Essential Oil Composition of Alluaudia procera and in Vitro Biological Activity on Two Drug-Resistant Models

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
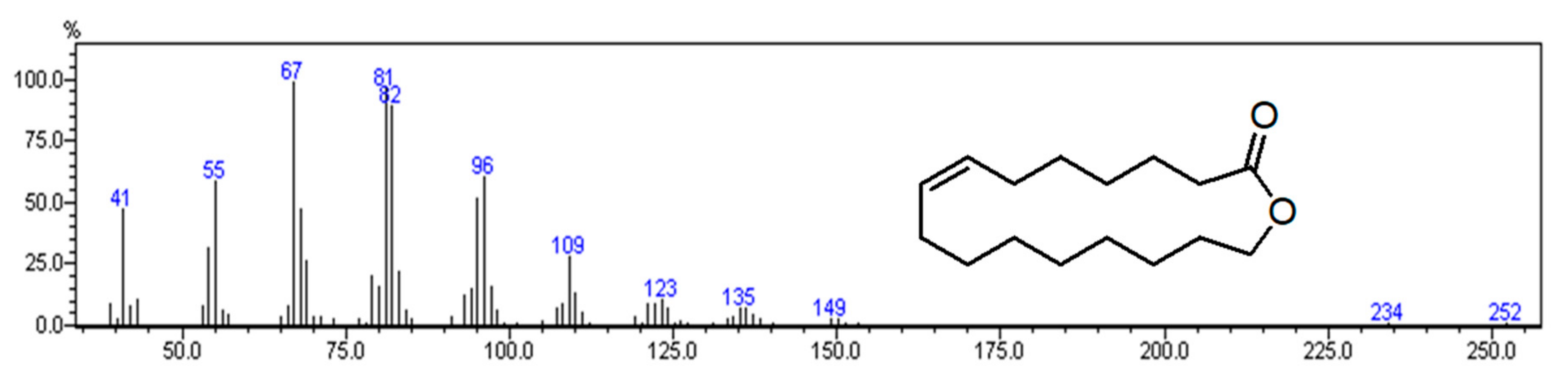
2.1. Chemical Composition
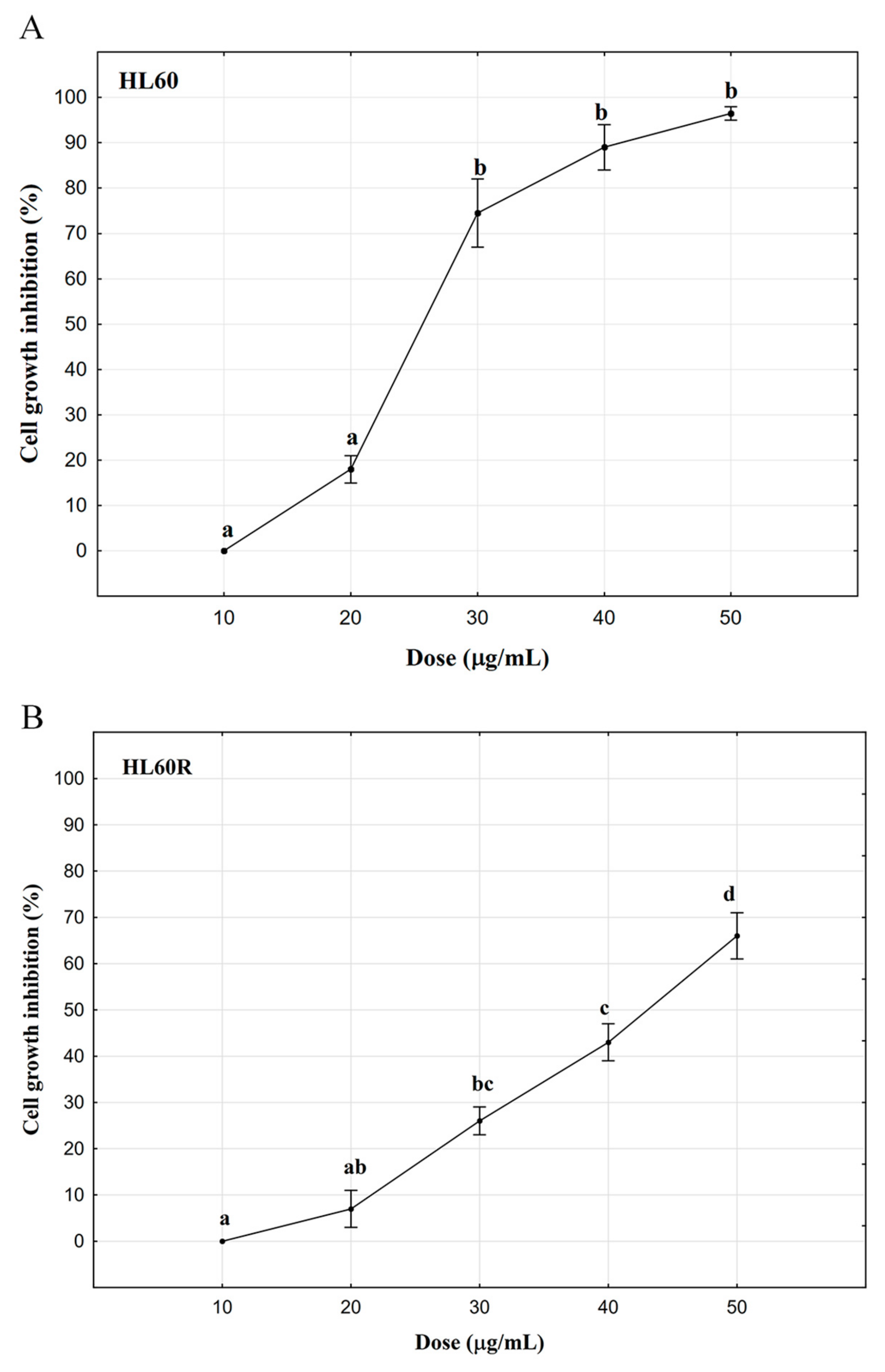
2.2. In Vitro Anticancer Activity
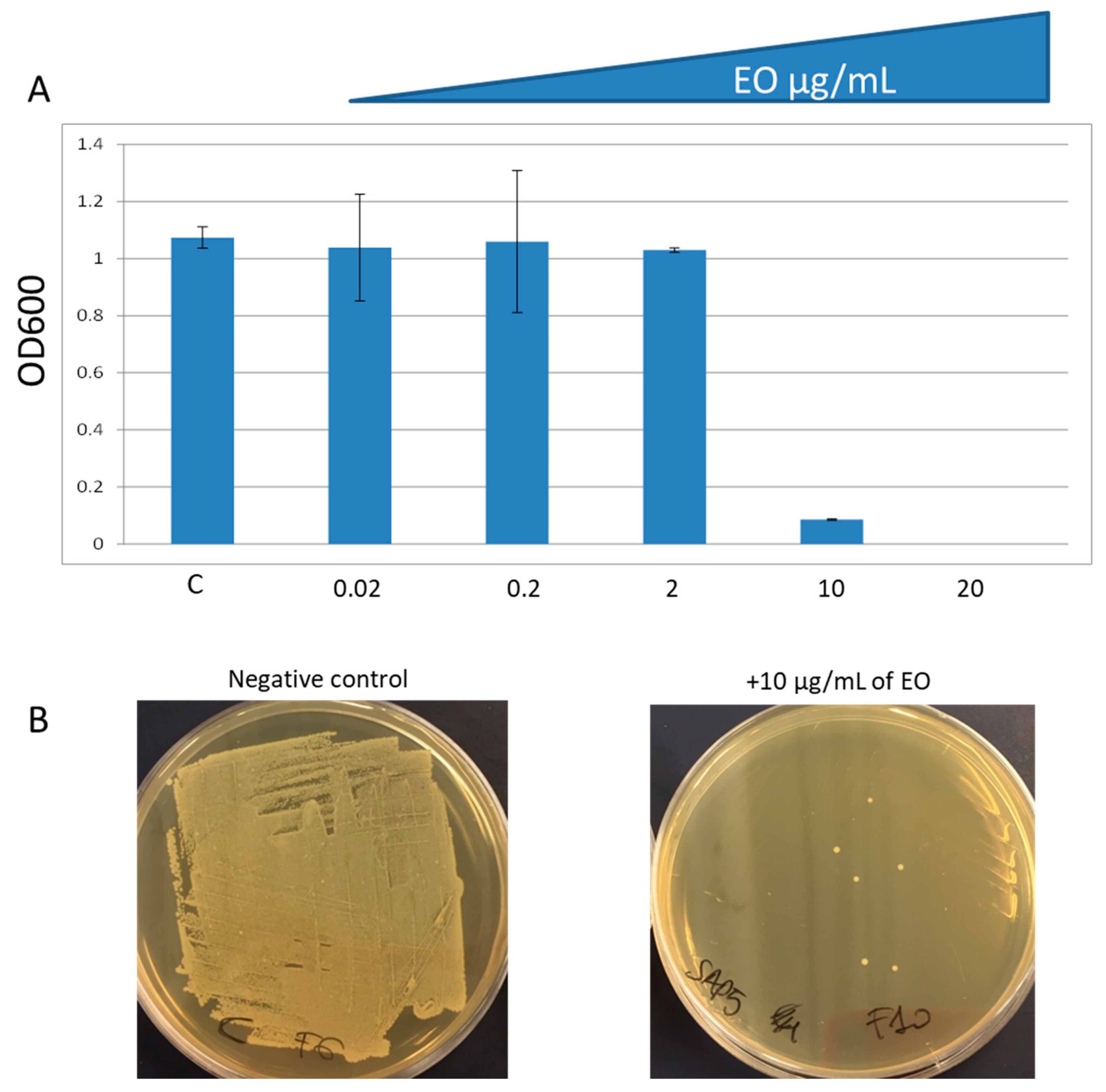
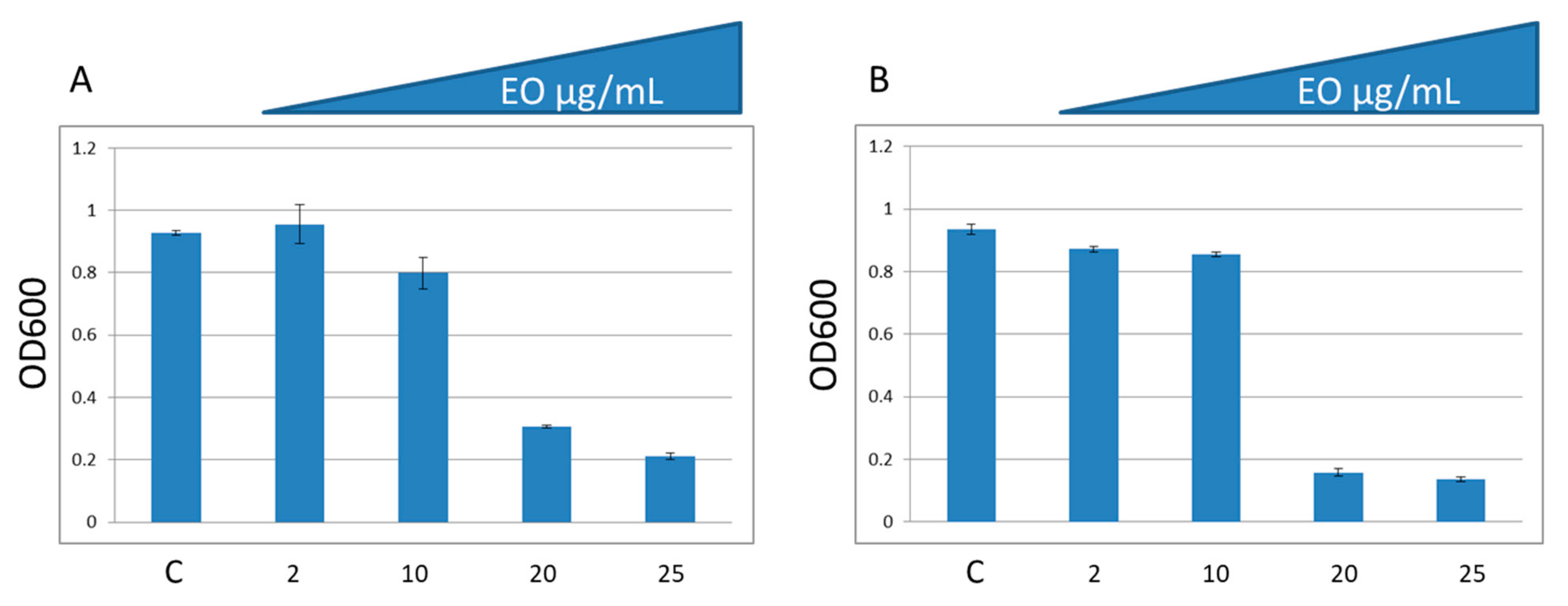
2.3. In Vitro Antibacterial Activity
3. Discussion
4. Materials and Methods
4.1. Plant Species
4.2. Plant Material
4.3. Essential Oil Isolation and Chemical Characterization
4.4. Identification of Compounds
4.5. Cell Culture
4.6. Cytotoxicity Assay
4.7. Microorganisms
4.8. Determination of the Antibacterial Activity
4.9. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sajeva, M.; Costanzo, M. Succulents, the Illustrated Dictionary; Timber Press: Portland, OR, USA, 1994. [Google Scholar]
- Crowley, B.E.; Godfrey, L.R. Why all those spines? Anachronistic defences in the Didiereoideae against now extinct lemurs. S. Afr. J. Sci. 2013, 109, 1–7. [Google Scholar] [CrossRef]
- Gould, L.; Kelley, E.A.; LaFleur, M. Reproductive female feeding strategies in spiny forest-dwelling Lemur catta in Southern and Southwestern Madagascar: How do females Meet the Challenges of Reproduction in this Harsh Habitat? Folia Primatol. 2015, 86, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Droscher, I.; Rothman, J.M.; Ganzhorn, J.U.; Kappeler, P.M. Nutritional consequences of folivory in a small-bodied lemur (Lepilemur Leucopus): Effects of season and reproduction on nutrient balancing. Am. J. Phys. Anthropol. 2016, 160, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Génin, F. Who sleeps with whom? Sleeping association and socio-territoriality in Microcebus griseorufus. J. Mammal. 2010, 91, 942–951. [Google Scholar] [CrossRef]
- Gardner, C.; Ferguson, B.; Rebara, F.; Ratsifandrihamanana, N. Integrating traditional values and management regimes into Madagascar’s expanded protected area system: The case of Ankodida. In Protected Landscapes and Cultural and Spiritual Values; Mallarach, J.M., Ed.; Volume 2 in the Series Values of Protected Landscapes and Seascapes, IUCN, GTZ and Obra Social de Caixa Catalunya; Kasparek Verlag: Heidelberg, Germany, 2008; pp. 92–103. ISBN 978-3-925064-60-9. [Google Scholar]
- Yoshioka, Y.; Ohshima, H.; Maeda, Y. Effects of Soil Chemical Properties on Vegetation of Alluaudia procera Drake, a Species Endemic to Madagascar. J. Agric. Sci. Tokyo Univ. Agric. 2013, 58, 117–124. [Google Scholar]
- Tengö, M.; von Heland, J. Trees and Tree-Planting in Southern Madagascar: Sacredness and Remembrance. In Greening in the Red Zone: Disaster, Resilience and Community Greening; Tidball, K.G., Krasny, M.E., Eds.; Springer Science + Business Media: Dordrecht, The Netherlands, 2014; pp. 334–337. [Google Scholar] [CrossRef]
- Linholm, P.; Gullbo, J.; Claeson, P.; Goeransson, U.; Johansson, S.; Backlund, A.; Larsson, R.; Bohlin, L. Selective cytotoxicity evaluation in anticancer drug screening of fractionated plant extracts. J. Biomol. Screen. 2002, 7, 333–340. [Google Scholar] [CrossRef]
- Rasamoelisendra, R.; Voirin, B.; Favre-Bonvin, J.; Andriantsiferana, M.; Rabesa, Z. 6-C-Methyl-and 6, 8-di-C-methyl-3, 7-di-O-methylkaempferol from Alluaudia dumosa. Phytochemistry 1989, 28, 1996–1997. [Google Scholar] [CrossRef]
- Poma, P.; Labbozzetta, M.; Notarbartolo, M.; Bruno, M.; Maggio, A.; Rosselli, S.; Sajeva, M.; Zito, P. Chemical composition, in vitro antitumor and pro-oxidant activities of Glandora rosmarinifolia (Boraginaceae) essential oil. PLoS ONE 2018, 13, e0196947. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Dudareva, N.; Negre, F.; Dinesh, A.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Zito, P.; Labbozzetta, M.; Notarbartolo, M.; Sajeva, M.; Poma, P. Essential oil of Cyphostemma juttae (Vitaceae): Chemical composition and antitumor mechanism in triple negative breast cancer cells. PLoS ONE 2019, 14, e0214594. [Google Scholar] [CrossRef]
- Notarbartolo, M.; Cervello, M.; Dusonchet, L.; Cusimano, A.; D’Alessandro, N. Resistance to diverse apoptotic triggers in multidrug resistant HL60 cells and its possible relationship to the expression of P-glycoprotein, Fas and of the novel anti-apoptosis factors IAP (inhibitory of apoptosis proteins). Cancer Lett. 2002, 180, 91–101. [Google Scholar] [CrossRef]
- Marcelletti, J.F.; Sikic, B.I.; Cripe, L.D.; Paietta, E. Evidence of a role for functional heterogeneity in multidrug resistance transporters in clinical trials of P-glycoprotein modulation in acute myeloid leukemia. Cytom. B Clin. Cytom. 2019, 96, 57–66. [Google Scholar] [CrossRef]
- Liu, B.; Li, L.J.; Gong, X.; Zhang, W.; Zhang, H.; Zhao, L. Co-expression of ATP binding cassette transporters is associated with poor prognosis in acute myeloid leukemia. Oncol. Lett. 2018, 15, 6671–6677. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, A.; Martinez, Y.Z.; Alduina, R.; Monteverde, V.; Molina, C.F.; Vitale, M. Comparison of two PCR methods for detection of Leptospira interrogans in formalin-fixed and paraffin-embedded tissues. Mem. Inst. Oswaldo Cruz 2012, 107, 85–88. [Google Scholar] [CrossRef]
- Vitale, M.; Scatassa, M.L.; Cardamone, C.; Oliveri, G.; Piraino, C.; Alduina, R.; Napoli, C. Staphylococcal food poisoning case and molecular analysis of toxin genes in Staphylococcus aureus strains isolated from food in Sicily, Italy. Foodborne Pathog. Dis. 2015, 12, 21–23. [Google Scholar] [CrossRef]
- Dresler, C.; Saladino, M.L.; Demirbag, C.; Caponetti, E.; Chillura Martino, D.F.; Alduina, R. Development of controlled release systems of biocides for the conservation of cultural heritage. Int. Biodeterior. Biodegrad. 2017, 125, 150–156. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging strategies to combat eskape pathogens in the era of antimicrobial resistance: A review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef]
- Mandal, S.M.; Ghosh, A.K.; Roy, A.; Hazra, T.; Franco, O.L. Challenges and future prospects of antibiotic therapy: From peptides to phages utilization. Front. Pharmacol. 2014, 5, 105. [Google Scholar] [CrossRef]
- Linstrom, P.J.; Mallard, W.G. NIST Chemistry WebBook, NIST Standard Reference Database Number 69; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2000; (last update 2018). Available online: https://webbook.nist.gov/chemistry/ (accessed on 18 June 2019).
- Zito, P.; Sajeva, M.; Bruno, M.; Rosselli, S.; Maggio, A.; Senatore, F. Essential oils composition of Periploca laevigata Aiton subsp. angustifolia (Labill.) Markgraf (Apocynaceae—Periplocoideae). Nat. Prod. Res. 2013, 27, 255–265. [Google Scholar] [CrossRef]
- Leffingwell, J.C.; Alford, E.D.; Leffingwell, D.; Penn, R. Identification of the volatile constituents of Cyprian Latakia tobacco by dynamic and static headspace analyses. Leffingwell Rep. 2013, 5, 1–29. [Google Scholar]
- Arokiyaraj, S.; Choi, S.H.; Lee, Y.; Bharanidharan, R.; Hairul-Islam, V.I.; Vijayakumar, B.; Oh, Y.K.; Dinesh-Kumar, V.; Vincent, S.; Kim, K.H. Characterization of ambrette seed oil and its mode of action in bacteria. Molecules 2015, 20, 384–395. [Google Scholar] [CrossRef]
- Witkowska-Banaszczak, E. Flavonoids from Trollius europaeus flowers and evaluation of their biological activity. J. Pharm. Pharmacol. 2018, 70, 550–558. [Google Scholar] [CrossRef]
- Hirose, S.; Joichi, A.; Nakamura, S.; Awano, K. Volatile components of musky scent of orchids. Flavour Fragr. J. 1999, 14, 183–184. [Google Scholar] [CrossRef]
- Dung, N.X.; Van Khiên, P.; Nhuân, D.D.; Hoi, T.M.; Ban, N.K.; Leclercq, P.A.; Muselli, A.; Bighelli, A.; Casanova, J. Composition of the seed oil of Hibiscus abelmoschus L. (Malvaceae) growing in Vietnam. J. Essent. Oil Res. 1999, 11, 447–452. [Google Scholar] [CrossRef]
- Nee, T.Y.; Cartt, S.; Pollard, M.R. Seed coat components of Hibiscus abelmoschus. Phytochemistry 1986, 25, 2157–2161. [Google Scholar] [CrossRef]
- Brunton, L.; Dandan, R.H.; Knollmann, B.C. Goodman and Gilman’s The Pharmacological Basis of Therapeutics, 13th ed.; McGraw-Hill: New York, NY, USA, 1996. [Google Scholar]
- Moes, D.J.; Guchelaar, H.J.; de Fijter, J.W. Sirolimus and everolimus in kidney transplantation. Drug Discov. Today 2015, 20, 1243–1249. [Google Scholar] [CrossRef]
- Nayman, A.H.; Siginc, H.; Zemheri, E.; Yencilek, F.; Yildirim, A.; Telci, D. Dual-Inhibition of mTOR and Bcl-2 enhances the anti-tumor effect of everolimus against renal cell carcinoma in vitro and in vivo. J. Cancer 2019, 10, 1466–1478. [Google Scholar] [CrossRef]
- Hamieh, L.; Choueiri, T.K.; Ogórek, B.; Khabibullin, D.; Rosebrock, D.; Livitz, D.; Fay, A.; Pignon, J.C.; McDermott, D.F.; Agarwal, N.; et al. Mechanisms of acquired resistance to rapalogs in metastatic renal cell carcinoma. PLoS Genet. 2018, 14, e1007679. [Google Scholar] [CrossRef]
- Notarbartolo, M.; Cervello, M.; Poma, P.; Dusonchet, L.; Meli, M.; D’Alessandro, N. Expression of the IAPs in multidrug resistant tumor cells. Oncol. Rep. 2004, 11, 133–136. [Google Scholar] [CrossRef]
- Notarbartolo, M.; Cervello, M.; Dusonchet, L.; D’Alessandro, N. NAIP-deltaEx10-11: A novel splice variant of the apoptosis inhibitor NAIP differently expressed in drug-sensitive and multidrug-resistant HL60 leukemia cells. Leuk. Res. 2002, 26, 857–862. [Google Scholar] [CrossRef]
- Tamm, I.; Kornblau, S.M.; Segall, H.; Krajewski, S.; Welsh, K.; Kitada, S.; Scudiero, D.A.; Tudor, G.; Qui, Y.H.; Monks, A.; et al. Expression and prognostic significance of IAP-family genes in human cancers and myeloid leukemias. Clin. Cancer Res. 2000, 6, 1796–1803. [Google Scholar]
- Stone, A.; Zukerman, T.; Flaishon, L.; Yakar, R.B.; Rowe, J.M. Efficacy outcomes in the treatment of older or medically unfit patients with acute myeloid leukaemia: A systematic review and meta-analysis. Leuk. Res. 2019, 82, 36–42. [Google Scholar] [CrossRef]
- Rubino, S.; Busà, R.; Attanzio, A.; Alduina, R.; Di Stefano, V.; Girasolo, M.A.; Orecchio, S.; Tesoriere, L. Synthesis, properties, antitumor and antibacterial activity of new Pt(II) and Pd(II) complexes with 2,2′-dithiobis(benzothiazole) ligand. Bioorg. Med. Chem. 2017, 25, 2378–2386. [Google Scholar] [CrossRef]
- Rubino, S.; Pibiri, I.; Minacori, C.; Alduina, R.; Di Stefano, V.; Orecchio, S.; Buscemi, S.; Girasolo, M.A.; Tesoriere, L.; Attanzio, A. Synthesis, structural characterization, anti-proliferative and antimicrobial activity of binuclear and mononuclear Pt(II) complexes with perfluoroalkyl-heterocyclic ligands. Inorg. Chim. Acta 2018, 483, 180–190. [Google Scholar] [CrossRef]
- Ciabocco, M.; Cancemi, P.; Saladino, M.L.; Caponetti, E.; Alduina, R.; Berrettoni, M. Synthesis and antibacterial activity of iron-hexacyanocobaltate nanoparticles. J. Biol. Inorg. Chem. 2018, 23, 385–398. [Google Scholar] [CrossRef]
- Vitale, M.; Galluzzo, P.; Buffa, P.G.; Carlino, E.; Spezia, O.; Alduina, R. Comparison of antibiotic resistance profile and biofilm production of Staphylococcus aureus isolates derived from human specimens and animal-derived samples. Antibiotics 2019, 8, 97. [Google Scholar] [CrossRef]
- Vitale, M.; Gaglio, S.; Galluzzo, P.; Cascone, G.; Piraino, C.; Di Marco Lo Presti, V.; Alduina, R. Antibiotic resistance profiling, analysis of virulence aspects and molecular genotyping of Staphylococcus aureus isolated in Sicily, Italy. Foodborne Pathog. Dis. 2018, 15, 177–185. [Google Scholar] [CrossRef]
- Rauh, W. Succulent and Xerophytic Plants of Madagascar; Strawberry Press: Mill Valley, CA, USA, 1995; Volume I. [Google Scholar]
- Rauh, W. Succulent and Xerophytic Plants of Madagascar; Strawberry Press: Mill Valley, CA, USA, 1998; Volume II. [Google Scholar]
- World Conservation Monitoring Centre. Alluaudia procera. The IUCN Red List of Threatened Species 1998: e.T30436A9550092. Available online: https://www.iucnredlist.org/species/30436/124141236 (accessed on 17 April 2019).
- Zito, P.; Sajeva, M.; Bruno, M.; Maggio, A.; Rosselli, S.; Senatore, F.; Formisano, C. Essential oil composition of the fruits of Periploca laevigata Aiton subsp. angustifolia (Labill.) Markgraf (Apocynaceae—Periplocoideae). Nat. Prod. Res. 2011, 25, 1339–1346. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing: Carol Stream, IL, USA, 2007. [Google Scholar]
- Giardina, A.; Alduina, R.; Gottardi, E.; Di Caro, V.; Süssmuth, R.D.; Puglia, A.M. Differential proteomic analysis highlights metabolic strategies associated with balhimycin production in Amycolatopsis balhimycina chemostat cultivations. Microb. Cell Fact. 2010, 9, 44–54. [Google Scholar] [CrossRef]
- Lo Grasso Maffioli, S.; Sosio, M.; Bibb, M.; Puglia, A.M.; Alduina, R. Two master switch regulators trigger A40926 biosynthesis in Nonomuraea sp. strain ATCC 39727. J. Bacteriol. 2015, 197, 2536–2544. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LRI a | LRI b | Compound | % | Id c | Class d |
|---|---|---|---|---|---|
| 1069 | 1435 | cis-Linalool oxide | 0.2 | 1, 2 | OM |
| 1085 | 1436 | trans-Linalool oxide | 0.1 | 1, 2 | OM |
| 1099 | 1551 | Linalool | 0.2 | 1, 2, 3 | OM |
| 1186 | 1693 | α-Terpineol | 0.3 | 1, 2, 3 | OM |
| 1253 | 1463 | cis-Edulan | 0.3 | 1, 2 | C13 |
| 1272 | 1508 | Vitispirane (isomer not identified) | 3.7 | 1, 2 | C13 |
| 1309 | 1595 | trans-Edulan | 0.9 | 1, 2 | C13 |
| 1345 | 1734 | 1,1,6-Trimethyl-1,2-dihydronaphthalene | 0.2 | 1, 2 | C13 |
| 1380 | 1807 | trans-β-Damascenone | 0.1 | 1, 2 | C13 |
| 1383 | 1804 | 1,4,6-trimethyl-1,2-dihydronaphthalene | 0.2 | 1, 2 | C13 |
| 1411 | 1569 | cis-α-Bergamotene | 0.5 | 1, 2 | SH |
| 1508 | 1735 | (E,E)-α-Farnesene | 0.3 | 1, 2 | SH |
| 1634 | 2032 | 1-epi-Cubenol | 0.3 | 1, 2 | OS |
| 1700 | 1700 | Heptadecane | 0.2 | 1, 2, 3 | H |
| 1738 | 2492 | γ-Costol | 1.0 | 1, 2 | OS |
| 1756 | 2606 | Benzyl benzoate | 0.5 | 1, 2, 3 | O |
| 1800 | 1800 | Octadecane | 0.4 | 1, 2, 3 | H |
| 1838 | 1926 | Neophytadiene (isomer not identified) | 5.6 | 1, 2 | DH |
| 1863 | 1955 | Neophytadiene (isomer not identified) | 1.1 | 1, 2 | DH |
| 1880 | 1982 | Neophytadiene (isomer not identified) | 3.7 | 1, 2 | DH |
| 1900 | 1900 | Nonadecane | 0.9 | 1, 2, 3 | H |
| 1906 | 2418 | Ambrettolide isomer | 7.4 | 1, 2 | ML |
| 1914 | 2385 | Ambrettolide [(Z)-Oxacycloheptadec-8-en-2-one] | 18.1 | 1, 2, 3 | ML |
| 1919 | 2414 | Ambrettolide isomer | 15.2 | 1, 2 | ML |
| 1924 | 2373 | Ambrettolide isomer | 10.8 | 1, 2 | ML |
| 1928 | 2393 | Ambrettolide isomer | 3.8 | 1, 2 | ML |
| 2000 | 2000 | Eicosane | 0.5 | 1, 2, 3 | H |
| 2100 | 2100 | Heneicosane | 0.6 | 1, 2, 3 | H |
| 2111 | 2614 | trans-Phytol | 8.0 | 1, 2, 3 | OD |
| 2114 | 2586 | 14-Methyl-8-hexadecyn-1-ol | 1.2 | 1, 2 | O |
| 2119 | 2575 | Oxacyclononadec-10-en-2-one (isomer not identified) | 1.3 | 1, 2 | ML |
| 2128 | 2610 | Oxacyclononadec-10-en-2-one (isomer not identified) | 2.1 | 1, 2 | ML |
| 2200 | 2200 | Docosane | 0.3 | 1, 2, 3 | H |
| 2300 | 2300 | Tricosane | 0.4 | 1, 2, 3 | H |
| 2400 | 2400 | Tetracosane | 0.1 | 1, 2, 3 | H |
| 2500 | 2500 | Pentacosane | 0.8 | 1, 2, 3 | H |
| 2600 | 2600 | Hexacosane | 0.5 | 1, 2, 3 | H |
| 2700 | 2700 | Heptacosane | 3.1 | 1, 2, 3 | H |
| 2800 | 2800 | Octacosane | 0.4 | 1, 2, 3 | H |
| 2900 | 2900 | Nonacosane | 1.1 | 1, 2, 3 | H |
| Class of Compounds | |||||
| Oxygenated Monoterpene | 0.8 | ||||
| Sesquiterpene Hydrocarbons | 0.8 | ||||
| Oxygenated Sesquiterpene | 1.3 | ||||
| Diterpene Hydrocarbons | 10.4 | ||||
| Oxygenated Diterpene | 8.0 | ||||
| Aliphatic Hydrocarbons | 9.3 | ||||
| C13 Norisoprenoids | 5.4 | ||||
| Macrolactones | 58.7 | ||||
| Others | 1.7 | ||||
| Total | 96.4 |
| Cell Lines and Treatments | Cell Viability (%) | Expected (%) |
|---|---|---|
| HL60 | ||
| Essential oil of A. procera 10 μg/mL | 100.0 ± 0.0 | |
| Essential oil of A. procera 20 μg/mL | 82.0 ± 2.1 | |
| Doxo 1 ng/mL | 90.0 ± 0.7 | |
| Doxo 2 ng/mL | 84.5 ± 3.9 | |
| Essential oil of A. procera 10 μg/mL + Doxo 1ng/mL | 100.0 ± 0.5 | 90.0 ± 0.7 |
| Essential oil of A. procera 10 μg/mL + Doxo 2 ng/mL | 93.0 ± 3.1 | 84.5 ± 3.9 |
| Essential oil of A. procera 20 μg/mL + Doxo 1 ng/mL | 80.0 ± 1.7 | 74.0 ± 1.4 |
| Essential oil of A. procera 20 μg/mL + Doxo 2 ng/mL | 71.0 ± 2.2 | 69.0 ± 1.4 |
| HL60R | ||
| Essential oil of A. procera 10 μg/mL | 100.0 ± 0.0 | |
| Essential oil of A. procera 20 μg/mL | 70.5 ± 3.2 | |
| Doxo 100 ng/mL | 87.5 ± 3.9 | |
| Doxo 500 ng/mL | 76.5 ± 4.6 | |
| Essential oil of A. procera 10 μg/mL + Doxo 100 ng/mL | 95.0 ± 2.2 | 87.5 ± 3.9 |
| Essential oil of A. procera 10 μg/mL + Doxo 500 ng/mL | 88.0 ± 3.8 | 76.5 ± 4.6 |
| Essential oil of A. procera 20 μg/mL + Doxo 100 ng/mL | 70.0 ± 1.9 | 62.0 ± 5.6 |
| Essential oil of A. procera 20 μg/mL + Doxo 500 ng/mL | 61.0 ± 5.9 | 54.0 ± 5.6 |
| Incubation of Bacteria in Presence of | Diameter (cm) of the Inhibition Halo |
|---|---|
| EO | 0.8 ± 0.05 |
| EO + Ery | 0.8 ± 0.03 |
| Ery | 0 ± 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poma, P.; Labbozzetta, M.; Zito, P.; Alduina, R.; Ramarosandratana, A.V.; Bruno, M.; Rosselli, S.; Sajeva, M.; Notarbartolo, M. Essential Oil Composition of Alluaudia procera and in Vitro Biological Activity on Two Drug-Resistant Models. Molecules 2019, 24, 2871. https://doi.org/10.3390/molecules24162871
Poma P, Labbozzetta M, Zito P, Alduina R, Ramarosandratana AV, Bruno M, Rosselli S, Sajeva M, Notarbartolo M. Essential Oil Composition of Alluaudia procera and in Vitro Biological Activity on Two Drug-Resistant Models. Molecules. 2019; 24(16):2871. https://doi.org/10.3390/molecules24162871
Chicago/Turabian StylePoma, Paola, Manuela Labbozzetta, Pietro Zito, Rosa Alduina, Aro Vonjy Ramarosandratana, Maurizio Bruno, Sergio Rosselli, Maurizio Sajeva, and Monica Notarbartolo. 2019. "Essential Oil Composition of Alluaudia procera and in Vitro Biological Activity on Two Drug-Resistant Models" Molecules 24, no. 16: 2871. https://doi.org/10.3390/molecules24162871