
Intensive Distribution of G2-Quaduplexes in the Pseudorabies Virus Genome and Their Sensitivity to Cations and G-Quadruplex Ligands

 and
and
Abstract
:1. Introduction
2. Results
2.1. Bioinformatic Analysis
2.1.1. Genome-Wide Analysis of G3-PQSs and G2-PQSs Distributions in the PRV Genome
2.1.2. Conserved G2-PQSs in the Coding Sequences of PRV Genes
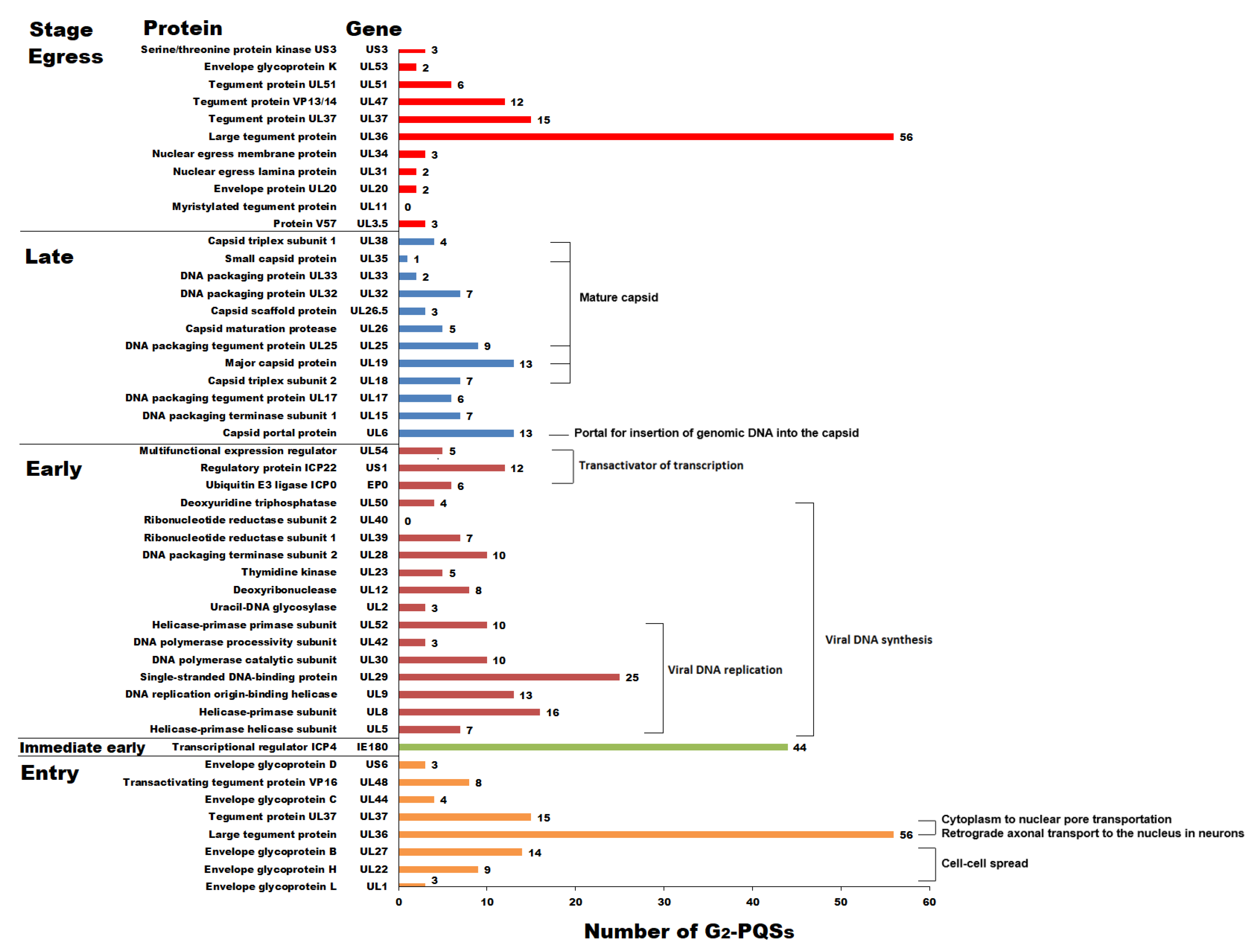
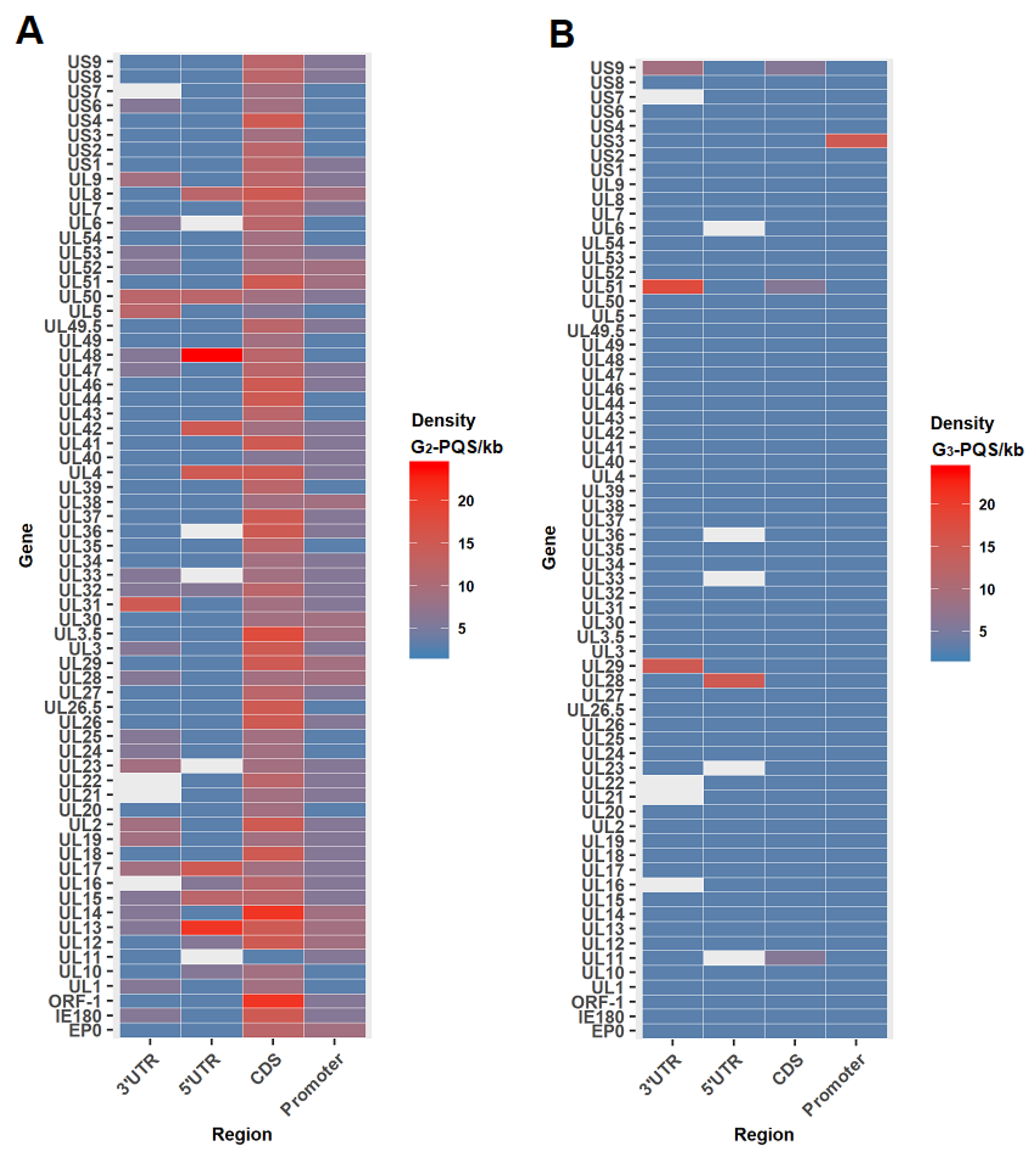
G2-PQS Distribution in the Coding Regions of Genes Involved in the Replication Cycle of PRV
G2-PQSs Involved in the Processes of DNA Replication and Encapsidation
G2-PQSs Involved in the Processes of Virus Entry and Egress
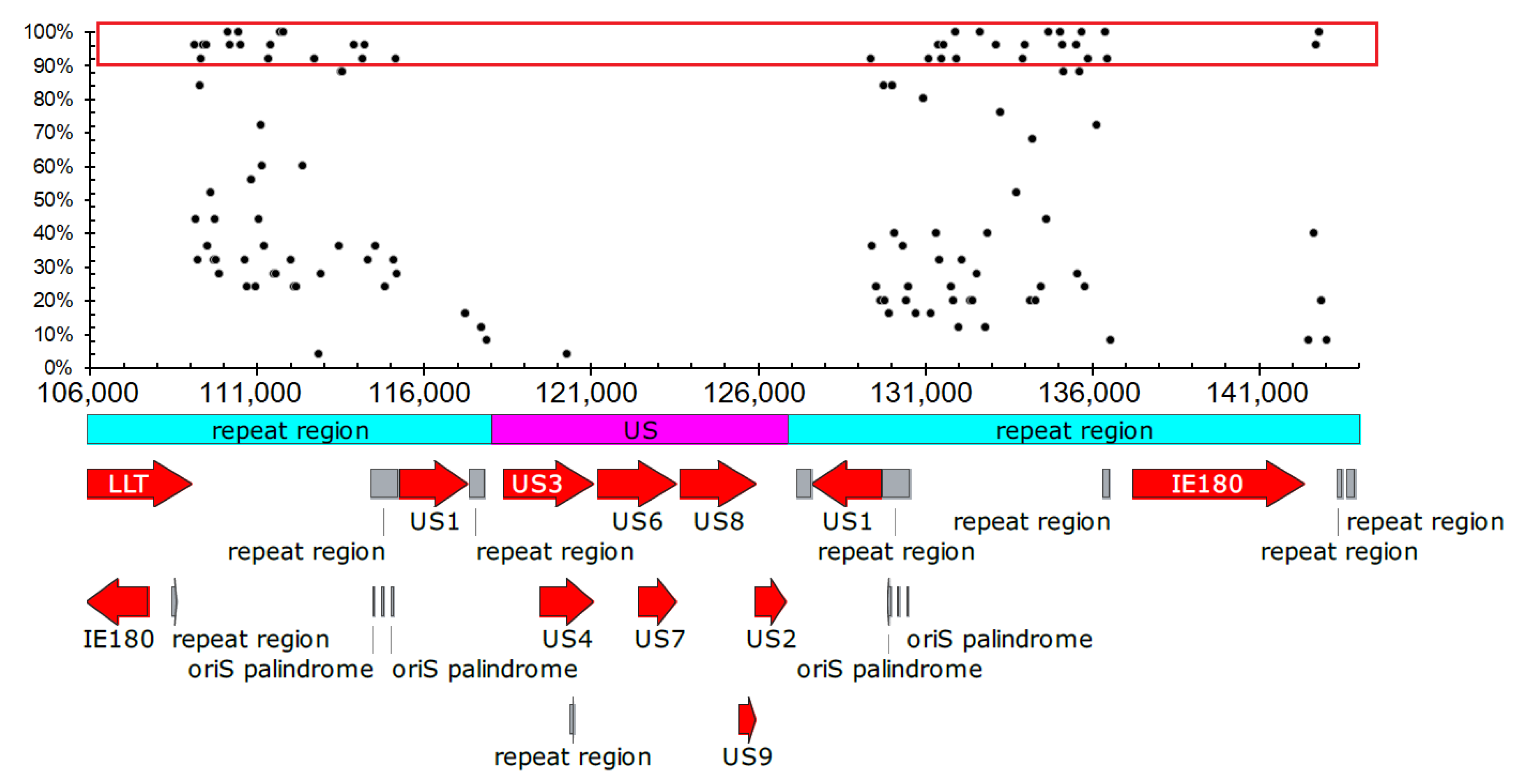
Conservation and Potential Function of G2-PQSs in CDS Region in the Varicellovirus Genus
2.1.3. Distribution Analysis of G2-PQSs in Regulatory Regions in PRV Genomes
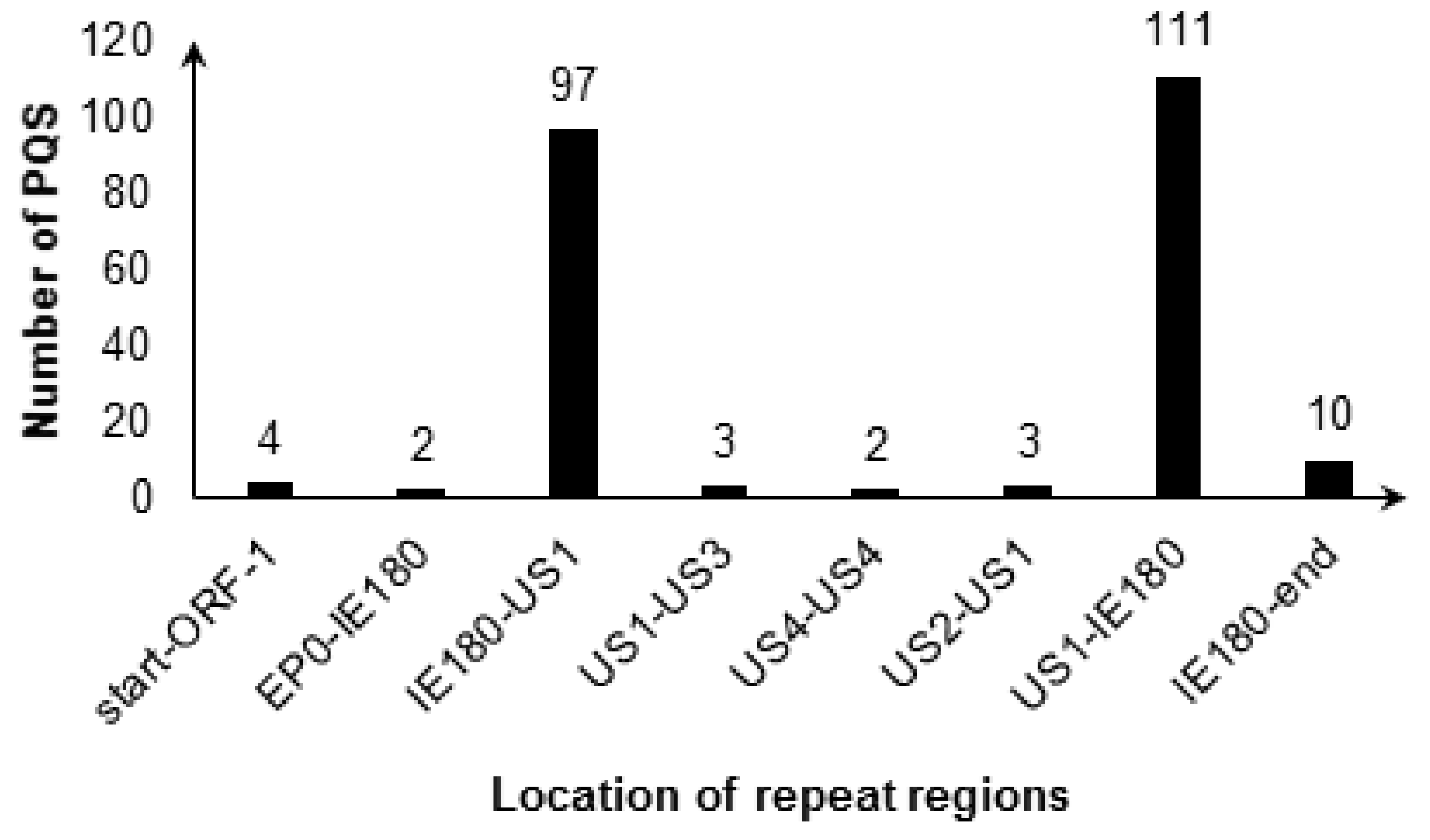
Dozens of Conserved G2-PQSs in the Repeat Regions Related to Genome Recombination
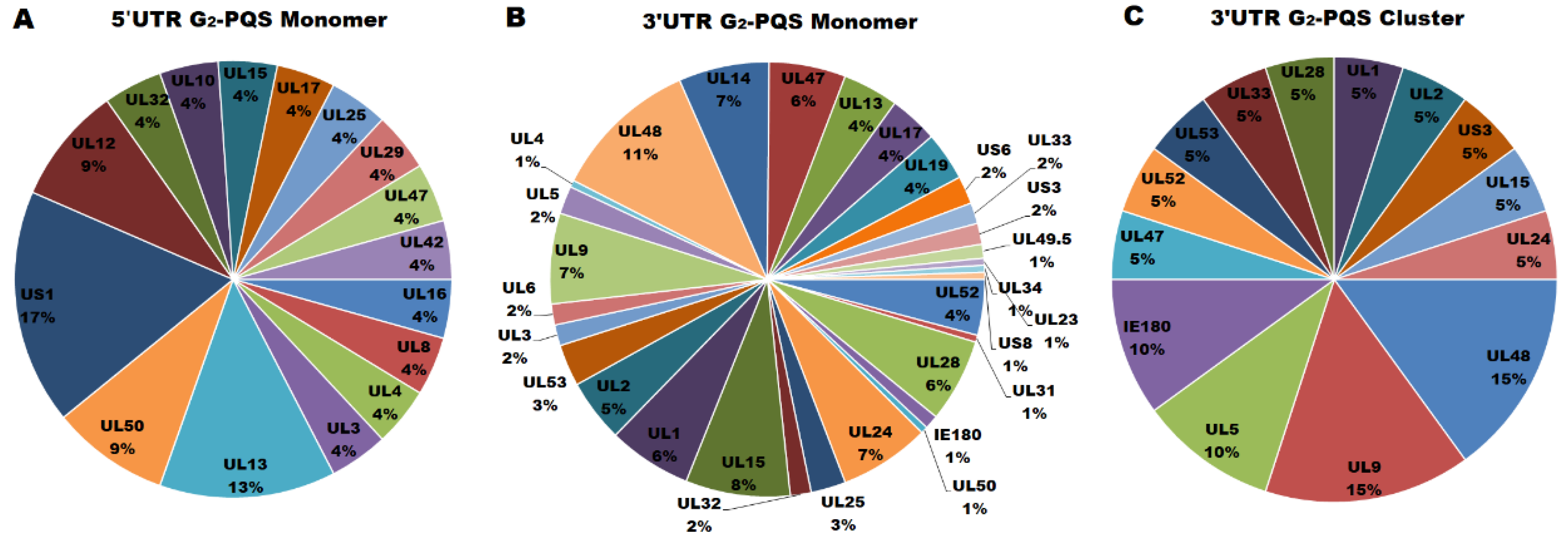
G2-PQSs in the Untranslated Regions of PRV Genes
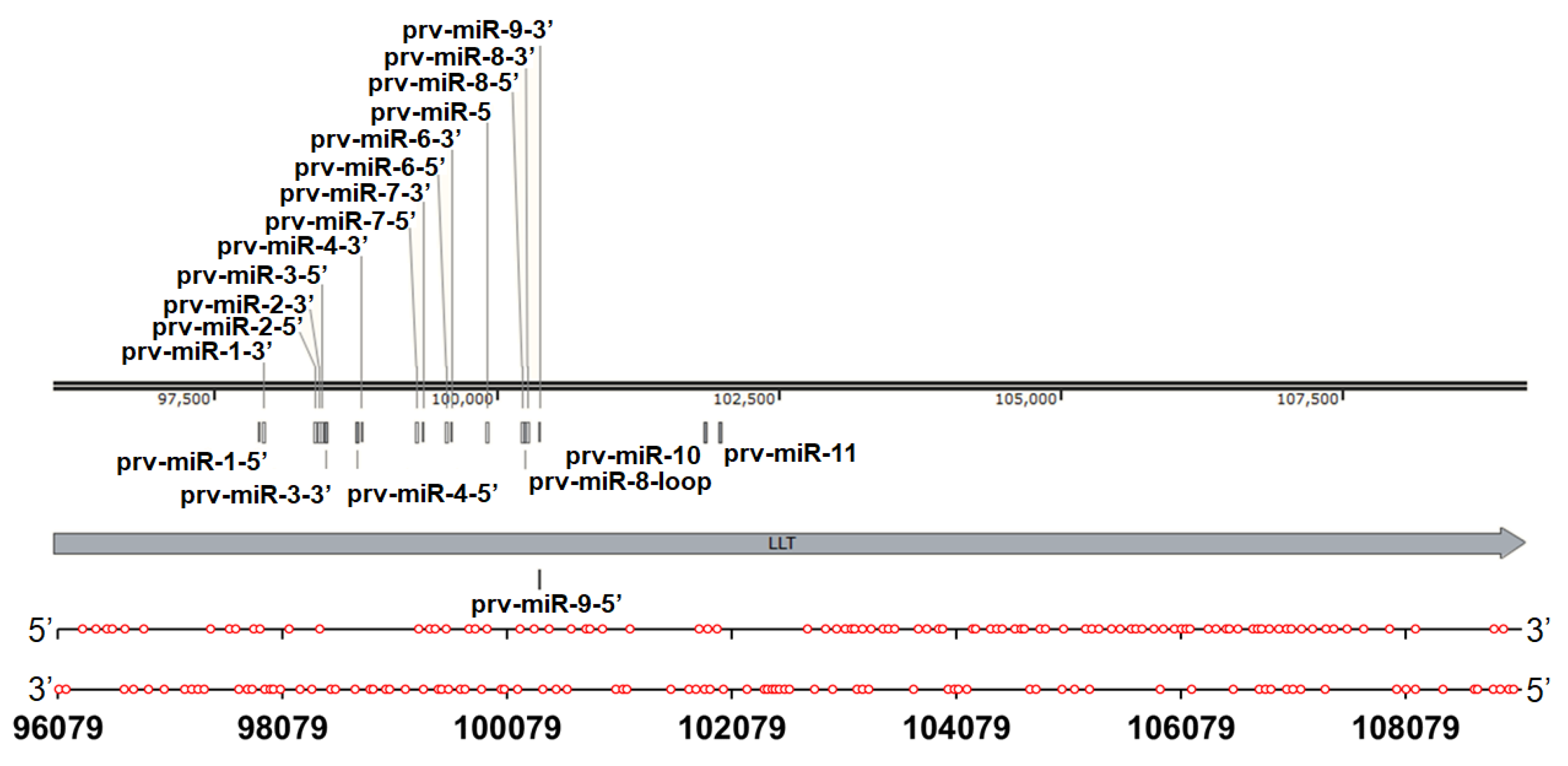
A Wide Distribution of G2-PQSs in the Large Latency Transcript
Density of the G2-PQSs in the Promoter Regions
2.1.4. Comparison of PQS Distribution among Three Herpesvirus Genomes
2.1.5. Summary of Bioinformatics Analysis
2.2. Experimental Validation
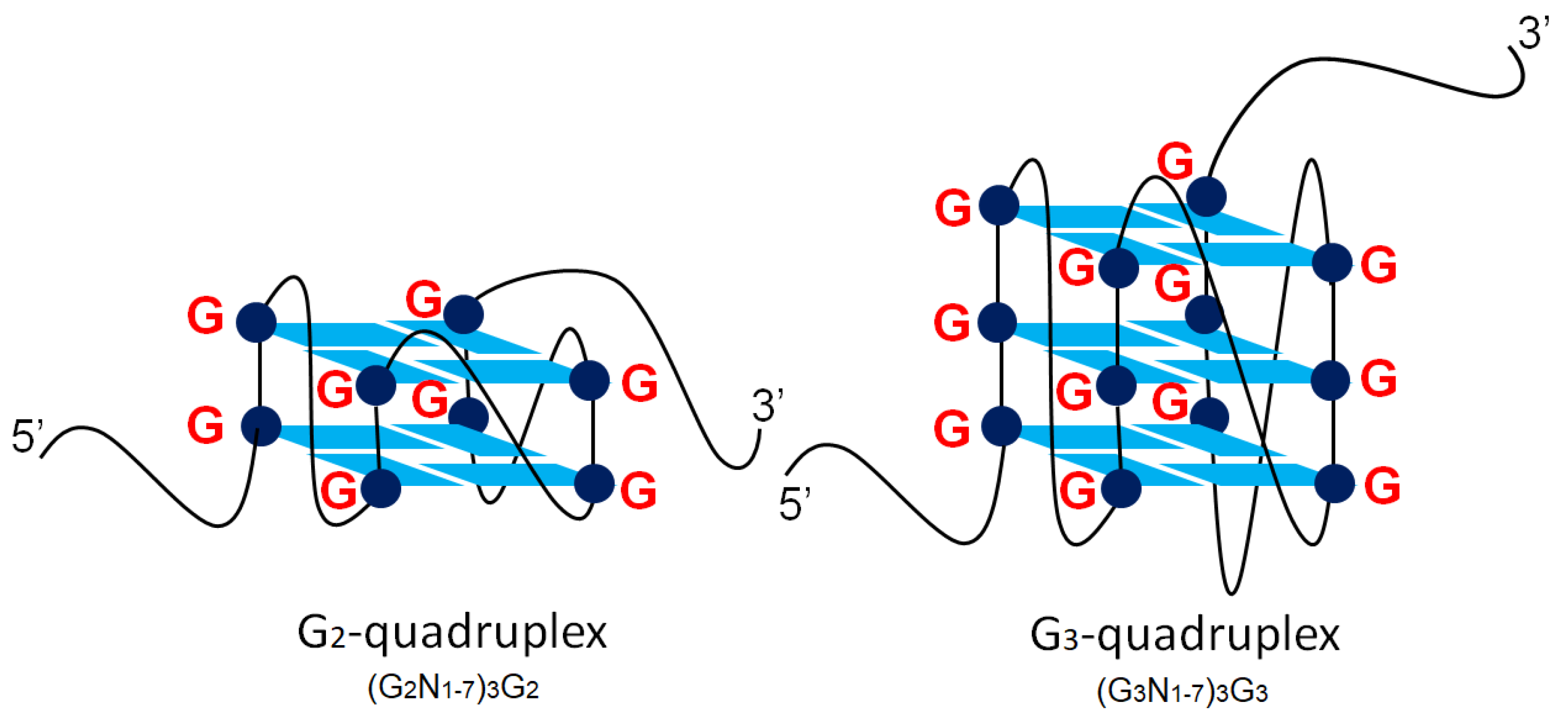
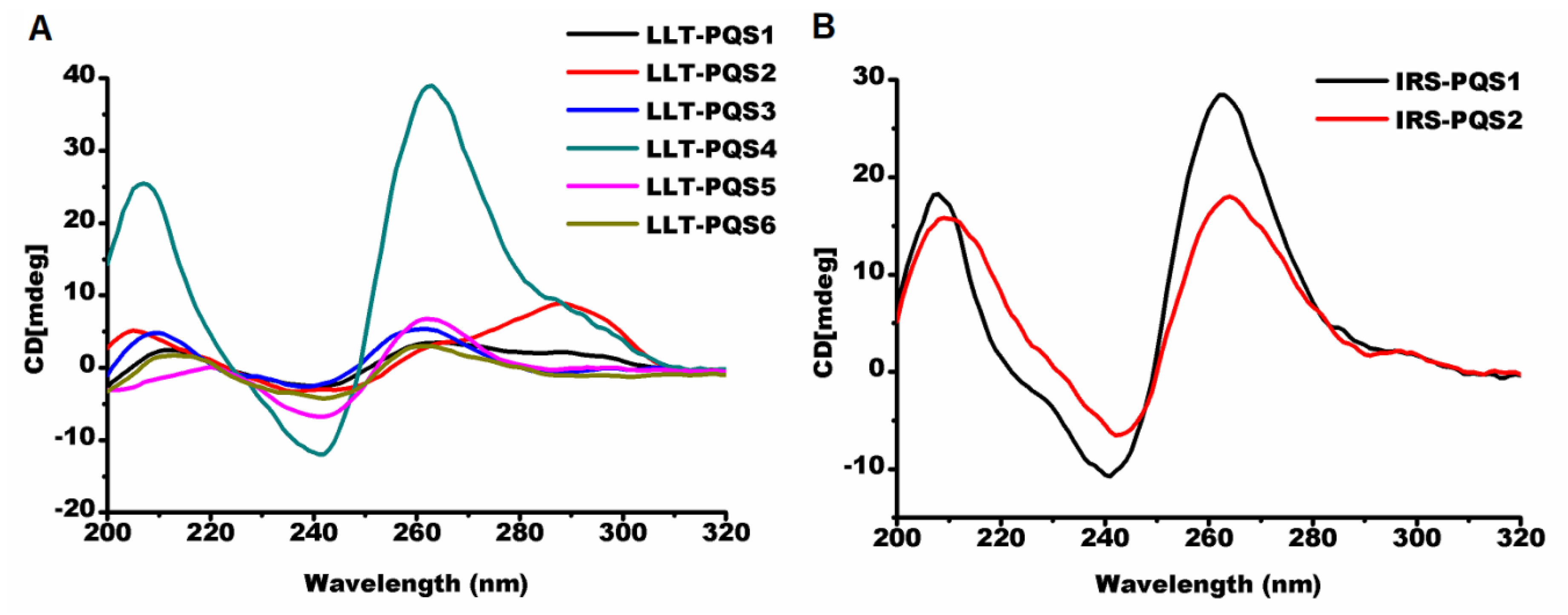
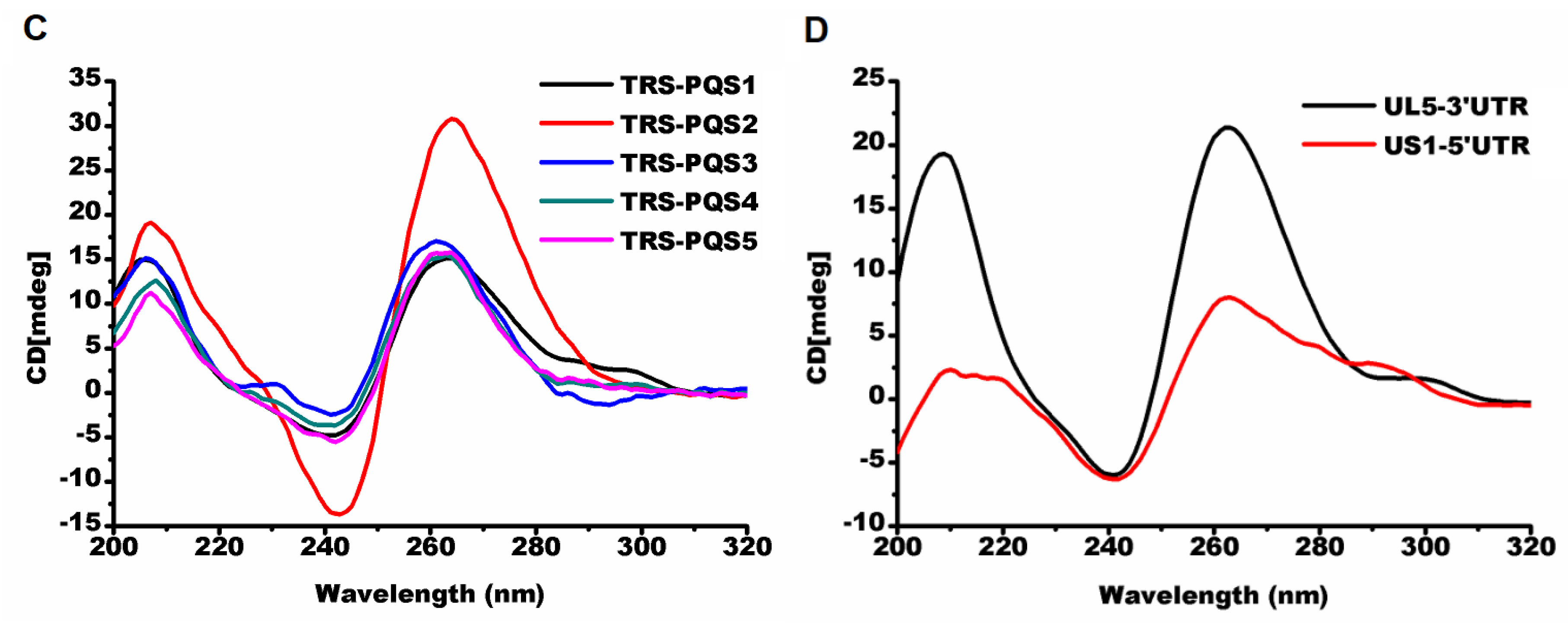
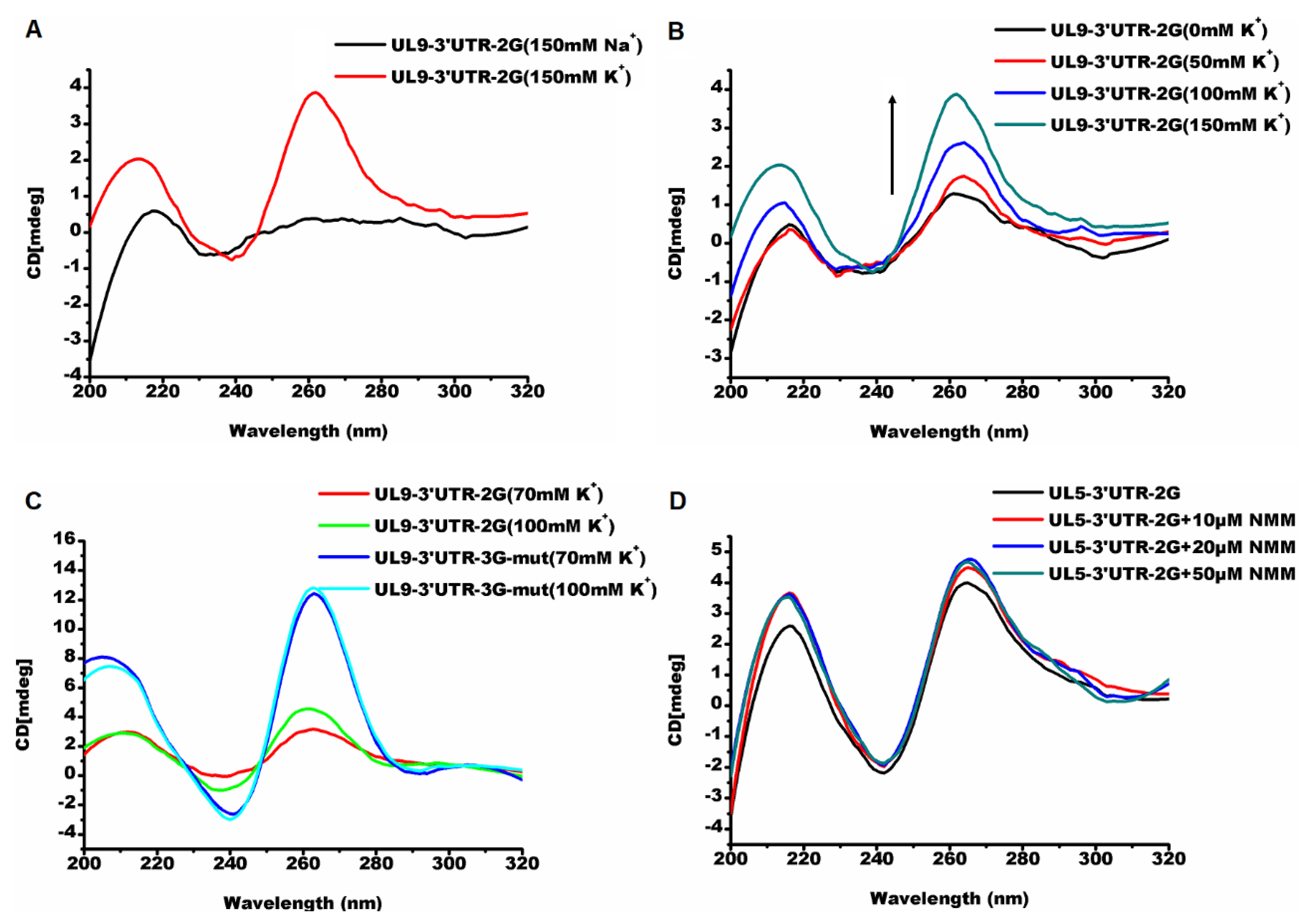
2.2.1. Parallel G-Quadruplexes formed by G2-PQSs In Vitro
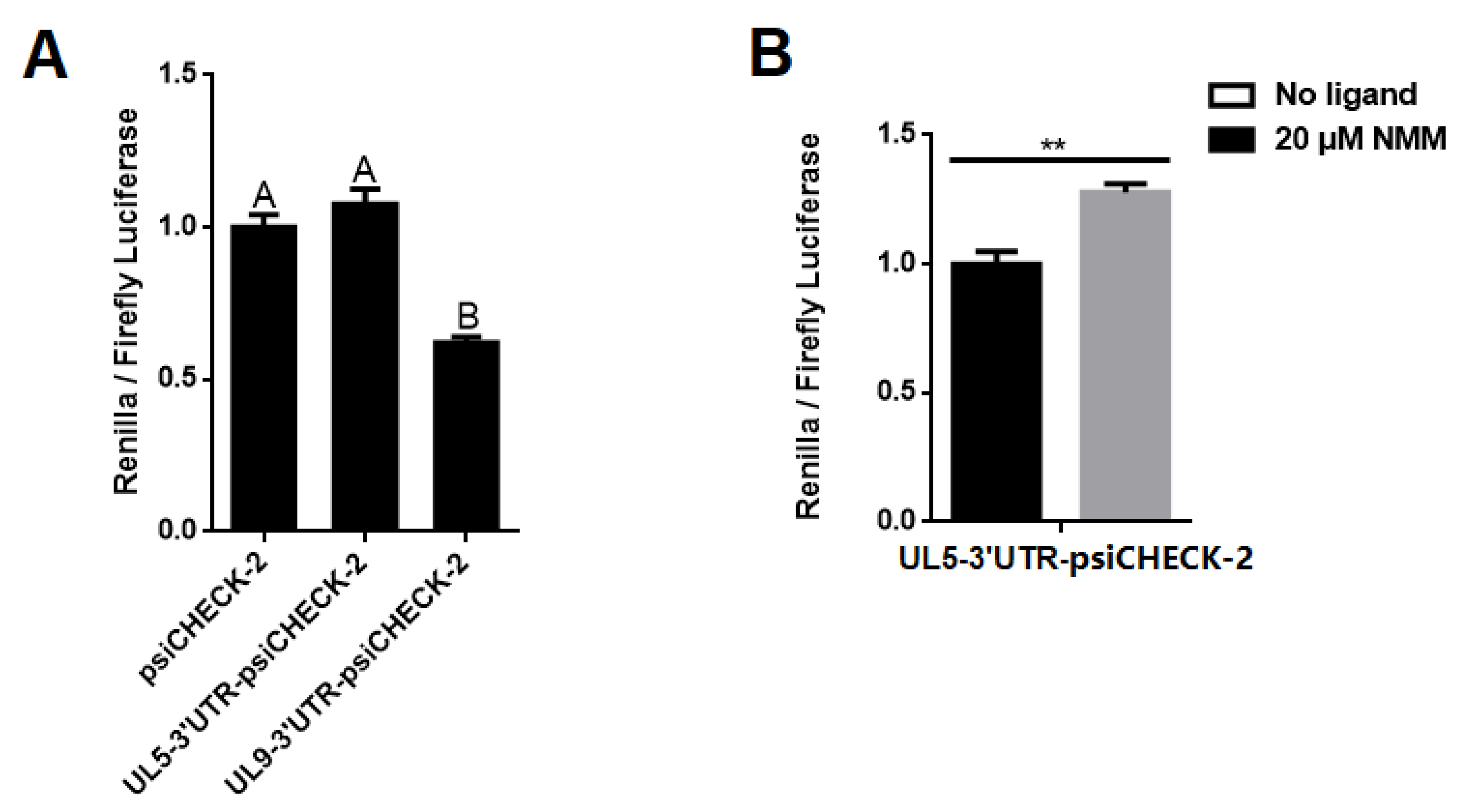
2.2.2. G2-quadruplexes in the 3′ UTR Affects Gene Expression In Vivo with Varying Sensitivities
2.2.3. The G-Quadruplex Ligand Decreases the Virulence of PRV
3. Discussion
4. Methods and Materials
4.1. Virus Sequences
4.2. Identification of Putative G-Quadruplex Sequences in the PRV Genome
4.3. Comparison of the Distribution of G2-PQS and G3-PQS between PRV and Two Herpesviruses
4.4. Conservation of Putative G-Quadruplex Sequences and I-Motif Sequences in the PRV CDS Region in the Varicellovirus Genus
4.5. Percentage Conservation of Putative G-Quadruplex Sequences in the Repeat Regions and LLTs of the PRV Genomes
4.6. Oligonucleotide Folding Conditions
4.7. Circular Dichroism Spectroscopy
4.8. Plasmid Construction
4.9. Cell Culture
4.10. Cytotoxicity Assay
4.11. Transfection and Dual Luciferase Assays
4.12. Plaque Assay
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, K.; Kolb, A.W.; Sverchkov, Y.; Cuellar, J.A.; Craven, M.; Brandt, C.R. Recombination Analysis of Herpes Simplex Virus 1 Reveals a Bias toward GC Content and the Inverted Repeat Regions. J. Virol. 2015, 89, 7214–7223. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, S.; Neidle, S. G-quadruplex nucleic acids as therapeutic targets. Curr. Opin. Chem. Biol. 2009, 13, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Huppert, J.L.; Balasubramanian, S. Prevalence of quadruplexes in the human genome. Nucleic Acids Res. 2005, 33, 2908–2916. [Google Scholar] [CrossRef] [Green Version]
- Andorf, C.M.; Kopylov, M.; Dobbs, D.; Koch, K.E.; Stroupe, M.E.; Lawrence, C.J.; Bass, H.W. G-Quadruplex (G4) Motifs in the Maize (Zea mays L.) Genome Are Enriched at Specific Locations in Thousands of Genes Coupled to Energy Status, Hypoxia, Low Sugar, and Nutrient Deprivation. J. Genet. Genomics 2014, 41, 627–647. [Google Scholar] [CrossRef]
- Biswas, B.; Kandpal, M.; Jauhari, U.K.; Vivekanandan, P. Genome-wide analysis of G-quadruplexes in herpesvirus genomes. BMC Genomics 2016, 17. [Google Scholar] [CrossRef]
- Lavezzo, E.; Berselli, M.; Frasson, I.; Perrone, R.; Palu, G.; Brazzale, A.R.; Richter, S.N.; Toppo, S. G-quadruplex forming sequences in the genome of all known human viruses: A comprehensive guide. PLoS Comput. Biol. 2018, 14, e1006675. [Google Scholar] [CrossRef]
- Murat, P.; Zhong, J.; Lekieffre, L.; Cowieson, N.P.; Clancy, J.L.; Preiss, T.; Balasubramanian, S.; Khanna, R.; Tellam, J. G-quadruplexes regulate Epstein-Barr virus-encoded nuclear antigen 1 mRNA translation. Nat. Chem. Biol. 2014, 10, 358–364. [Google Scholar] [CrossRef]
- Lightfoot, H.L.; Hagen, T.; Clery, A.; Allain, F.H.; Hall, J. Control of the polyamine biosynthesis pathway by G2-quadruplexes. eLife 2018, 7. [Google Scholar] [CrossRef]
- Adler, B.; Sattler, C.; Adler, H. Herpesviruses and Their Host Cells: A Successful Liaison. Trends Microbiol 2017, 25, 229–241. [Google Scholar] [CrossRef]
- Pomeranz, L.E.; Reynolds, A.E.; Hengartner, C.J. Molecular biology of pseudorabies virus: Impact on neurovirology and veterinary medicine. Microbiol Mol. Biol. Rev. 2005, 69, 462–500. [Google Scholar] [CrossRef]
- Muller, T.; Hahn, E.C.; Tottewitz, F.; Kramer, M.; Klupp, B.G.; Mettenleiter, T.C.; Freuling, C. Pseudorabies virus in wild swine: A global perspective. Arch. Virol 2011, 156, 1691–1705. [Google Scholar] [CrossRef]
- Masot, A.J.; Gil, M.; Risco, D.; Jimenez, O.M.; Nunez, J.I.; Redondo, E. Pseudorabies virus infection (Aujeszky’s disease) in an Iberian lynx (Lynx pardinus) in Spain: A case report. BMC Vet. Res. 2017, 13, 6. [Google Scholar] [CrossRef]
- Musante, A.R.; Pedersen, K.; Hall, P. First reports of pseudorabies and winter ticks (Dermacentor albipictus) associated with an emerging feral swine (Sus scrofa) population in New Hampshire. J. Wildlife Dis. 2014, 50, 121–124. [Google Scholar] [CrossRef]
- Pedersen, K.; Bevins, S.N.; Baroch, J.A.; Cumbee, J.C., Jr.; Chandler, S.C.; Woodruff, B.S.; Bigelow, T.T.; DeLiberto, T.J. Pseudorabies in feral swine in the United States, 2009–2012. J. Wildlife Dis. 2013, 49, 709–713. [Google Scholar] [CrossRef]
- An, T.Q.; Peng, J.M.; Tian, Z.J.; Zhao, H.Y.; Li, N.; Liu, Y.M.; Chen, J.Z.; Leng, C.L.; Sun, Y.; Chang, D.; et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs, China, 2012. Emerg. Infect. Dis. 2013, 19, 1749–1755. [Google Scholar] [CrossRef]
- Hu, H.; Yu, T.; Fan, J.; Cheng, S.; Liu, C.; Deng, F.; Guo, N.; Wu, B.; He, Q.G. Genetic Properties of the New Pseudorabies Virus Isolates in China in 2012. Pak. Vet. J. 2016, 36, 264–269. [Google Scholar]
- Liu, H.; Li, X.T.; Hu, B.; Deng, X.Y.; Zhang, L.; Lian, S.Z.; Zhang, H.L.; Lv, S.; Xue, X.H.; Lu, R.G.; et al. Outbreak of severe pseudorabies virus infection in pig-offal-fed farmed mink in Liaoning Province, China. Arch. Virol 2017, 162, 863–866. [Google Scholar] [CrossRef]
- Wang, X.; Wu, C.X.; Song, X.R.; Chen, H.C.; Liu, Z.F. Comparison of pseudorabies virus China reference strain with emerging variants reveals independent virus evolution within specific geographic regions. Virology 2017, 506, 92–98. [Google Scholar] [CrossRef]
- Wu, R.; Bai, C.; Sun, J.; Chang, S.; Zhang, X. Emergence of virulent pseudorabies virus infection in northern China. J. Vet. Sci. 2013, 14, 363–365. [Google Scholar] [CrossRef]
- Yu, T.; Chen, F.; Ku, X.; Fan, J.; Zhu, Y.; Ma, H.; Li, S.; Wu, B.; He, Q. Growth characteristics and complete genomic sequence analysis of a novel pseudorabies virus in China. Virus Genes 2016, 52, 474–483. [Google Scholar] [CrossRef]
- Yu, T.; Chen, F.; Ku, X.; Zhu, Y.; Ma, H.; Li, S.; He, Q. Complete Genome Sequence of Novel Pseudorabies Virus Strain HNB Isolated in China. Genome. Announc. 2016, 4. [Google Scholar] [CrossRef]
- Yu, X.; Zhou, Z.; Hu, D.; Zhang, Q.; Han, T.; Li, X.; Gu, X.; Yuan, L.; Zhang, S.; Wang, B.; et al. Pathogenic pseudorabies virus, China, 2012. Emerg. Infect. Dis. 2014, 20, 102–104. [Google Scholar] [CrossRef]
- Dong, B.; Zarlenga, D.S.; Ren, X. An overview of live attenuated recombinant pseudorabies viruses for use as novel vaccines. J. Immunol. Res. 2014, 2014, 824630. [Google Scholar] [CrossRef]
- The National Center for Biotechnology Information Genome Home Page. Available online: https://www.ncbi.nlm.nih.gov/genome (accessed on 13 August 2018).
- Artusi, S.; Nadai, M.; Perrone, R.; Biasolo, M.A.; Palu, G.; Flamand, L.; Calistri, A.; Richter, S.N. The Herpes Simplex Virus-1 genome contains multiple clusters of repeated G-quadruplex: Implications for the antiviral activity of a G-quadruplex ligand. Antiviral. Res. 2015, 118, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Depledge, D.P.; Ouwendijk, W.J.D.; Sadaoka, T.; Braspenning, S.E.; Mori, Y.; Cohrs, R.J.; Verjans, G.; Breuer, J. A spliced latency-associated VZV transcript maps antisense to the viral transactivator gene 61. Nat. Commun 2018, 9, 1167. [Google Scholar] [CrossRef] [Green Version]
- Granzow, H.; Klupp, B.G.; Mettenleiter, T.C. Entry of pseudorabies virus: An immunogold-labeling study. J. Virol. 2005, 79, 3200–3205. [Google Scholar] [CrossRef]
- Madireddy, A.; Purushothaman, P.; Loosbroock, C.P.; Robertson, E.S.; Schildkraut, C.L.; Verma, S.C. G-quadruplex-interacting compounds alter latent DNA replication and episomal persistence of KSHV. Nucleic Acids Res. 2016, 44, 3675–3694. [Google Scholar] [CrossRef] [Green Version]
- Bugaut, A.; Balasubramanian, S. 5’-UTR RNA G-quadruplexes: Translation regulation and targeting. Nucleic Acids Res. 2012, 40, 4727–4741. [Google Scholar] [CrossRef]
- Kumari, S.; Bugaut, A.; Huppert, J.L.; Balasubramanian, S. An RNA G-quadruplex in the 5’ UTR of the NRAS proto-oncogene modulates translation. Nat. Chem. Biol. 2007, 3, 218–221. [Google Scholar] [CrossRef]
- Subramanian, M.; Rage, F.; Tabet, R.; Flatter, E.; Mandel, J.L.; Moine, H. G-quadruplex RNA structure as a signal for neurite mRNA targeting. EMBO Rep. 2011, 12, 697–704. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.M.; Yan, K.; Tang, Q.; Wu, Y.Q.; He, W.B.; Chen, H.C.; Liu, Z.F. The full-length microRNA cluster in the intron of large latency transcript is associated with the virulence of pseudorabies virus. Virology 2018, 520, 59–66. [Google Scholar] [CrossRef]
- Mahjoub, N.; Dhorne-Pollet, S.; Fuchs, W.; Endale Ahanda, M.L.; Lange, E.; Klupp, B.; Arya, A.; Loveland, J.E.; Lefevre, F.; Mettenleiter, T.C.; et al. A 2.5-kilobase deletion containing a cluster of nine microRNAs in the latency-associated-transcript locus of the pseudorabies virus affects the host response of porcine trigeminal ganglia during established latency. J. Virol. 2015, 89, 428–442. [Google Scholar] [CrossRef]
- Amrane, S.; Kerkour, A.; Bedrat, A.; Vialet, B.; Andreola, M.L.; Mergny, J.L. Topology of a DNA G-quadruplex structure formed in the HIV-1 promoter: A potential target for anti-HIV drug development. J. Am. Chem. Soc. 2014, 136, 5249–5252. [Google Scholar] [CrossRef]
- Perrone, R.; Nadai, M.; Frasson, I.; Poe, J.A.; Butovskaya, E.; Smithgall, T.E.; Palumbo, M.; Palu, G.; Richter, S.N. A dynamic G-quadruplex region regulates the HIV-1 long terminal repeat promoter. J. Med. Chem. 2013, 56, 6521–6530. [Google Scholar] [CrossRef]
- Kong, J.N.; Zhang, C.; Zhu, Y.C.; Zhong, K.; Wang, J.; Chu, B.B.; Yang, G.Y. Identification and characterization of G-quadruplex formation within the EP0 promoter of pseudorabies virus. Sci. Rep. 2018, 8, 14029. [Google Scholar] [CrossRef]
- Depledge, D.P.; Sadaoka, T.; Ouwendijk, W.J.D. Molecular aspects of Varicella-Zoster Virus latency. Viruses 2018, 10. [Google Scholar] [CrossRef]
- Kypr, J.; Kejnovska, I.; Renciuk, D.; Vorlickova, M. Circular dichroism and conformational polymorphism of DNA. Nucleic Acids Res. 2009, 37, 1713–1725. [Google Scholar] [CrossRef] [Green Version]
- Beaudoin, J.D.; Perreault, J.P. Exploring mRNA 3’-UTR G-quadruplexes: Evidence of roles in both alternative polyadenylation and mRNA shortening. Nucleic Acids Res. 2013, 41, 5898–5911. [Google Scholar] [CrossRef]
- Sahakyan, A.B.; Murat, P.; Mayer, C.; Balasubramanian, S. G-quadruplex structures within the 3’ UTR of LINE-1 elements stimulate retrotransposition. Nat. Struct. Mol. Biol. 2017, 24, 243–247. [Google Scholar] [CrossRef]
- Kim, M.Y.; Gleason-Guzman, M.; Izbicka, E.; Nishioka, D.; Hurley, L.H. The different biological effects of telomestatin and TMPyP4 can be attributed to their selectivity for interaction with intramolecular or intermolecular G-quadruplex structures. Cancer Res. 2003, 63, 3247–3256. [Google Scholar]
- Grand, C.L.; Han, H.; Munoz, R.M.; Weitman, S.; Von Hoff, D.D.; Hurley, L.H.; Bearss, D.J. The cationic porphyrin TMPyP4 down-regulates c-MYC and human telomerase reverse transcriptase expression and inhibits tumor growth in vivo. Mol. Cancer Ther. 2002, 1, 565–573. [Google Scholar]
- Mikami-Terao, Y.; Akiyama, M.; Yuza, Y.; Yanagisawa, T.; Yamada, O.; Yamada, H. Antitumor activity of G-quadruplex-interactive agent TMPyP4 in K562 leukemic cells. Cancer Lett. 2008, 261, 226–234. [Google Scholar] [CrossRef]
- Varizhuk, A.M.; Protopopova, A.D.; Tsvetkov, V.B.; Barinov, N.A.; Podgorsky, V.V.; Tankevich, M.V.; Vlasenok, M.A.; Severov, V.V.; Smirnov, I.P.; Dubrovin, E.V.; et al. Polymorphism of G4 associates: From stacks to wires via interlocks. Nucleic Acids Res. 2018, 46, 8978–8992. [Google Scholar] [CrossRef]
- Beaudoin, J.D.; Perreault, J.P. 5’-UTR G-quadruplex structures acting as translational repressors. Nucleic Acids Res. 2010, 38, 7022–7036. [Google Scholar] [CrossRef]
- Rouleau, S.; Glouzon, J.S.; Brumwell, A.; Bisaillon, M.; Perreault, J.P. 3’ UTR G-quadruplexes regulate miRNA binding. RNA 2017, 23, 1172–1179. [Google Scholar] [CrossRef]
- Bolduc, F.; Garant, J.M.; Allard, F.; Perreault, J.P. Irregular G-quadruplexes Found in the Untranslated Regions of Human mRNAs Influence Translation. J. Biol. Chem. 2016, 291, 21751–21760. [Google Scholar] [CrossRef] [Green Version]
- Greco, A.; Fester, N.; Engel, A.T.; Kaufer, B.B. Role of the short telomeric repeat region in Marek’s disease virus replication, genomic integration, and lymphomagenesis. J. Virol. 2014, 88, 14138–14147. [Google Scholar] [CrossRef]
- Lieberman, P.M. Epigenetics and Genetics of Viral Latency. Cell Host Microbe 2016, 19, 619–628. [Google Scholar] [CrossRef]
- Gilbert-Girard, S.; Gravel, A.; Artusi, S.; Richter, S.N.; Wallaschek, N.; Kaufer, B.B.; Flamand, L. Stabilization of Telomere G-Quadruplexes Interferes with Human Herpesvirus 6A Chromosomal Integration. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Sun, D.; Thompson, B.; Cathers, B.E.; Salazar, M.; Kerwin, S.M.; Trent, J.O.; Jenkins, T.C.; Neidle, S.; Hurley, L.H. Inhibition of human telomerase by a G-quadruplex-interactive compound. J. Med. Chem. 1997, 40, 2113–2116. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A flexible suite of utilities for comparing genomic features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Nicoludis, J.M.; Miller, S.T.; Jeffrey, P.D.; Barrett, S.P.; Rablen, P.R.; Lawton, T.J.; Yatsunyk, L.A. Optimized end-stacking provides specificity of N-methyl mesoporphyrin IX for human telomeric G-quadruplex DNA. J. Am. Chem. Soc. 2012, 134, 20446–20456. [Google Scholar] [CrossRef]
- Campbell, N.H.; Parkinson, G.N.; Reszka, A.P.; Neidle, S. Structural basis of DNA quadruplex recognition by an acridine drug. J. Am. Chem. Soc. 2008, 130, 6722–6724. [Google Scholar] [CrossRef]
- Koirala, D.; Dhakal, S.; Ashbridge, B.; Sannohe, Y.; Rodriguez, R.; Sugiyama, H.; Balasubramanian, S.; Mao, H. A single-molecule platform for investigation of interactions between G-quadruplexes and small-molecule ligands. Nat. Chem. 2011, 3, 782–787. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Herpesviruses | Region † | Length (bp) | Putative G2-quadruplex Sequences | Putative G3-quadruplex Sequences | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Number | Monomer | Cluster | Density | Number | Monomer | Cluster | Density | |||
| (PQS/kb) | (PQS/kb) | |||||||||
| Pseudorabies virus (PRV, NC_006151.1) | coding sequence (CDS) | 105,363 | 1126 | 1004 | 122 | 10.69 | 51 | 51 | 0 | 0.48 |
| 3’ end untranslated region (3′ UTR) | 35,688 | 166 | 149 | 17 | 4.65 | 6 | 6 | 0 | 0.17 | |
| 5’ end untranslated region (5′ UTR) | 6760 | 19 | 18 | 1 | 2.81 | 5 | 3 | 2 | 0.74 | |
| Promoter | 56,324 | 301 | 276 | 25 | 5.34 | 36 | 32 | 4 | 0.64 | |
| large latency transcript (LLT) | 13,040 | 139 | 109 | 30 | 10.66 | 27 | 25 | 2 | 2.07 | |
| Repeat region | 36,234 | 338 | 269 | 69 | 9.33 | 131 | 109 | 22 | 3.62 | |
| Human alphaherpesvirus 1 (Herpes simplex virus type 1, HSV-1, NC_001806.2) | CDS | 121,089 | 1351 | 1248 | 103 | 11.16 | 109 | 104 | 5 | 0.90 |
| 3′ UTR | 49,322 | 235 | 223 | 12 | 4.76 | 27 | 27 | 0 | 0.55 | |
| 5′ UTR | 3434 | 18 | 18 | 0 | 5.24 | 3 | 1 | 2 | 0.87 | |
| Promoter | 64,966 | 346 | 317 | 29 | 5.33 | 64 | 56 | 8 | 0.99 | |
| latency-associated transcript (LAT) | 15,944 | 158 | 146 | 12 | 9.91 | 54 | 44 | 10 | 3.39 | |
| Repeat region | 31,875 | 338 | 307 | 31 | 10.60 | 120 | 87 | 33 | 3.76 | |
| Human alphaherpesvirus 3 (Varicella-zoster virus, VZV, NC_001348.1) | CDS | 111,496 | 296 | 282 | 14 | 2.65 | 10 | 10 | 0 | 0.09 |
| 3′ UTR | 29,758 | 37 | 36 | 1 | 1.24 | 1 | 1 | 0 | 0.03 | |
| 5′ UTR * | 496 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Promoter | 60,900 | 71 | 69 | 2 | 1.17 | 5 | 3 | 2 | 0.08 | |
| VZV latency-associated transcript (VLT) | 2417 | 13 | 12 | 1 | 5.38 | 0 | 0 | 0 | 0 | |
| Repeat region | 15,514 | 119 | 111 | 8 | 7.67 | 11 | 9 | 2 | 0.71 | |
| Gene Common Name | Function | Putative G2-quadruplex Sequence (5′-3′) | Score | Nucleotide Identity |
|---|---|---|---|---|
| IE180 | Transactivator, homolog of ICP4 | GGGCCGGGAACTGGACCGGG | 1.000 | 0.579 |
| GGACTGGCCCGCGGACGGCCCGGCCGTGGGGG | 0.900 | 0.579 | ||
| GGCTCGGCGCGGCGCGGCGCCGG | 0.800 | 0.579 | ||
| GGCCAACGTGGCCGCGGCCCGG | 0.700 | 0.579 | ||
| GGGCCCCGGTCCCGGGCCGGCTCCGGGCCCCGG | 0.700 | 0.579 | ||
| UL30 | DNA replication | GGACGACGGCGGCGGCTACCAGGGCGCCAAGG | 0.727 | 0.678 |
| GGTGTACGGGTTCACGGGCGTGGCCAACGGG | 0.727 | 0.678 | ||
| UL33 | DNA cleavage and packaging | GGGGAGGCGCTGCGGGCGCGG | 1.000 | 0.664 |
| UL17 | DNA cleavage and encapsidation | GGGGCGGCCGGCGCGGGCCCCGG | 0.778 | 0.589 |
| UL16 | Tegument protein | GGTCCTGGCCCCCGGCGCGTGGTGGGCGCGCGG | 0.800 | 0.580 |
| UL47 | Tegument protein | GGGGACGAGGAGGAGGAGGAGGAGGAGGAGGAGAGCGAGGGGGGCGCGTGGTCCGACGGGG | 0.700 | 0.486 |
| UL13 | Protein-serine/threonine kinase | GGCCGTCGGGGCCGGATCGTACGG | 1.000 | 0.525 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, H.; Gong, B.; Yang, Z.; Li, Z.; Zhou, H.; Zhang, Y.; Niu, X.; Liu, S.; Wei, D. Intensive Distribution of G2-Quaduplexes in the Pseudorabies Virus Genome and Their Sensitivity to Cations and G-Quadruplex Ligands. Molecules 2019, 24, 774. https://doi.org/10.3390/molecules24040774
Deng H, Gong B, Yang Z, Li Z, Zhou H, Zhang Y, Niu X, Liu S, Wei D. Intensive Distribution of G2-Quaduplexes in the Pseudorabies Virus Genome and Their Sensitivity to Cations and G-Quadruplex Ligands. Molecules. 2019; 24(4):774. https://doi.org/10.3390/molecules24040774
Chicago/Turabian StyleDeng, Hui, Bowen Gong, Zhiquan Yang, Zhen Li, Huan Zhou, Yashu Zhang, Xiaohui Niu, Sisi Liu, and Dengguo Wei. 2019. "Intensive Distribution of G2-Quaduplexes in the Pseudorabies Virus Genome and Their Sensitivity to Cations and G-Quadruplex Ligands" Molecules 24, no. 4: 774. https://doi.org/10.3390/molecules24040774