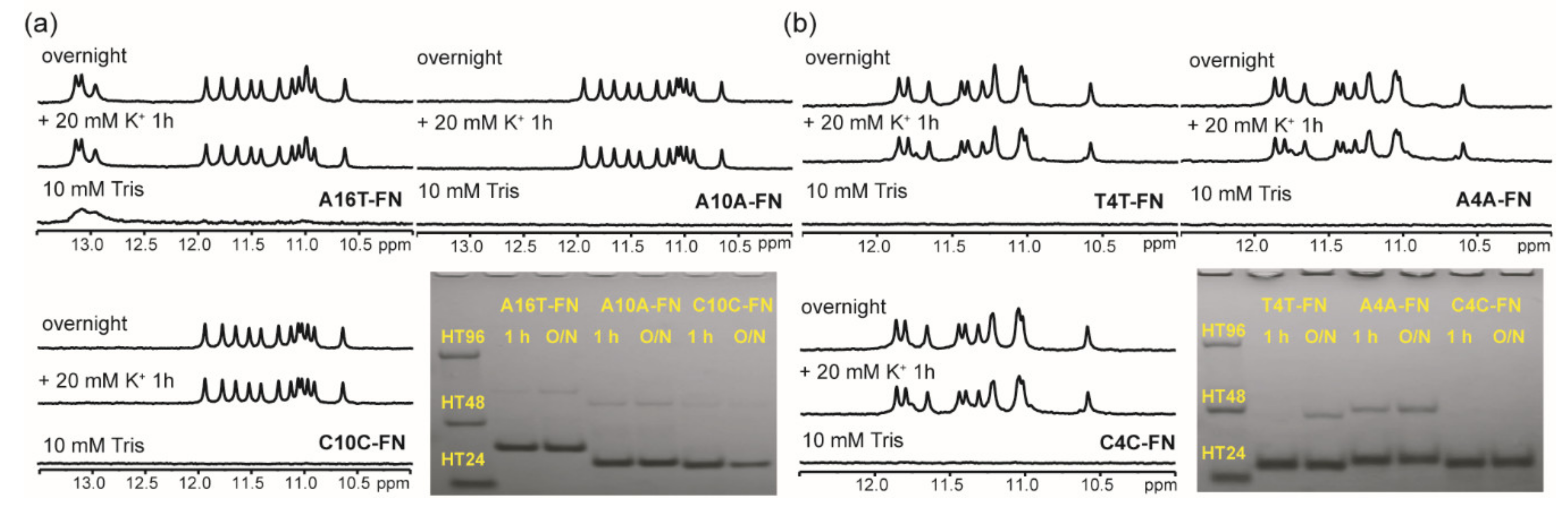
2.4. Effect of a Single Base Difference in Loop 1 and 3 on G4 Formation of (G3HG3NmG3HG3)-FN Sequences
2.5. Effect on G4 Formation of Native G-Rich Sequences without Flanking Nucleotides
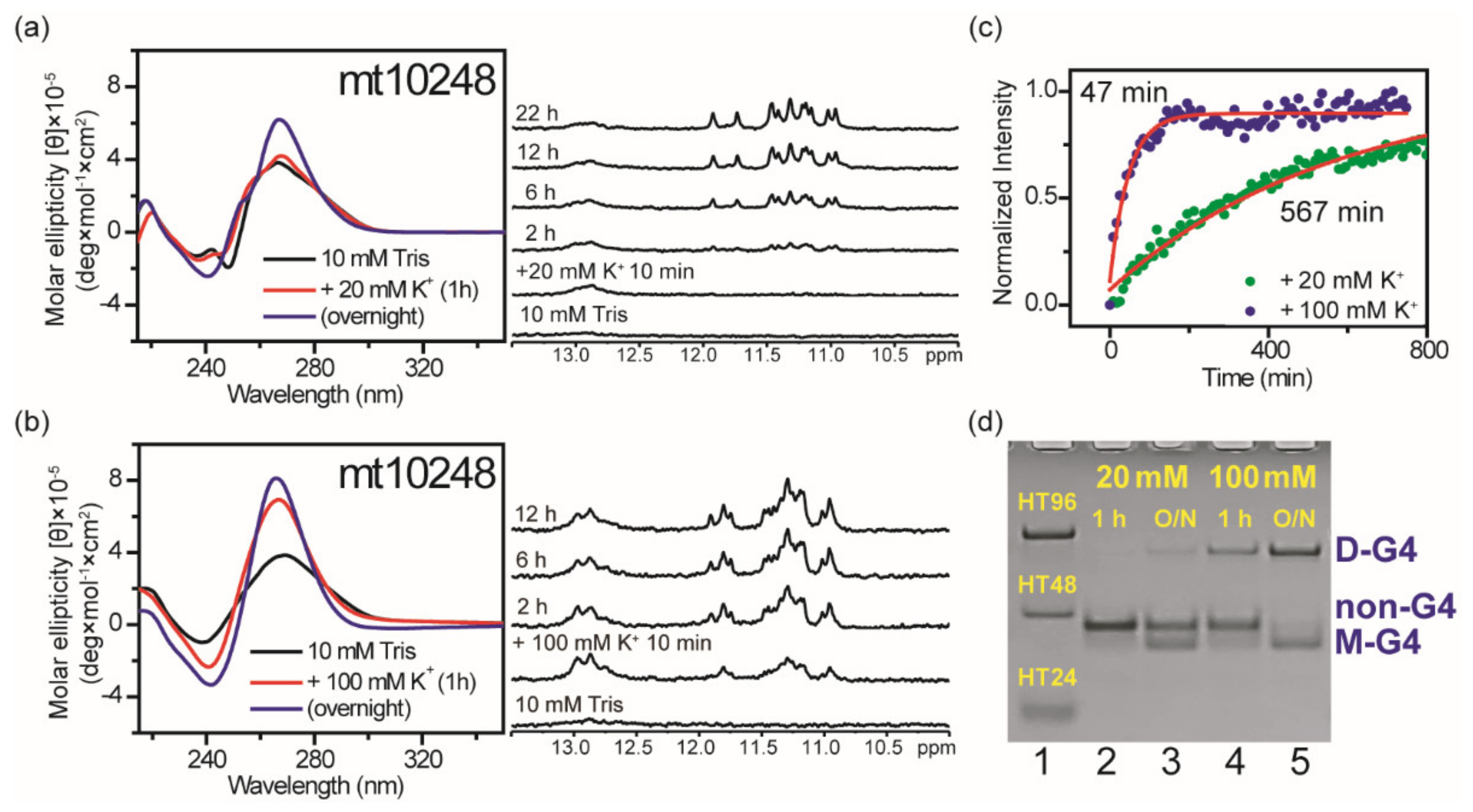
2.6. Effect of Flanking Nucleotides on G4 Formation of mt10251-FN (mt10248)
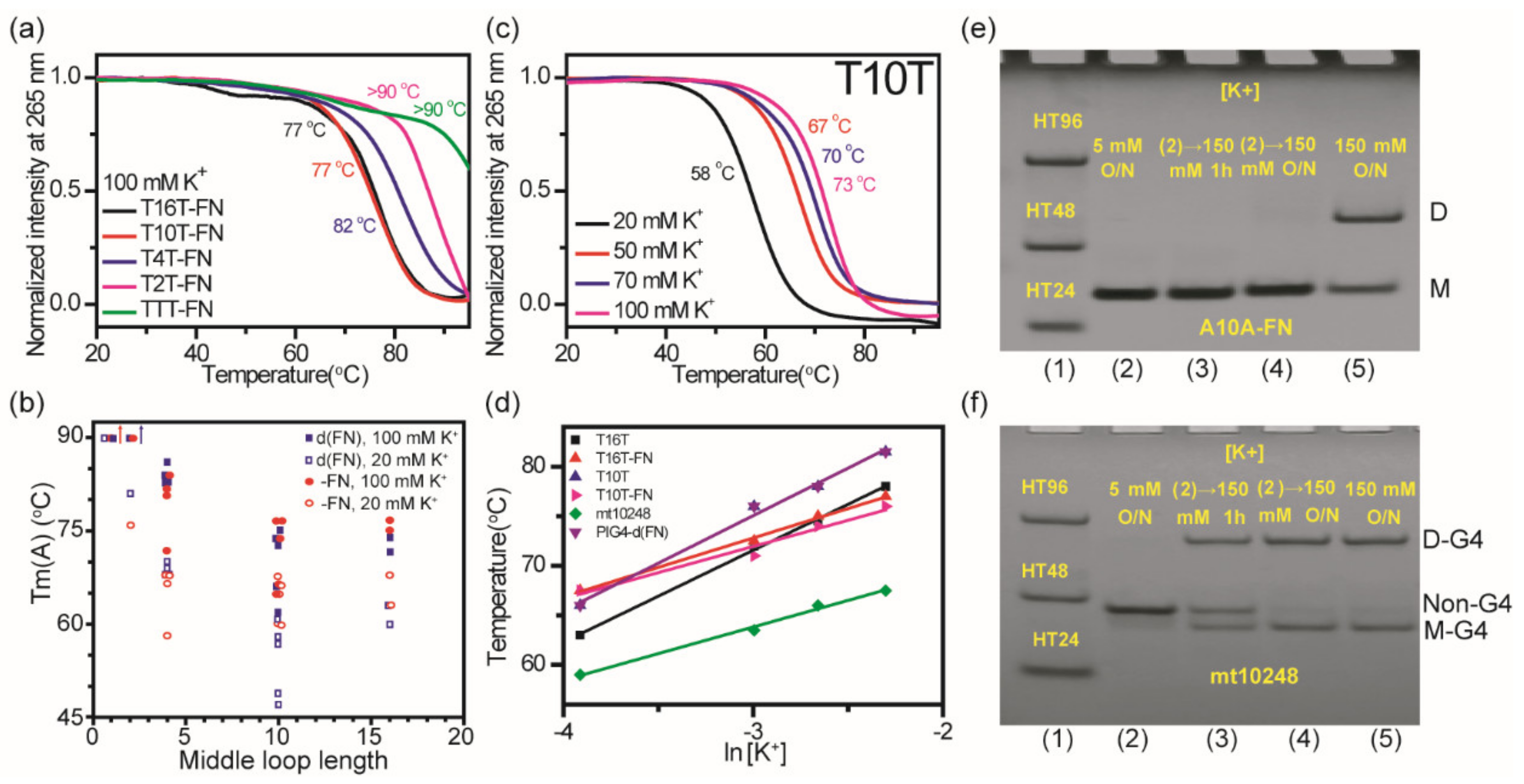
2.7. Effect of K+ Concentration on Thermal Stability of (G3HG3NmG3HG3) G4 Structures
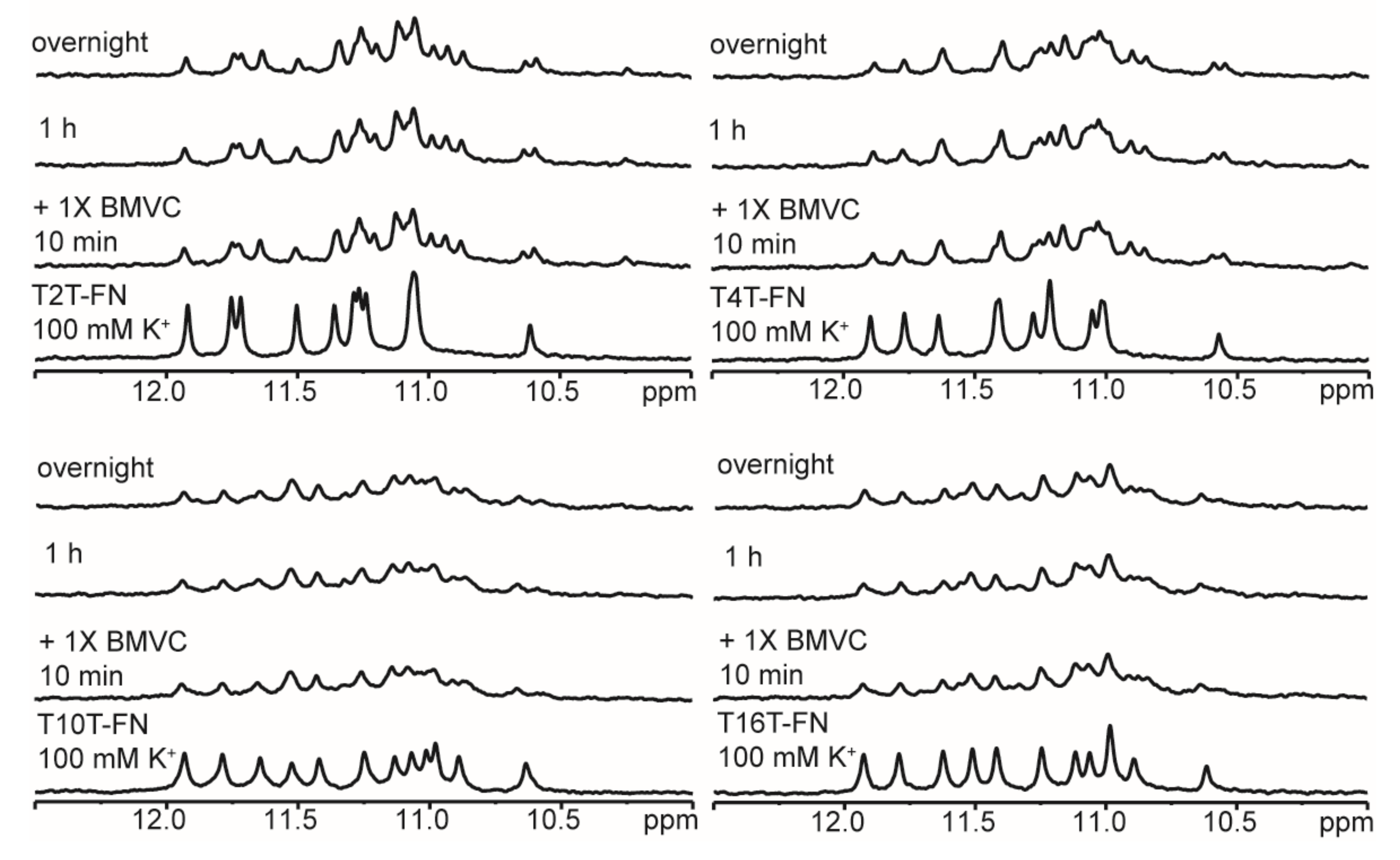
2.8. Ligand Binding to (G3TG3NmG3TG3)-FN G4 Structures
3. Discussion
3.1. Structural Diversity and Similarity of (G3HG3NmG3HG3) Sequences with and without Flanking Nucleotides
3.2. Effects of Intramolecular and Intermolecular WC Hydrogen Bonds on G4 Structures
3.3. Effects of Potassium Concentration on (G3HG3NmG3HG3)-FN G4 Structures
3.4. Implication to Native Sequences
3.5. Similarity for the Development of G4 Binding Ligands
4. Materials and Methods
4.1. DNA Preparation
4.2. Circular Dichroism (CD)
4.3. Nuclear Magnetic Resonance (NMR) Spectroscopy
4.4. Polyacrylamide Gel Electrophoresis (PAGE)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huppert, J.L.; Balasubramanian, S. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, A.K.; Johnston, M.; Neidle, S. Highly prevalent putative quadruplex sequence motifs in human DNA. Nucleic Acids Res. 2005, 33, 2901–2907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Antonio, M.D.; Smith, G.P.; Balasubramania, S. High-resolution sequencing of DNA G-quadruplex secondary structures in the human genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnov, I.; Shafer, R.H. Effect of loop sequence and size on DNA aptamer stability. Biochemistry 2000, 39, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Hazel, P.; Huppert, J.; Balasubramanian, S.; Neidle, S. Loop-length-dependent folding of G-quadruplexes. J. Am. Chem. Soc. 2004, 126, 16405–16415. [Google Scholar] [CrossRef]
- Rachwal, P.; Brown, T.; Fox, K.R. Sequence effects of single base loops in intramolecular quadruplex DNA. FEBS Lett. 2007, 581, 1657–1660. [Google Scholar] [CrossRef] [Green Version]
- Bugaut, A.; Balasubramanian, S. A sequence-dependent study of the influence of short loop lengths on the stability and topology of intramolecular DNA G-quadruplexes. Biochemistry 2008, 47, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Smargiasso, N.; Rosu, F.; Hsia, W.; Colson, P.; Baker, E.S.; Bowers, M.T.; De Pauw, E.; Gabelica, V. G-quadruplex DNA assemblies: Loop length, cation identity, and multimer formation. J. Am. Chem. Soc. 2008, 130, 10208–10216. [Google Scholar] [CrossRef]
- Guedin, A.; Gros, J.; Alberti, P.; Mergny, J.-L. How long is too long? Effects of loop size on G-quadruplex stability. Nucleic Acids Res. 2010, 38, 7858–7868. [Google Scholar] [CrossRef] [Green Version]
- Kwok, C.K.; Sherlock, M.E.; Bevilacqua, P.C. Effect of loop sequence and loop length on the intrinsic fluorescence of G-quadruplexes. Biochemistry 2013, 52, 3019–3021. [Google Scholar] [CrossRef]
- Piazza, A.; Adrian, M.; Samazan, F.; Heddi, B.; Hamon, F.; Serero, A.; Lopes, J.; Teulade-Fichou, M.-P.; Phan, A.T.; Nicolas, A. Short loop length and high thermal stability determine genomic instability induced by G-quadruplex-forming minisatellites. EMBO J. 2015, 34, 1718–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.; Cheng, Y.; Hao, J.; Jia, G.; Zhou, J.; Mergny, J.-L.; Li, C. Loop permutation affects the topology and stability of G-quadruplexes. Nucleic Acids Res. 2018, 46, 9264–9275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergny, J.L.; Helene, C. G-quadruplex DNA: A target for drug design. Nat. Med. 1998, 4, 1366–1367. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui-Jain, A.; Grand, C.L.; Bearss, D.J.; Hurley, L.H. Direct evidence for a G-quadruplex in a promoter region and its targeting with a small molecule to repress c-MYC transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11593–11598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Zyner, K.G.; Mulhearn, D.S.; Adhikari, S.; Martinez Cuesta, S.; Di Antonio, M.; Erard, N.; Hannon, G.J.; Tannahill, D.; Balasubramanian, S. Genetic interactions of G-quadruplexes in humans. eLife 2019, 8, e46793. [Google Scholar] [CrossRef]
- Zizza, P.; Cingolani, C.; Artuso, S.; Salvati, E.; Rizzo, A.; D’Angelo, C.; Porru, M.; Pagano, B.; Amato, J.; Randazzo, A.; et al. Intragenic G-quadruplex structure formed in the human CD133 and its biological and translational relevance. Nucleic Acids Res. 2016, 44, 1579–1590. [Google Scholar] [CrossRef]
- Sutherland, C.; Cui, Y.; Mao, H.; Hurley, L.H. A Mechanosensor Mechanism Controls the G-Quadruplex/i-Motif Molecular Switch in the MYC Promoter NHE III. J. Am. Chem. Soc. 2016, 138, 14138–14151. [Google Scholar] [CrossRef] [Green Version]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative visualization of DNA G-quadruplex structures in human cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef]
- Biffi, G.; Di Antonio, M.; Tannahill, D.; Balasubramanian, S. Visualization and selective chemical targeting of RNA G-quadruplex structures in the cytoplasm of human cells. Nat. Chem. 2014, 6, 75–80. [Google Scholar] [CrossRef]
- Tseng, T.Y.; Chien, C.H.; Chu, J.F.; Huang, W.C.; Lin, M.Y.; Chang, C.C.; Chang, T.C. Fluorescent probe for visualizing guanine-quadruplex DNA by fluorescence lifetime imaging microscopy. J. Biomed. Opt. 2013, 18, 101309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.C.; Tseng, T.Y.; Chen, Y.T.; Chang, C.C.; Wang, Z.F.; Wang, C.L.; Hsu, T.N.; Li, P.T.; Chen, C.T.; Lin, J.J.; et al. Direct evidence of mitochondrial G-quadruplex DNA by using fluorescent anti-cancer agents. Nucleic Acids Res. 2015, 43, 10102–10113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, T.Y.; Chen, W.W.; Chu, I.T.; Wang, C.L.; Chang, C.C.; Lin, M.C.; Lou, P.J.; Chang, T.C. The G-quadruplex fluorescent probe 3, 6-bis (1-methyl-2-vinyl-pyridinium) carbazole diiodide as a biosensor for human cancers. Sci. Rep. 2018, 8, 16082. [Google Scholar] [CrossRef]
- Chu, I.-T.; Wu, C.C.; Chang, T.C. G-quadruplex formation by single-base mutation or deletion of mitochondrial DNA sequences. Biochim. Biophys. Acta 2019, 1863, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, A.; Chen, D.; Dai, J.; Jones, R.A.; Yang, D. Solution structure of the biologically relevant G-quadruplex element in the human c-MYC promoter. Implications for G-quadruplex stabilization. Biochemistry 2005, 44, 2048–2058. [Google Scholar] [CrossRef]
- Liu, W.; Lin, C.; Wu, G.; Dai, J.; Chang, T.C.; Yang, D. Structures of 1:1 and 2:1 complexes of BMVC and MYC promoter G-quadruplex reveal a mechanism of ligand conformation adjustment for G4-recognition. Nucleic Acids Res. 2019, 47, 11931–11942. [Google Scholar] [CrossRef]
- Kuryavy, V.; Patel, D.J. Solution structure of a unique G-quadruplex scaffold adopted by a guanosine-rich human intronic sequence. Structure 2010, 18, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.W.; Lacroix, L.; Yue, D.J.E.; Lim, J.K.C.; Lim, J.M.W.; Phan, A.T. Coexistence of two distinct G-quadruplex conformations in the hTERT promoter. J. Am. Chem. Soc. 2010, 132, 12331–12342. [Google Scholar] [CrossRef]
- Onel, B.; Carver, M.; Wu, G.; Timonina, D.; Kalarn, S.; Larriva, M.; Yang, D. A new G-quadruplex with hairpin loop immediately upstream of the human BCL2 P1 promoter modulates transcription. J. Am. Chem. Soc. 2016, 138, 2563–2570. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.W.; Phan, A.T. Structural basis of DNA quadruplex-duplex junction formation. Angew. Chem. 2013, 52, 8566–8569. [Google Scholar] [CrossRef]
- Tran, P.L.T.; De Cian, A.; Gros, J.; Moriyama, R.; Mergny, J.L. Tetramolecular quadruplex stability and assembly. In Quadruplex Nucleic Acids: Topics in Current Chemistry; Chaires, J., Graves, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 330, pp. 243–273. [Google Scholar]
- Olsen, C.M.; Lee, H.T.; Marky, L.A. Unfolding thermodynamics of intramolecular G-quadruplexes: Base sequence contributions of the loops. J. Phys. Chem. B 2009, 113, 2587–2595. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.T.; Tseng, T.Y.; Wang, Z.F.; Chang, T.C. Structural conversion of intramolecular and intermolecular G-quadruplexes of bcl2mid: The effect of potassium concentration and ion exchange. J. Phys. Chem. B 2011, 115, 2360–2370. [Google Scholar] [CrossRef] [PubMed]
- Rachwal, P.A.; Findlow, I.S.; Werner, J.M.; Brown, T.; Fox, K.R. Intramolecular DNA quadruplexes with different arrangements of short and long loops. Nucleic Acids Res. 2007, 35, 4214–4222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, I.T.; Tseng, T.Y.; Chang, T.C. Comparison and elucidation of structural diversity and variation of G-rich sequences with a single G-base difference. J. Phys. Chem. B 2019, 123, 5423–5431. [Google Scholar] [CrossRef]
- Krauss, I.R.; Ramaswamy, S.; Neidle, S.; Haider, S.; Parkinson, G.N. Structural insights into the quadruplex-duplex 3’ interface formed from a telomeric repeat: A potential molecular target. J. Am. Chem. Soc. 2016, 138, 1226–1233. [Google Scholar] [CrossRef] [Green Version]
- Yatsunyk, L.A.; Mendoza, O.; Mergny, J.L. “Nano-oddities”: Unusual nucleic acid assemblies for DNA-based nanostructures and nanodevices. Acc. Chem. Res. 2014, 47, 1836–1844. [Google Scholar] [CrossRef]
- Kuo, M.H.; Wang, Z.F.; Tseng, T.Y.; Li, M.H.; Hsu, S.T.; Lin, J.J.; Chang, T.C. Conformational transition of a hairpin structure to G-quadruplex within the WNT1 gene promoter. J. Am. Chem. Soc. 2015, 137, 210–218. [Google Scholar] [CrossRef]
- Wang, Z.F.; Li, M.H.; Chu, I.T.; Winnerdy, F.R.; Phan, A.T.; Chang, T.C. Cytosine epigenetic modification modulates the formation of an unprecedented G4 structure in the WNT1 promoter. Nucleic Acids Res. 2020, 48, 1120–1130. [Google Scholar] [CrossRef]
- Tseng, T.Y.; Chu, I.T.; Lin, S.J.; Li, J.; Chang, T.C. Binding of small molecules to G-quadruplex DNA in cells revealed by fluorescence lifetime imaging microscopy of o-BMVC foci. Molecules 2019, 24, 35. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Not available. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence(5′→3′) | Tm(A) (°C, 20 mM K+) | Tm(A) (°C, 100 mM K+) | |
|---|---|---|---|
| T16T | GGGTGGGTTGCGCAATTGCGCTTGGGTGGG | 63 ± 1.55 (D, wM) a | 78 ± 0.65 (D, wM) |
| T10T | GGGTGGGTTGCATGCTTGGGTGGG | 58 ± 1.44 (D) | 73 ± 1.12 (D) |
| T4T | GGGTGGGCATGGGGTGGG | 68 ± 0.30 (M→H) b | 84 ± 1.56 (M→H) |
| T2T | GGGTGGGTAGGGTGGG | 81 ± 0.99 (H) | >90 (H) |
| TTT | GGGTGGGTGGGTGGG | 62 ± 1.70, >90 | 78 ± 0.92, >90 |
| T4T-1 | GGGTGGGATTAGGGTGGG | 69 ± 0.92 (H) | 85 ± 0.77 (H) |
| T4T-2 | GGGTGGGCATCGGGTGGG | 68 ± 1.61 (H) | 83 ± 0.62 (H) |
| T4T-3 | GGGTGGGACGTGGGTGGG | 70 ± 0.49 (M→H) | 86 ± 1.51 (M→H) |
| T16T-FN | TGAGGGTGGGTTGCGCAATTGCGCTTGGGTGGGTAA | 68 ± 0.25 (M) | 77 ± 0.49 (M) |
| T10T-FN | TGAGGGTGGGTTGCATGCTTGGGTGGGTAA | 68 ± 1.02 (M, wD) | 77 ± 0.62 (M, wD) |
| T4T-FN | TGAGGGTGGGCATGGGGTGGGTAA | 68 ± 0.44 (M) | 82 ± 0.35 (M, wD) |
| T2T-FN | TGAGGGTGGGTAGGGTGGGTAA | 76 ± 0.55 (M) | >90 (M) |
| TTT-FN | TGAGGGTGGGTGGGTGGGTAA | >90 (M) | >90 (M) |
| A16T | GGGAGGGTTGCGCAATTGCGCTTGGGTGGG | - (D) | 74 ± 1.21 (D) |
| A10A | GGGAGGGTTGCATGCTTGGGAGGG | 47 ± 1.10 (D) | 62 ± 0.38(D) |
| C10C | GGGCGGGTTGCATGCTTGGGCGGG | 57 ± 1.20 (D) | 74 ± 1.10 (D) |
| T10T-1 | GGGTGGGTTGCTTGCTTGGGTGGG | 61 ± 0.60 (D) | 76 ± 0.46 (D) |
| A10A-1 | GGGAGGGTTGCTTGCTTGGGAGGG | 49 ± 0.86 (D) | 66 ± 1.12 (D) |
| A16T-FN | TGAGGGAGGGTTGCGCAATTGCGCTTGGGTGGGTAA | - (M, wD) | 75 ± 0.29 (M, wD) |
| A10A-FN | TGAGGGAGGGTTGCATGCTTGGGAGGGTAA | 60 ± 0.13 (M, wD) | 66 ± 0.42 (M, D) |
| C10C-FN | TGAGGGCGGGTTGCATGCTTGGGCGGGTAA | 65 ± 0.42 (M, wD) | 74 ± 1.16 (M, D) |
| T10T-1-FN | TGAGGGTGGGTTGCTTGCTTGGGTGGGTAA | 67 ± 0.28 (M) | 77 ± 0.31 (M) |
| A10A-1-FN | TGAGGGAGGGTTGCTTGCTTGGGAGGGTAA | 59 ± 0.13 (M) | 68 ± 0.64 (M) |
| A4A-FN | TGAGGGAGGGCATGGGGAGGGTAA | 58 ± 0.36 (M, wD) | 73 ± 0.62 (M, D) |
| C4C-FN | TGAGGGCGGGCATGGGGCGGGTAA | 68 ± 0.48 (M) | 81 ± 0.74 (M, wD) |
| T4T-1-FN | TGAGGGTGGGATTAGGGTGGGTAA | 67 ± 1.63 (M) | 83 ± 0.95 (M, wD) |
| mt10248 | CCAGGGTGGGAGTAGTTCCCTGCTAAGGGAGGGTAG | 58 ± 1.06 (M, wD) | 67 ± 1.08 (D, wM) |
| chl1-d(FN) | GGGTGGGGAAGGGGTGGG | 71 ± 0.60 | 63 ± 0.39, 84 ± 0.61 (M) |
| GTERT-d(FN) | GGGAGGGGCTGGGAGGG | 70 ± 0.66 | 84 ± 1.48 (M→H) |
| PIG4-d(FN) | GGGCGGGAGCGCGGCGGGCGGGCGGG | - | 82 ± 1.52 (D) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Chu, I.-T.; Yeh, T.-A.; Chen, D.-Y.; Wang, C.-L.; Chang, T.-C. Effects of Length and Loop Composition on Structural Diversity and Similarity of (G3TG3NmG3TG3) G-Quadruplexes. Molecules 2020, 25, 1779. https://doi.org/10.3390/molecules25081779
Li J, Chu I-T, Yeh T-A, Chen D-Y, Wang C-L, Chang T-C. Effects of Length and Loop Composition on Structural Diversity and Similarity of (G3TG3NmG3TG3) G-Quadruplexes. Molecules. 2020; 25(8):1779. https://doi.org/10.3390/molecules25081779
Chicago/Turabian StyleLi, Jie, I-Te Chu, Ting-An Yeh, De-Yu Chen, Chiung-Lin Wang, and Ta-Chau Chang. 2020. "Effects of Length and Loop Composition on Structural Diversity and Similarity of (G3TG3NmG3TG3) G-Quadruplexes" Molecules 25, no. 8: 1779. https://doi.org/10.3390/molecules25081779