
Cucurbitacin E Inhibits Huh7 Hepatoma Carcinoma Cell Proliferation and Metastasis via Suppressing MAPKs and JAK/STAT3 Pathways

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
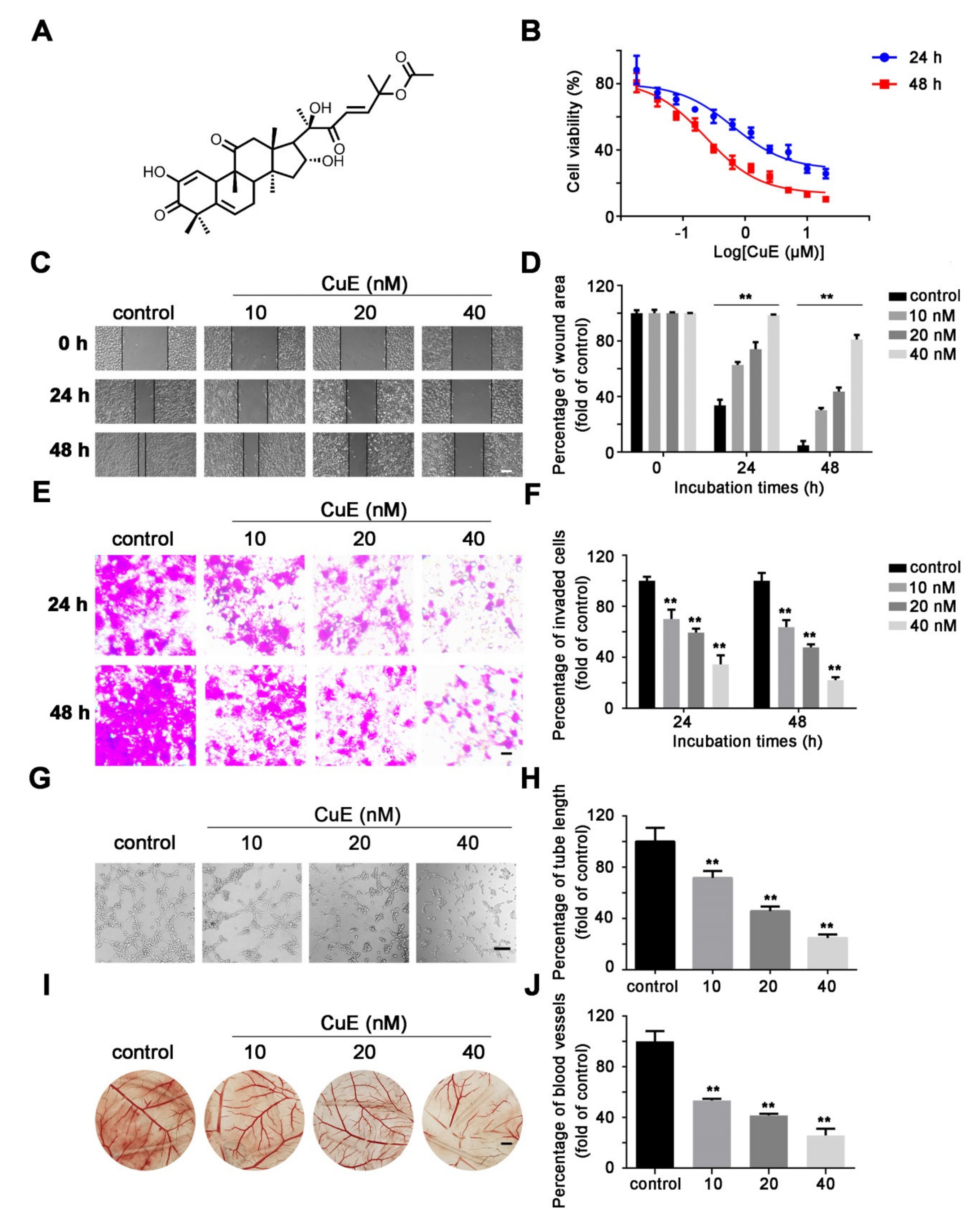
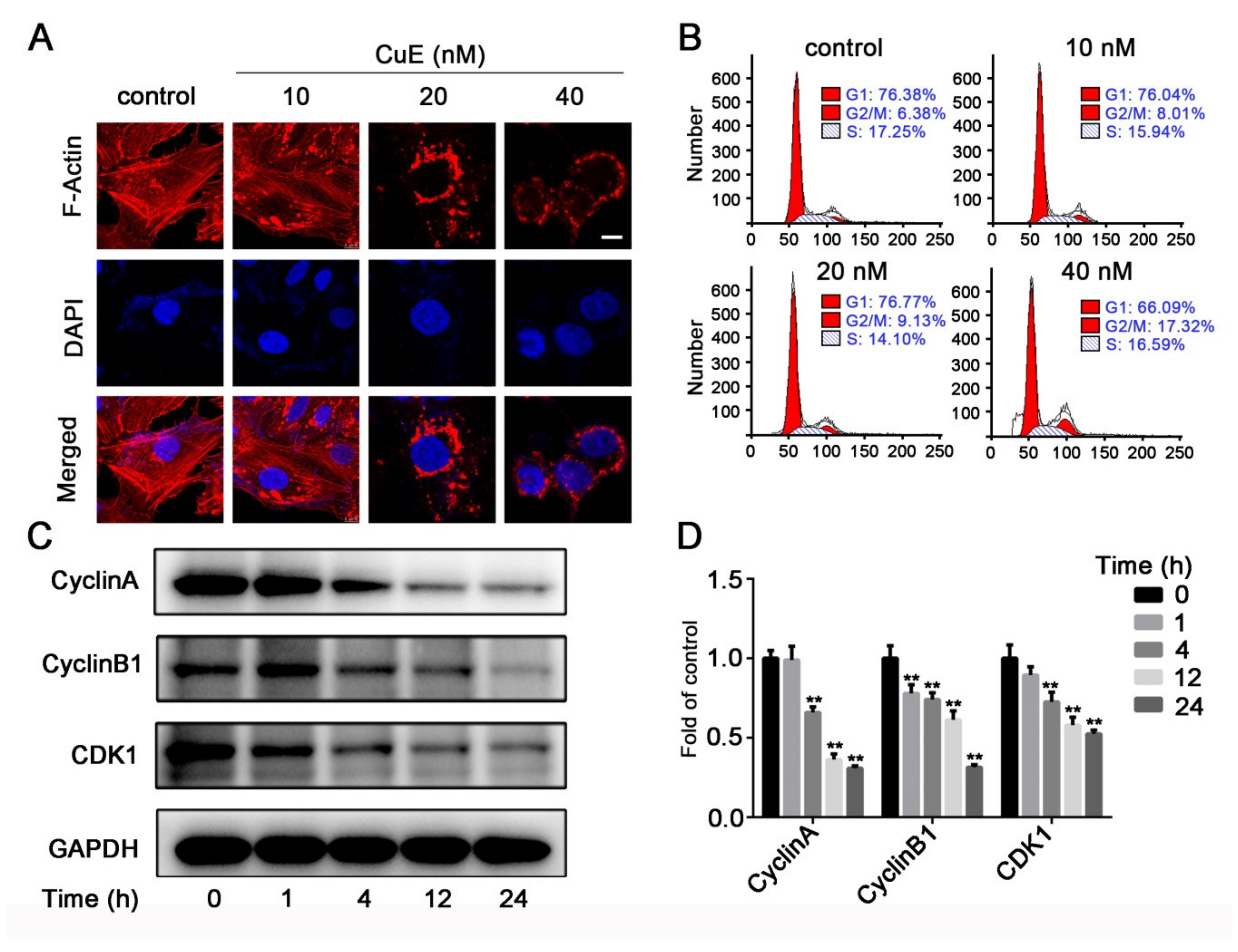
2.1. CuE Inhibited the Proliferation, Migration and Angiogenesis of Huh7 Cells
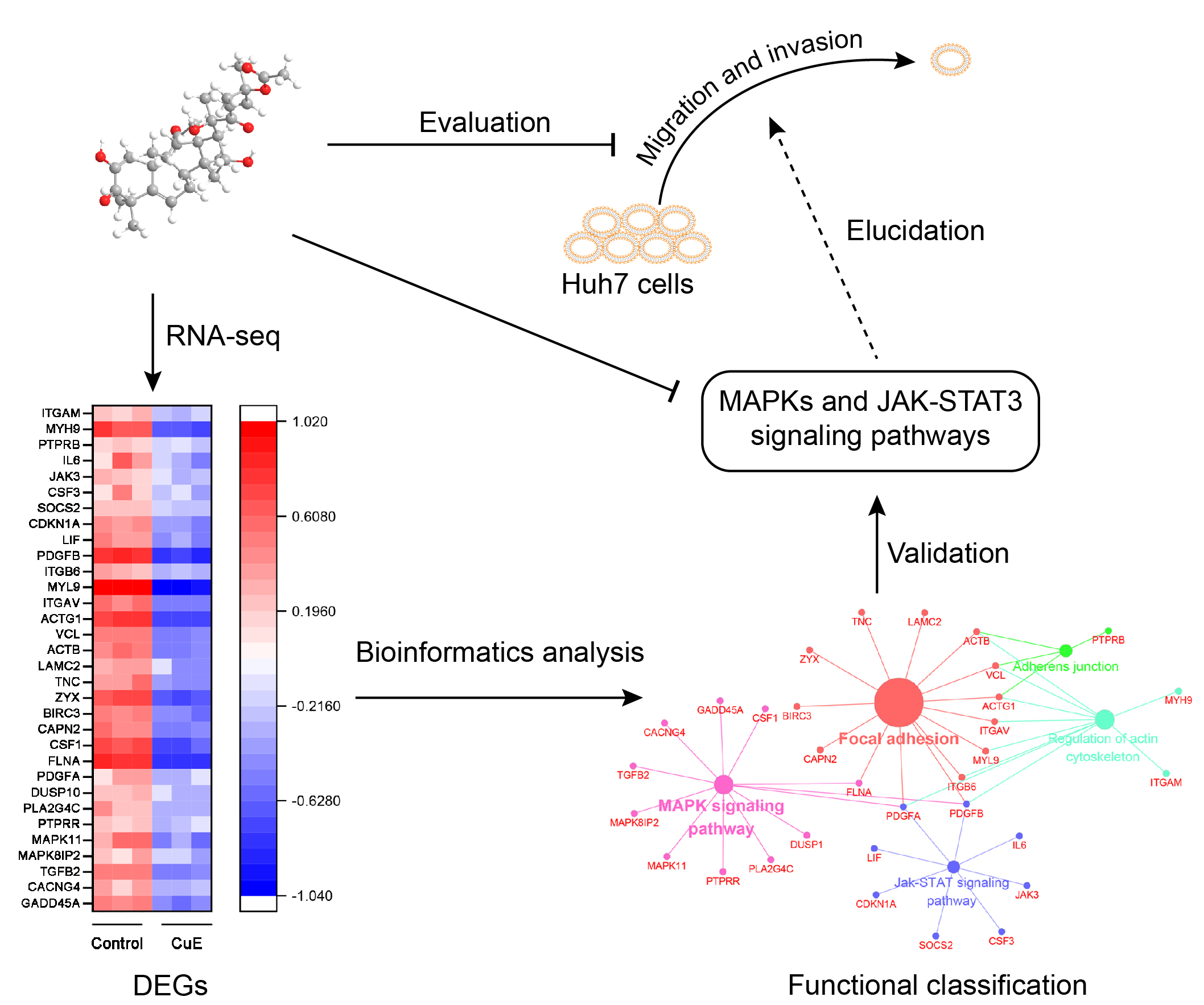
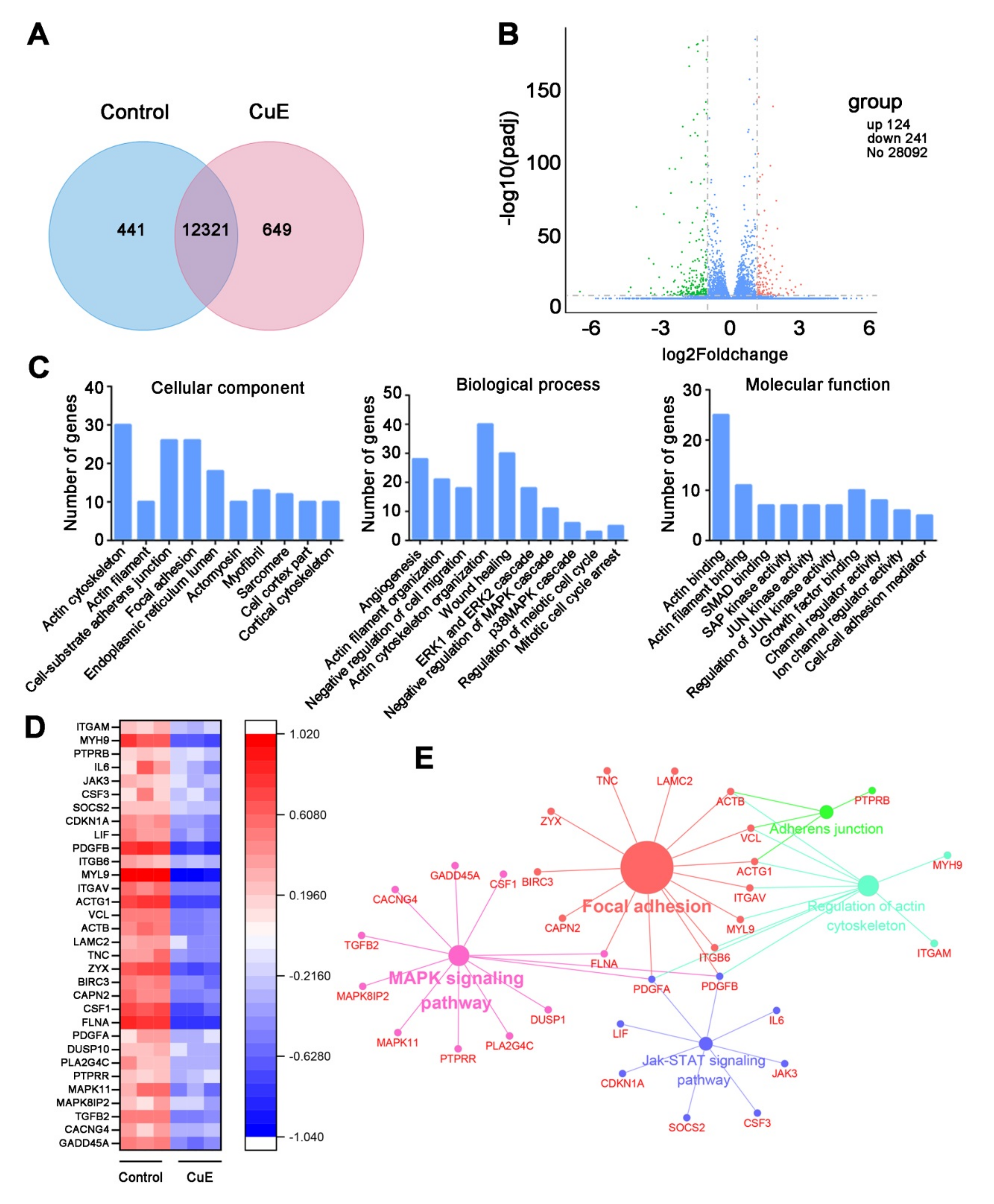
2.2. Bioinformatics Analysis for the CuE-Regulated Signaling Networks
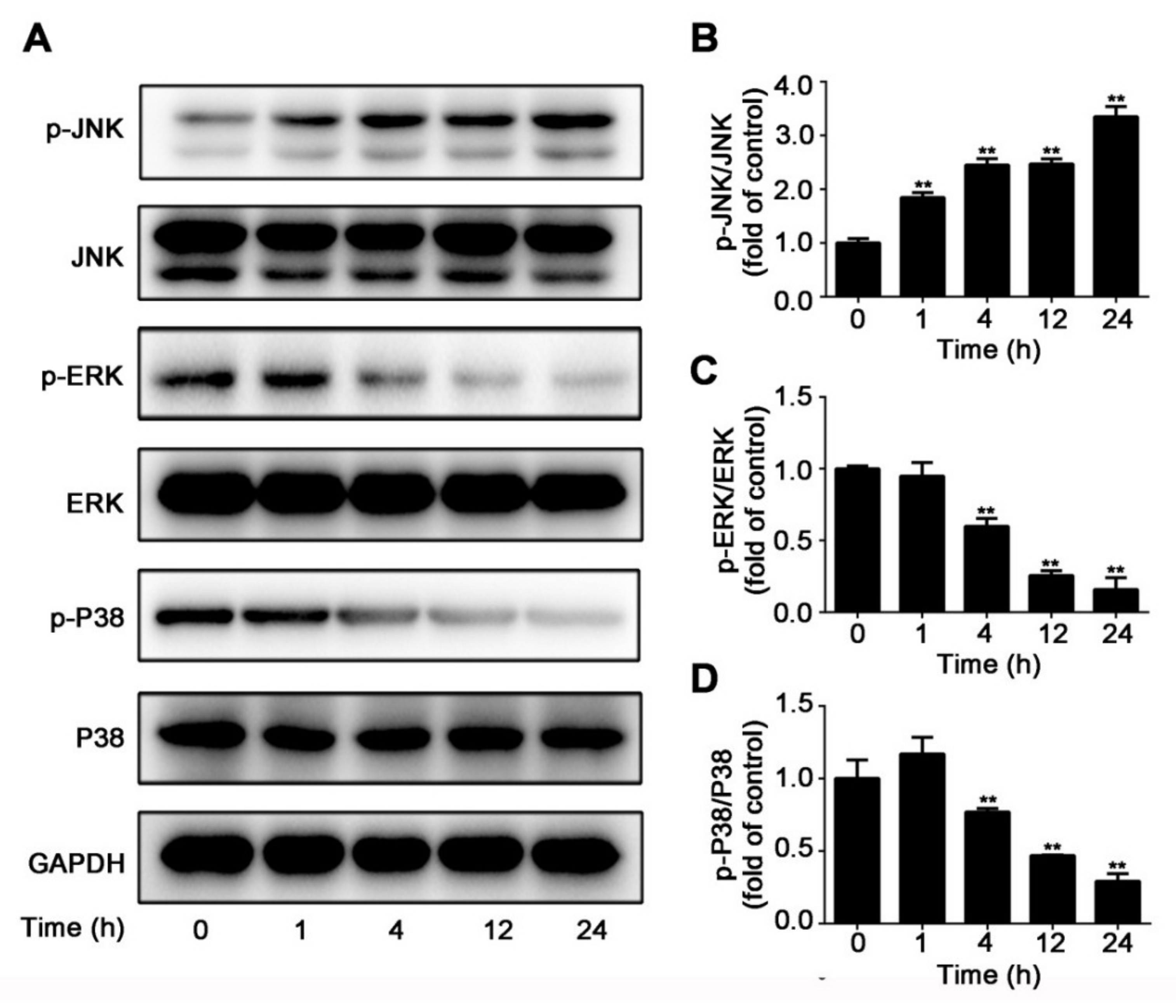
2.3. CuE Inhibited JNK/ERK/p38 MAPK Pathways in Huh7 Cells
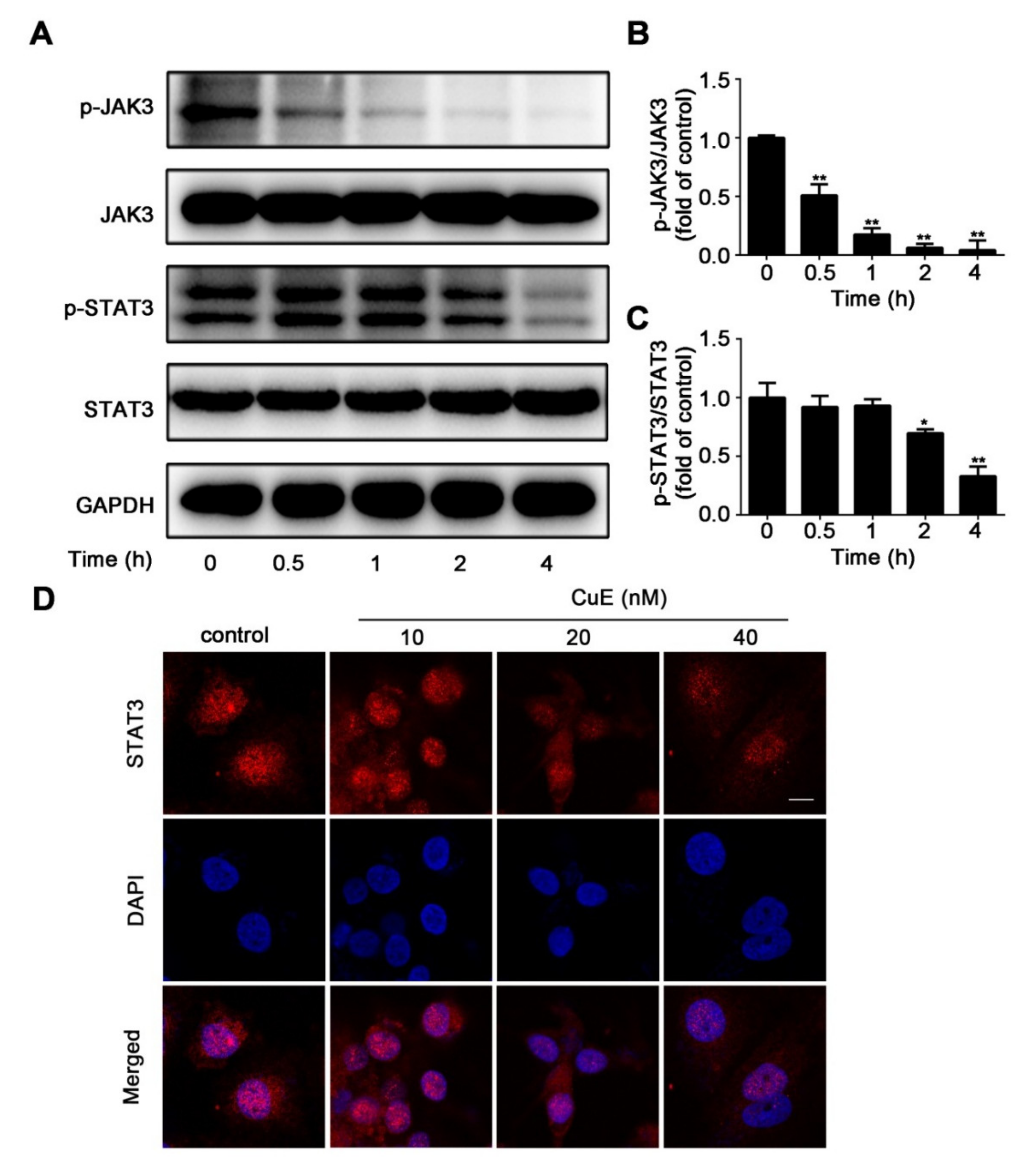
2.4. CuE Inhibited JAK3/STAT3 Signaling
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell lines and Culture
4.3. Wound Healing Assay
4.4. Transwell Migration Assay
4.5. Tube Formation Assay
4.6. Chorioallantoic Membranes Assay (CAM)
4.7. Cell Cycle Distribution Analysis
4.8. Rhodamine-Phalloidin Staining and Fluorescence Microscopy
4.9. Immunofluorescent Analysis
4.10. Western Blot Analysis
4.11. RNA Sequencing Assay
Differentially Expressed Genes (DEGs) Library Construction and Sequencing
Functional Classification of DEGs
Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dimitroulis, D.; Damaskos, C.; Valsami, S.; Davakis, S.; Garmpis, N.; Spartalis, E.; Athanasiou, A.; Moris, D.; Sakellariou, S.; Kykalos, S.; et al. From diagnosis to treatment of hepatocellular carcinoma: An epidemic problem for both developed and developing world. World J. Gastroenterol. 2017, 23, 5282–5294. [Google Scholar] [CrossRef]
- Forner, A.; Reig, M.; Bruix, J. Hepatocellular carcinoma. Lancet 2018, 391, 1301–1314. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.T.; He, W.; Song, S.S.; Wei, W. Paeoniflorin inhibited the tumor invasion and metastasis in human hepatocellular carcinoma cells. Bratisl. Med. J. 2014, 115, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, G.L.; Zeng, S.; Shen, H. Chemotherapy and target therapy for hepatocellular carcinoma: New advances and challenges. World J. Hepatol 2015, 7, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.C.; Chow, P.K.; Allen, J.C.; Chia, G.S.; Lim, M.; Cheow, P.C.; Chung, A.Y.; Ooi, L.L.; Tan, S.B. Microvascular invasion is a better predictor of tumor recurrence and overall survival following surgical resection for hepatocellular carcinoma compared to the Milan criteria. Ann. Surg. 2011, 254, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Moulick, K.; Ahn, J.H.; Zong, H.; Rodina, A.; Cerchietti, L.; Gomes DaGama, E.M.; Caldas-Lopes, E.; Beebe, K.; Perna, F.; Hatzi, K.; et al. Affinity-based proteomics reveal cancer-specific networks coordinated by Hsp90. Nat. Chem. Biol. 2011, 7, 818–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuaje, F.; Tiemann, K.; Niclou, S.P. Therapeutic control and resistance of the EGFR-driven signaling network in glioblastoma. Cell Commun. Signal. 2015, 13, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banudevi, S.; Swaminathan, S.; Maheswari, K.U. Pleiotropic role of dietary phytochemicals in cancer: Emerging perspectives for combinational therapy. Nutr. Cancer 2015, 67, 1021–1048. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Maryam, A.; Qazi, J.I.; Ma, T. Targeting apoptosis and multiple signaling pathways with icariside II in cancer cells. Int. J. Biol. Sci. 2015, 11, 1100–1112. [Google Scholar] [CrossRef] [Green Version]
- Pao, H.P.; Liao, W.I.; Wu, S.Y.; Hung, K.Y.; Huang, K.L.; Chu, S.J. PG490-88, a derivative of triptolide, suppresses ischemia/reperfusion-induced lung damage by maintaining tight junction barriers and targeting multiple signaling pathways. Int. Immunopharmacol. 2019, 68, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Rejhová, A.; Opattová, A.; Cumová, A.; Slíva, D.; Vodička, P. Natural compounds and combination therapy in colorectal cancer treatment. Eur. J. Med. Chem. 2018, 144, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Ma, Y.; An, N.; Sun, P.; Wang, Y.; Sun, C. Clinical efficacy of combination of pertuzumab, trastuzumab, and docetaxel for treatment of patients with HER2-positive breast cancer. Medicine 2019, 98, 38. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Qu, H.; Zhu, W.; Xu, K.; Xu, A.; Jia, B.; Qing, Y.; Li, H.; Wei, H.J.; Zhao, H.Y. Low-dose paclitaxel inhibits tumor cell growth by regulating glutaminolysis in colorectal carcinoma cells. Front. Pharmacol. 2017, 8, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolleddula, J.; Navindra, P.S.; Muraleedharan, G.N. Anticancer and antiinflammatory activities of cucurbitacins from Cucurbita andreana. Cancer Lett. 2003, 189, 11–16. [Google Scholar]
- Wu, D.; Wang, Z.; Lin, M.; Shang, Y.; Wang, F.; Zhou, J.; Luo, X.; Huang, W. In vitro and in vivo anti-tumor activity of cucurbitacin C, a novel natural product from cucumber. Front. Pharmacol. 2019, 10, 1287. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.L.; Andújar, I.; Escandell, J.M.; Giner, R.M.; Recio, M.C. Cucurbitacins as inducers of cell death and a rich source of potential anticancer compounds. Curr. Pharm. Design 2012, 18, 1663–1676. [Google Scholar]
- Park, C.S.; Lim, H.; Han, K.J.; Baek, S.H.; Sohn, H.O.; Lee, D.W.; Kim, Y.G.; Yun, H.Y.; Baek, K.J.; Kwon, N.S. Inhibition of nitric oxide generation by 23,24-dihydrocucurbitacin D in mouse peritoneal macrophages. J. Pharmacol. Exp. Ther. 2004, 309, 705–710. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.M.; Shen, C.J.; Chang, C.C.; Chou, C.Y.; Tsai, C.C.; Hsu, Y.C. Inducement of apoptosis by cucurbitacin E, a tetracyclic triterpenes, through death receptor 5 in human cervical cancer cell lines. Cell Death Discov. 2017, 3, 17014. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Huang, T.Y.; Chen, M.J. Therapeutic ROS targeting of GADD45 gamma in the induction of G2/M arrest in primary human colorectal cancer cell lines by cucurbitacin E. Cell Death Dis. 2014, 5, e1198. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.M.; Chang, C.C.; Lin, C.W.; Ko, S.Y.; Hsu, Y.C. Cucurbitacin E as inducer of cell death and apoptosis in human oral squamous cell carcinoma cell line SAS. Int. J. Mol. Sci. 2013, 14, 17147–17156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jevtić, B.; Djedović, N.; Stanisavljević, S.; Despotović, J.; Miljković, D.; Timotijević, G. Cucurbitacin E potently modulates the activity of encephalitogenic cells. J. Agric. Food Chem. 2016, 64, 4900–4907. [Google Scholar] [CrossRef] [PubMed]
- Qiao, J.; Xu, L.H.; He, J.; Ouyang, D.Y.; He, X.H. Cucurbitacin E exhibits anti-inflammatory effect in RAW 264.7 cells via suppression of NF-kappaB nuclear translocation. Inflamm. Res. 2013, 62, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, S.; Wu, Y.; Zhang, J. Cucurbitacin E inhibits osteosarcoma cells proliferation and invasion through attenuation of PI3K/AKT/mTOR signalling pathway. Biosci. Rep. 2016, 36, e00405. [Google Scholar] [CrossRef]
- Maind, A.; Raut, S. Mining conditions specific hub genes from RNA-Seq gene-expression data via biclustering and their application to drug discovery. IET Syst. Biol. 2019, 13, 194–203. [Google Scholar] [CrossRef]
- Zhao, B.; Shen, C.; Zheng, Z.; Wang, X.; Zhao, W.; Chen, X.; Peng, F.; Xue, L.; Shu, M.; Hou, X.; et al. Peiminine inhibits glioblastoma in vitro and in vivo through cell cycle arrest and autophagic flux blocking. Cell Physiol. Biochem. 2018, 51, 1566–1583. [Google Scholar] [CrossRef]
- Yang, B.; Wang, N.; Wang, S.; Li, X.; Zheng, Y.; Li, M.; Song, J.; Zhang, F.; Mei, W.; Lin, Y.; et al. Network-pharmacology-based identification of caveolin-1 as a key target of Oldenlandia diffusa to suppress breast cancer metastasis. Biomed. Pharmacother. 2019, 112, 108607. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Q.; Dahan, A.; Xue, J.; Wei, L.; Tan, W.; Zhang, G. Transcriptomic analyses reveal the molecular mechanisms of schisandrin B alleviates CCl4-induced liver fibrosis in rats by RNA-sequencing. Chem. Biol. Interact. 2019, 309, 108675. [Google Scholar] [CrossRef]
- Galustian, C.; Dye, J.; Leach, L.; Clark, P.; Firth, J.A. Actin cytoskeletal isoforms in human endothelial cells in vitro: Alteration with cell passage. Vitro Cell. Dev. Biol. Anim. 1995, 31, 796–802. [Google Scholar] [CrossRef]
- Mark, D.D.; Michael, C.A.; Edward, A.S. Cucurbitacin E induced disruption of the actin and vimentin cytoskeleton in prostate carcinoma cells. Biochem. Pharmacol. 1996, 52, 1553–1560. [Google Scholar]
- Statsuk, A.V.; Bai, R.; Baryza, J.L.; Verma, V.A.; Hamel, E.; Wender, P.A.; Kozmin, S.A. Actin is the primary cellular receptor of bistramide A. Nat. Chem. Biol. 2005, 1, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.O.; Ahn, C.H.; Yang, I.H.; Han, J.M.; Shin, J.A.; Cho, S.D.; Hong, S.D. Norcanthardin suppresses YD-15 cell invasion through inhibition of FAK/paxillin and F-actin reorganization. Molecules 2019, 24, 1928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Yang, J.; Song, P. Correlation of ERK/MAPK signaling pathway with proliferation and apoptosis of colon cancer cells. Oncol. Lett. 2019, 17, 2266–2270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lake, D.; Corrêa, S.A.; Müller, J. Negative feedback regulation of the ERK1/2 MAPK pathway. Cell. Mol. Life Sci. 2016, 73, 4397–4413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Yang, X.; Geng, M.; Huang, M. Targeting ERK, an Achilles’ Heel of the MAPK pathway, in cancer therapy. Acta Pharm. Sin. B 2018, 8, 552–562. [Google Scholar] [CrossRef]
- Chen, D.; Ma, Y.; Li, P.; Liu, M.; Fang, Y.; Zhang, J.; Zhang, B.; Hui, Y.; Yin, Y. Piperlongumine induces apoptosis and synergizes with doxorubicin by inhibiting the JAK2/STAT3 pathway in triple-negative breast cancer. Molecules 2019, 24, 2238. [Google Scholar] [CrossRef] [Green Version]
- Khandelwal, R.; Chauhan, A.P.S.; Bilawat, S.; Gandhe, A.; Hussain, T.; Hood, E.A.; Nayarisseri, A.; Singh, S.K. Strcture-based virtual screening for the identification of high-affinity small molecule towards STAT3 for the clinical treatment of Osteosarcoma. Curr. Top. Med. Chem. 2018, 18, 2511–2526. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compound CuE is available from the authors. |





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yang, H.; Guo, Q.; Liu, T.; Jiang, Y.; Zhao, M.; Zeng, K.; Tu, P. Cucurbitacin E Inhibits Huh7 Hepatoma Carcinoma Cell Proliferation and Metastasis via Suppressing MAPKs and JAK/STAT3 Pathways. Molecules 2020, 25, 560. https://doi.org/10.3390/molecules25030560
Liu Y, Yang H, Guo Q, Liu T, Jiang Y, Zhao M, Zeng K, Tu P. Cucurbitacin E Inhibits Huh7 Hepatoma Carcinoma Cell Proliferation and Metastasis via Suppressing MAPKs and JAK/STAT3 Pathways. Molecules. 2020; 25(3):560. https://doi.org/10.3390/molecules25030560
Chicago/Turabian StyleLiu, Yang, Heng Yang, Qiang Guo, Tingting Liu, Yong Jiang, Mingbo Zhao, Kewu Zeng, and Pengfei Tu. 2020. "Cucurbitacin E Inhibits Huh7 Hepatoma Carcinoma Cell Proliferation and Metastasis via Suppressing MAPKs and JAK/STAT3 Pathways" Molecules 25, no. 3: 560. https://doi.org/10.3390/molecules25030560