
Schisandra Chinensis Lignans Suppresses the Production of Inflammatory Mediators Regulated by NF-κB, AP-1, and IRF3 in Lipopolysaccharide-Stimulated RAW264.7 Cells

Abstract
:1. Introduction
2. Results
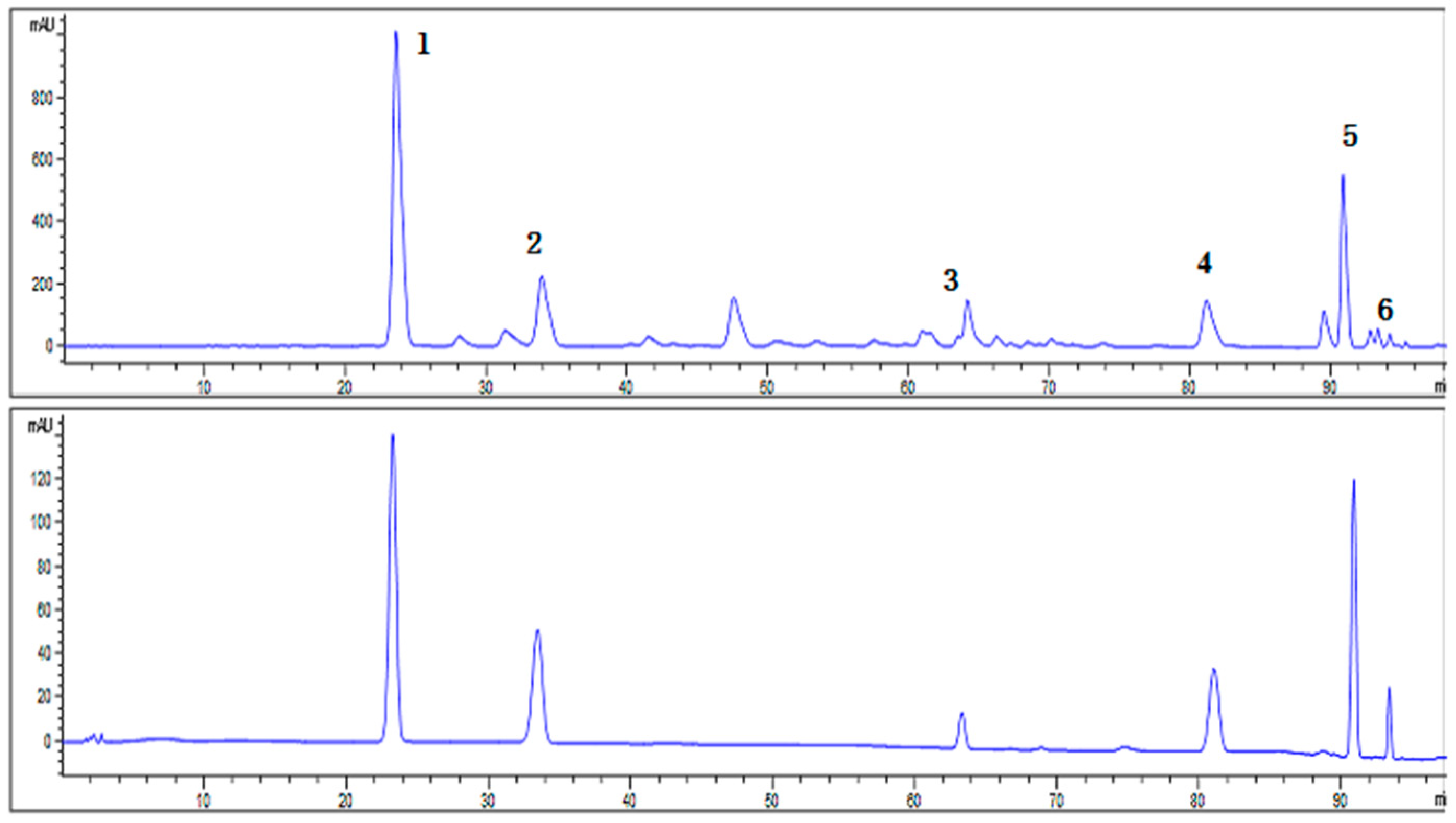
2.1. Characterization of SCL
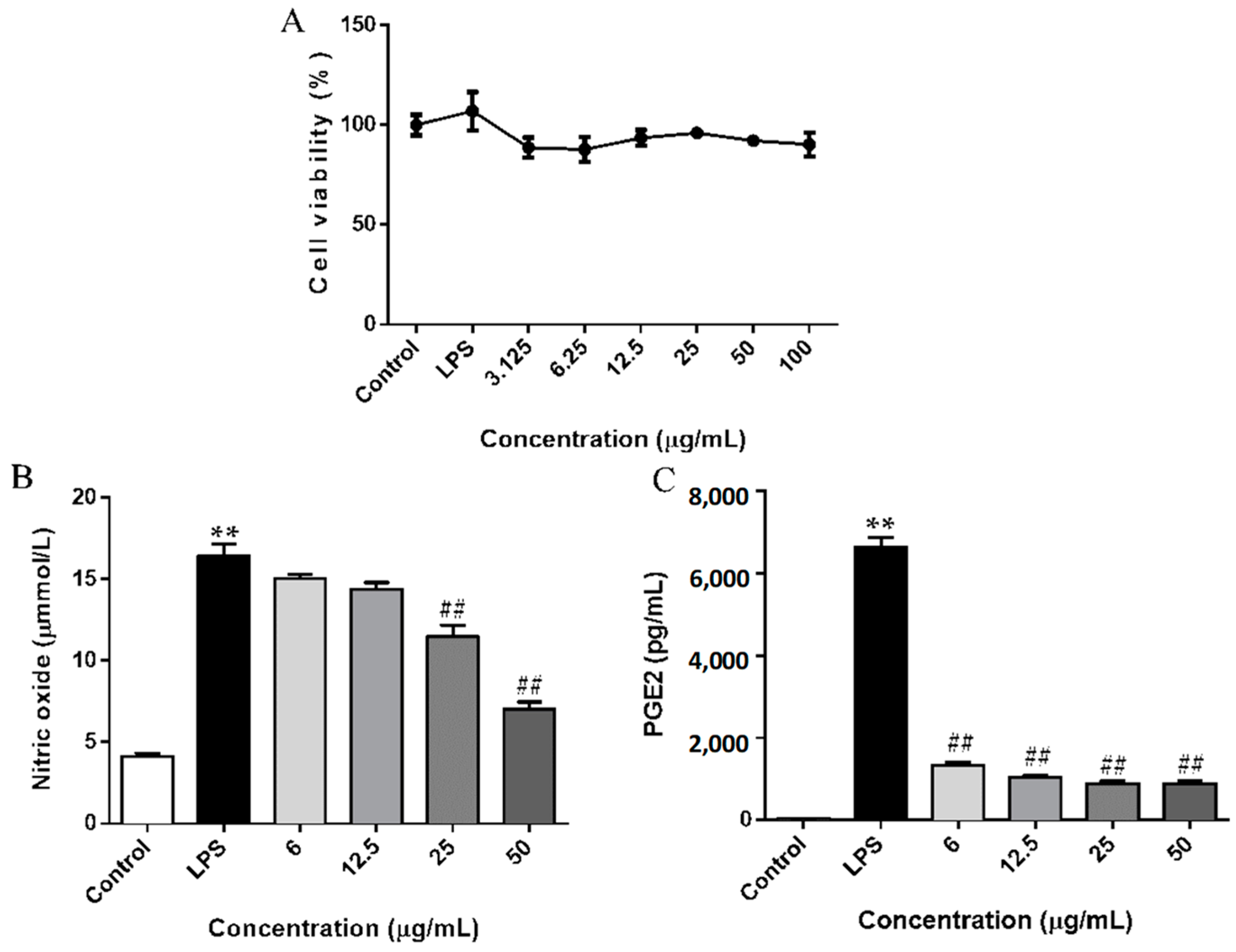
2.2. SCL Decreased the Release of NO and PGE2 in RAW264.7 Cells
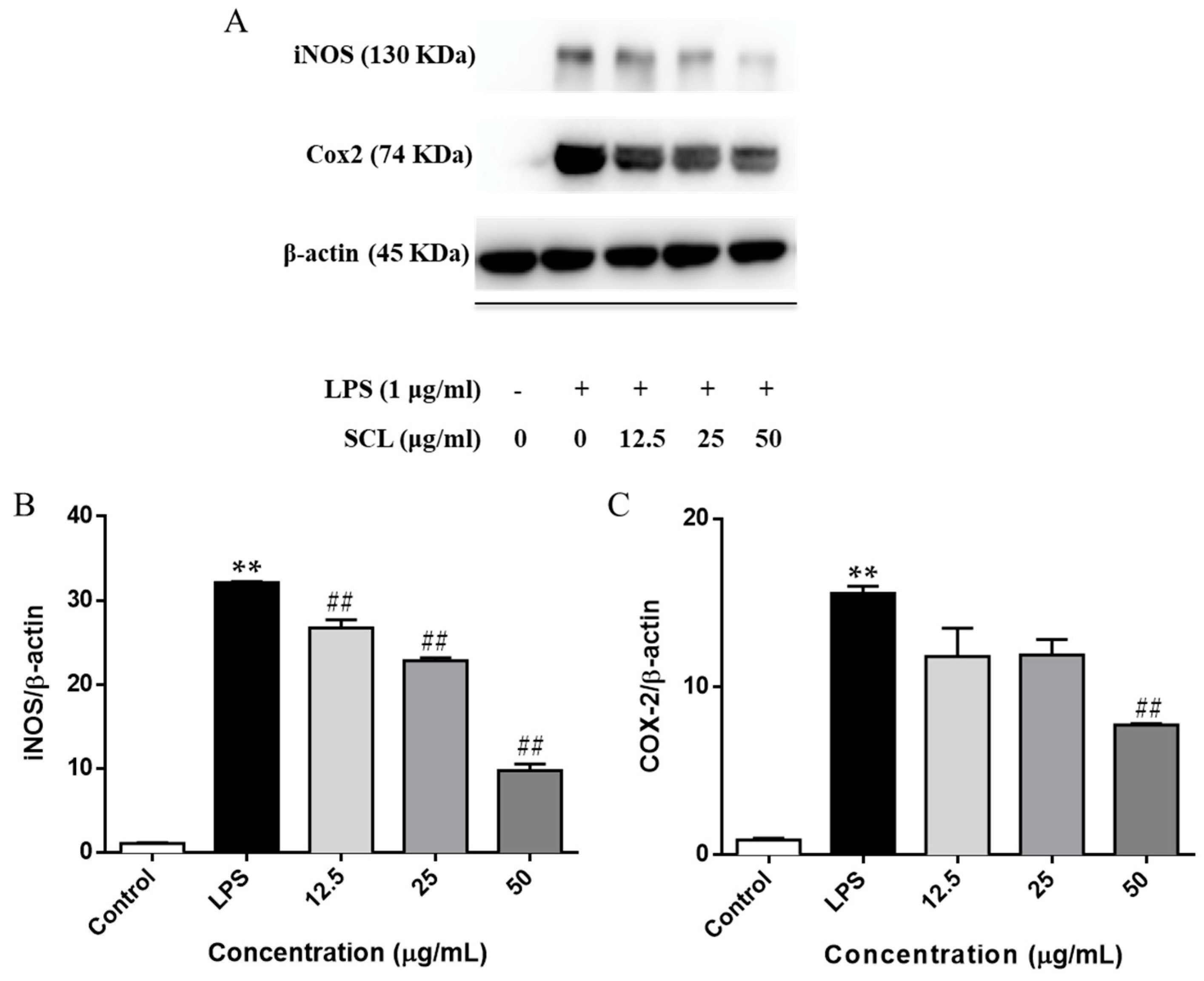
2.3. SCL Reduced the Expression of iNOS and COX-2 in LPS-Stimulated RAW264.7 Cells
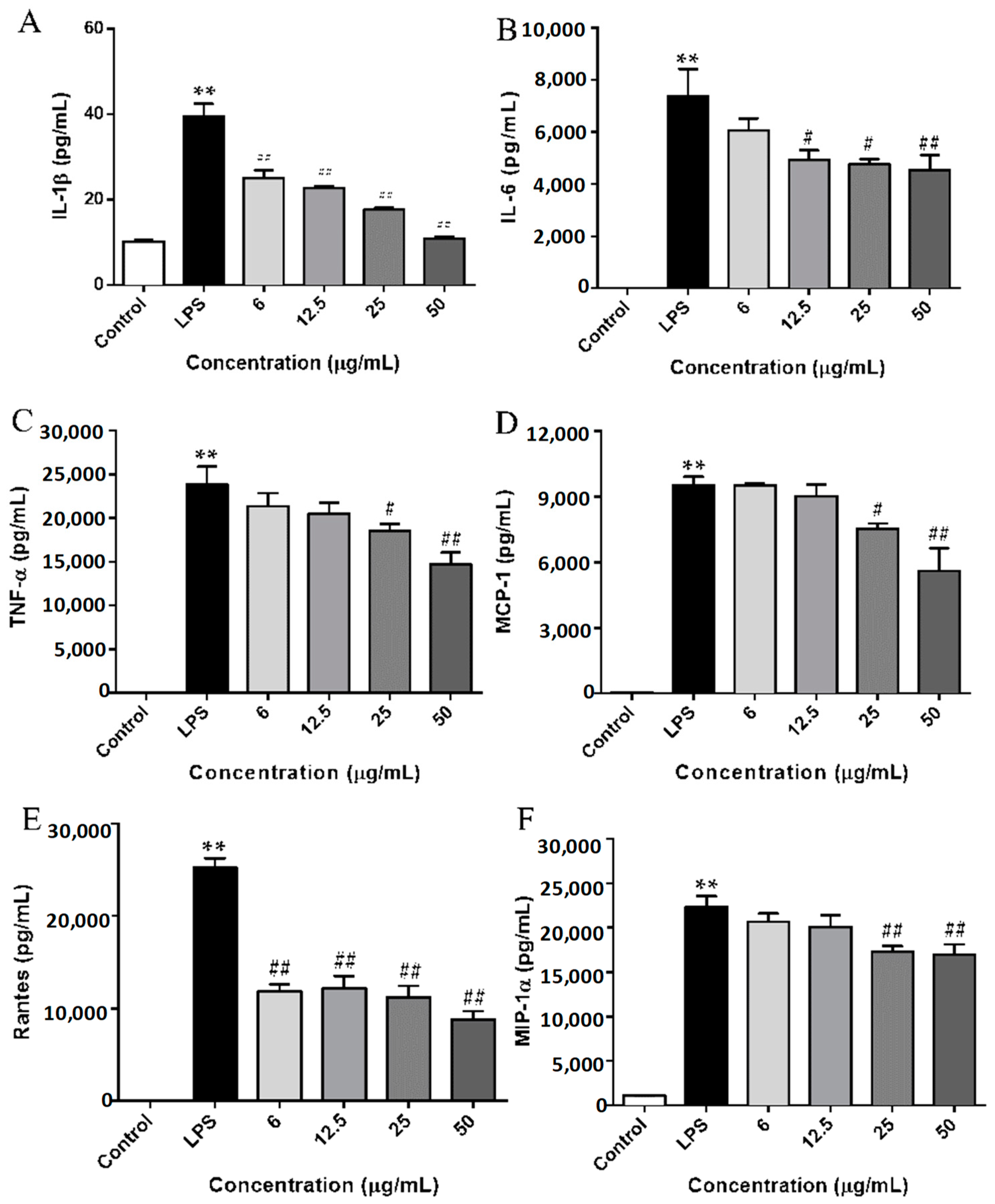
2.4. SCL Repressed the Production of Cytokines and Chemokines in LPS-Stimulated RAW264.7 Cells
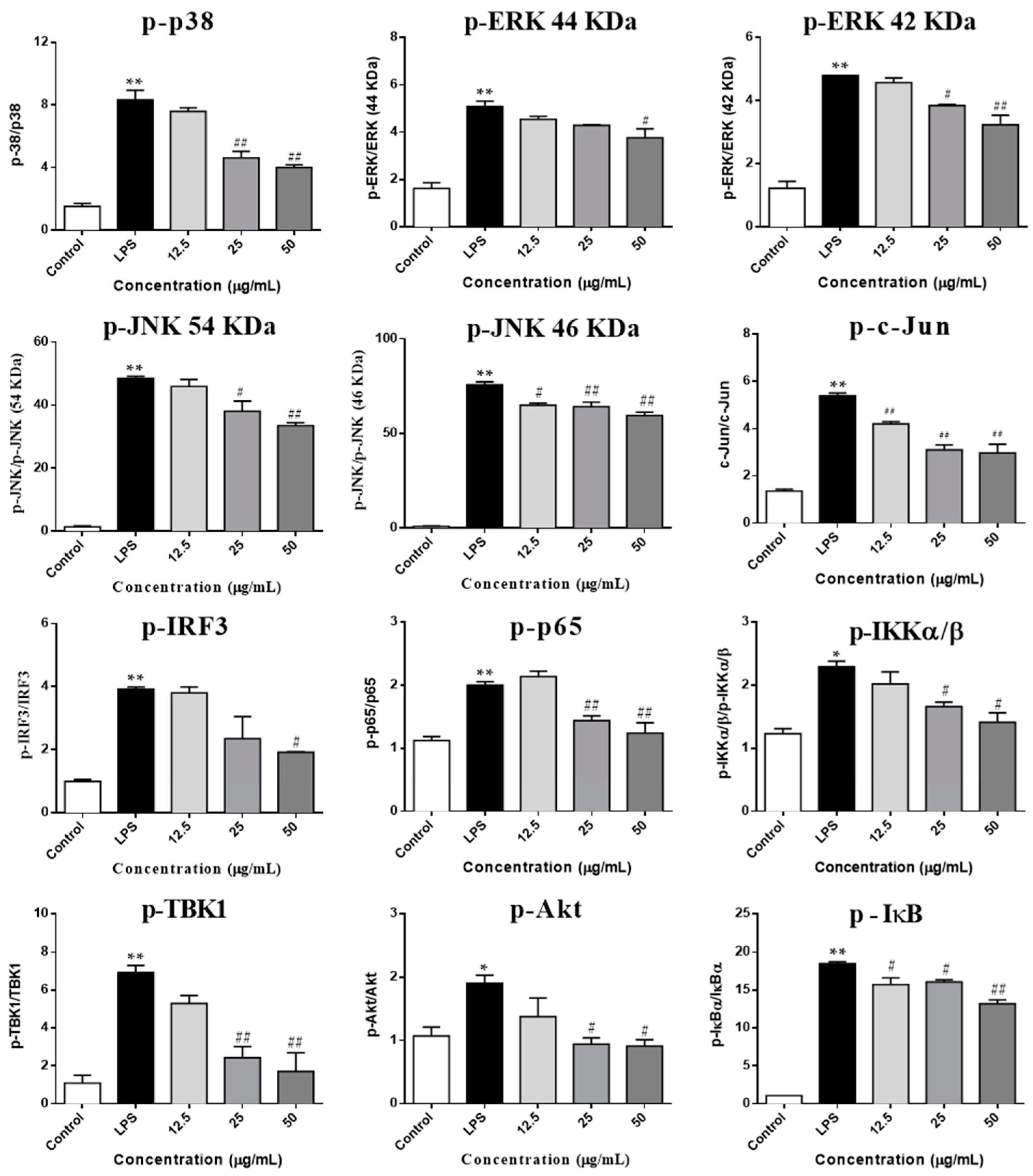
2.5. SCL Regulated on the Components of IKKα/β/NF-κB, MAPKs/AP-1 and TBK1/IRF3 Signaling Pathways
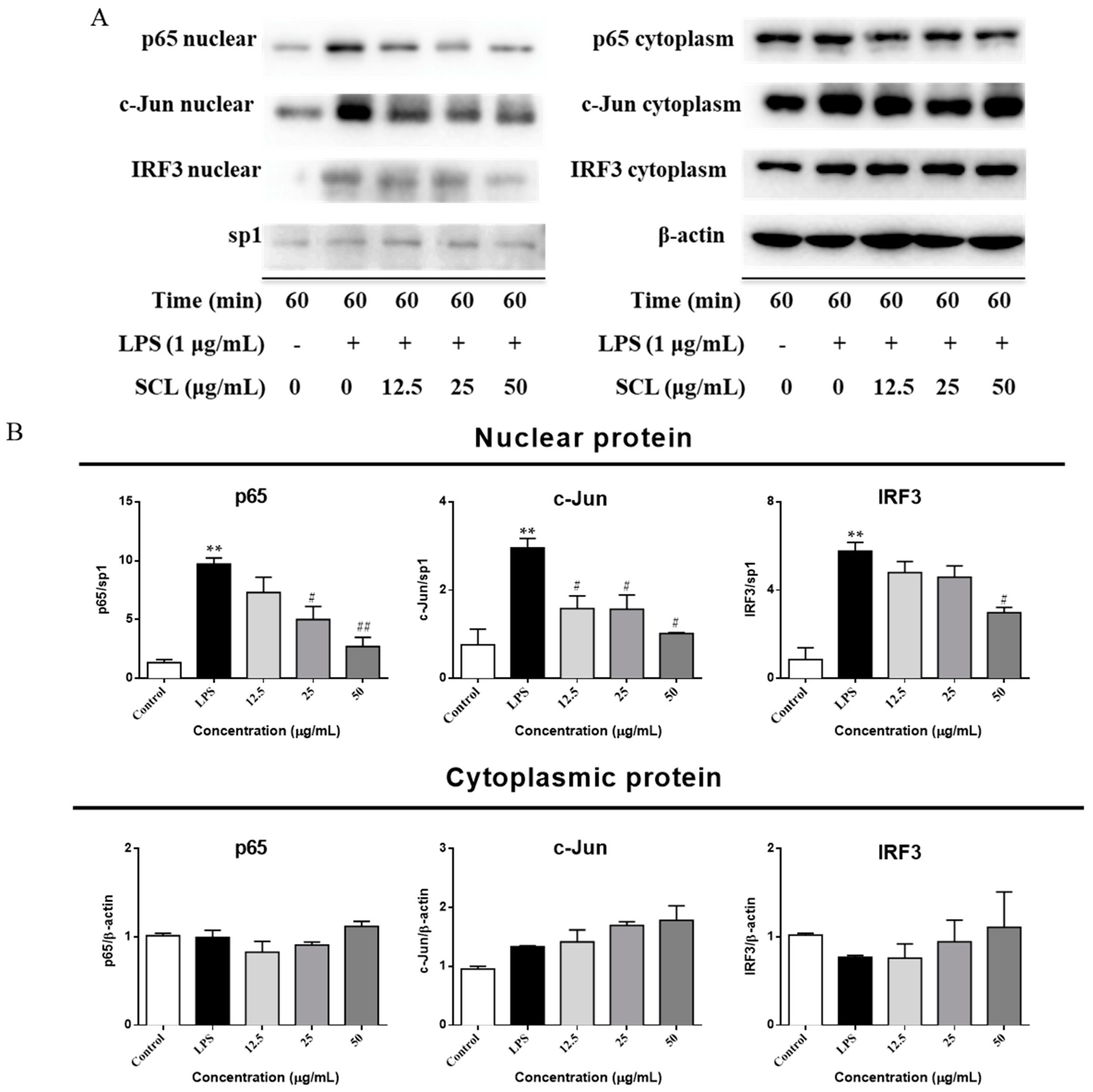
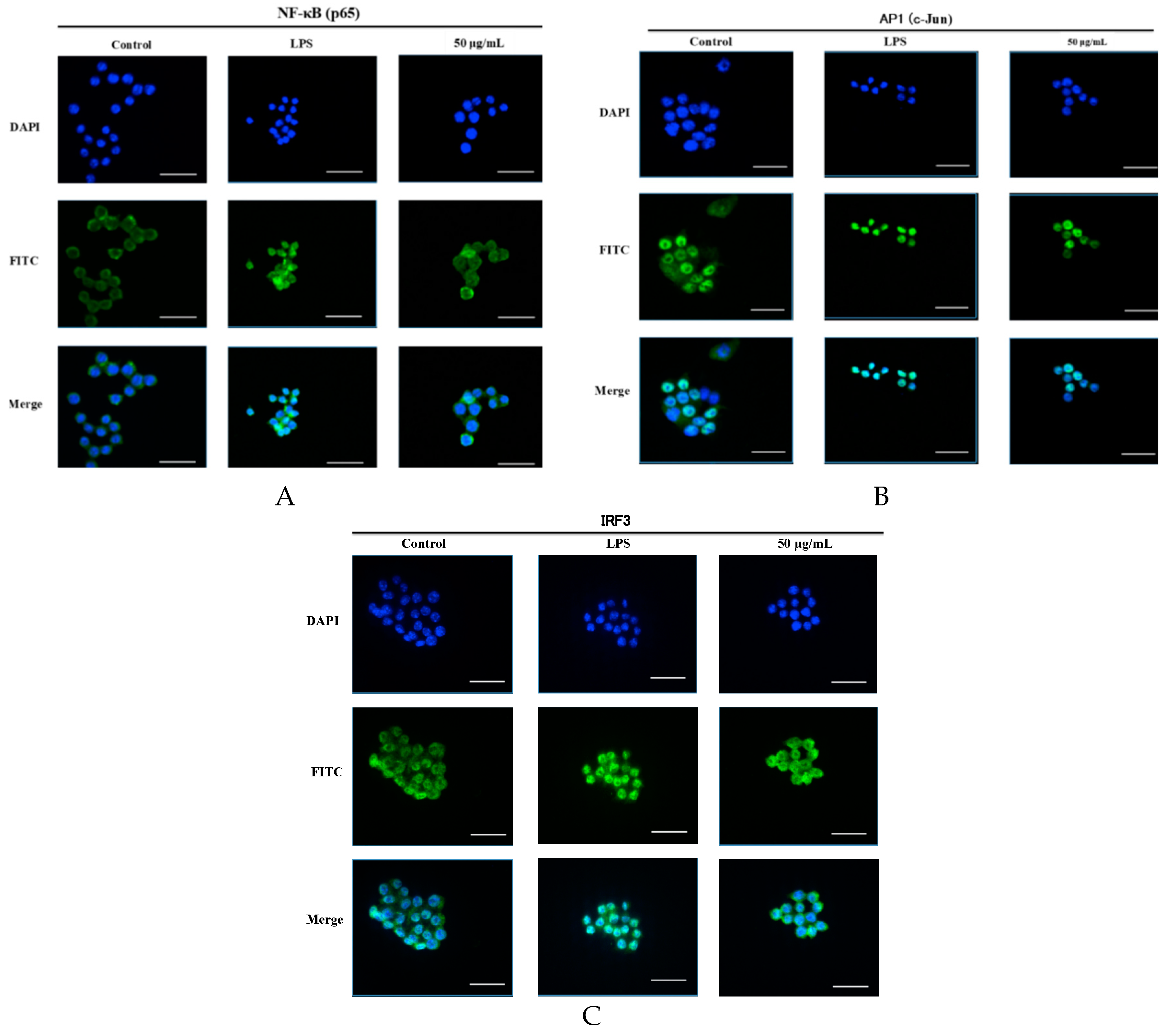
2.6. SCL Suppressed the Nuclear Translocation of AP-1, IRF3, and NF-κB in RAW264.7 Cells
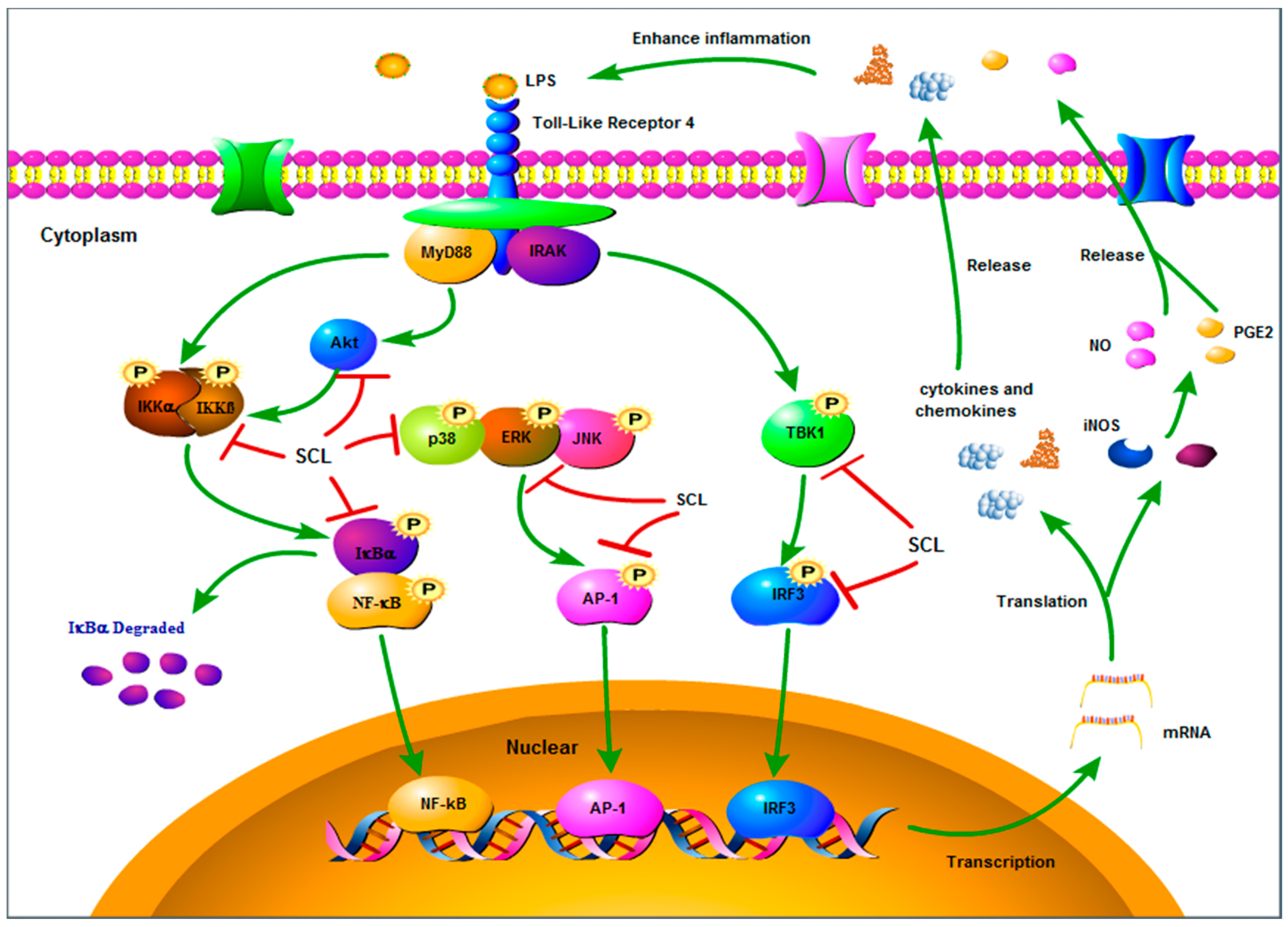
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Preparation of SCL
4.3. Characterization of SCL
4.4. Cell Culture
4.5. Cell Viability Assay
4.6. Detection of Nitric Oxide (NO)
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Western Blot Analysis
4.9. Immunofluorescence Staining
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Medzhitov, R. Inflammation 2010: new adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Choy, E.H.; Panayi, G.S. Cytokine pathways and joint inflammation in rheumatoid arthritis. N. Engl. J. Med. 2001, 344, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Montón, C.; Torres, A. Lung inflammatory response in pneumonia. Monaldi Arch. Chest Dis. 1998, 53, 56–63. [Google Scholar]
- Jenna, R.M.; Clare, M.L. Chronic inflammation and asthma. Mutat. Res. 2010, 690, 24–39. [Google Scholar]
- Liu, X.; Lin, Y.J.; Cheng, Y. Complementary and alternative therapies for inflammatory diseases. Evid. Based Complement. Altern. Med. 2016, 2016, 8324815. [Google Scholar] [CrossRef] [PubMed]
- Sostres, C.; Gargallo, C.J.; Arroyo, M.T.; Lanas, A. Adverse effects of non-steroidal anti-inflammatory drugs (NSAIDs, aspirin and coxibs) on upper gastrointestinal tract. Best Pract. Res. Clin. Gastroenterol. 2010, 24, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Ciriaco, M.; Ventrice, P.; Russo, G.; Scicchitano, M.; Mazzitello, G.; Scicchitano, F.; Russo, E. Corticosteroid-related central nervous system side effects. J. Pharmacol. Pharmacother. 2013, 4, S94–S98. [Google Scholar] [PubMed]
- Fujiwara, N.; Kobayashi, K. Macrophages in inflammation. Curr. Drug Targets Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Iwanaszko, M.; Kimmel, M. NF-κB and IRF pathways: Cross-regulation on target genes promoter level. BMC Genom. 2015, 16, 307. [Google Scholar] [CrossRef] [PubMed]
- Czaja, A.J. Hepatic inflammation and progressive liver fibrosis in chronic liver disease. World J. Gastroenterol. 2014, 20, 2515–2532. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, Y.; Wang, Y.; Ning, Q.; Zhang, Y.; Gong, C.; Zhao, W.; Jing, G.; Wang, Q. Dexmedetomidine attenuates inflammatory reaction in the lung tissues of septic mice by activating cholinergic anti-inflammatory pathway. Int. Immunopharmacol. 2016, 35, 210–216. [Google Scholar] [CrossRef] [PubMed]
- WHO Traditional Medicine Strategy 2002–2005. Available online: http://apps.who.int/medicinedocs/en/d/Js2297e/ (accessed on 25 November 2018).
- Li, X.M. Traditional Chinese herbal remedies for asthma and food allergy. J. Allergy Clin. Immunol. 2007, 120, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Jiang, M.; Lü, A.P. Evidence-based Chinese medicine for rheumatoid arthritis. J. Tradit. Chin. Med. 2011, 31, 152–157. [Google Scholar] [CrossRef]
- Hou, P.W.; Fu, P.K.; Hsu, H.C.; Hsieh, C.L. Traditional Chinese medicine in patients with osteoarthritis of the knee. J. Tradit. Complement. Med. 2015, 5, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Panossian, A.; Wikman, G. Pharmacology of Schisandra Chinensis Bail.: An overview of Russian research and uses in medicine. J. Ethnopharmacol. 2008, 118, 183–212. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; He, Q.; Zhang, Y.; Qu, H.; Cheng, Y. Characterisation and identification of isomeric dibenzocyclooctadiene lignans from Schisandra Chinensis by high-performance liquid chromatography combined with electrospray ionisation tandem mass spectrometry. Phytochem. Anal. 2009, 20, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, W.J.; Song, Y.Z.; Liang, Z.Q. The protective mechanism of schisandrin A in galactosamine-induced acute liver injury through activation of autophagy. Pharm. Biol. 2014, 52, 1302. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, Y.; Qian, H.Y.; Yoshigai, E.; Okumura, T.; Ikeya, Y.; Nishizawa, M. Gomisin N in the herbal drug gomishi (Schisandra Chinensis) suppresses inducible nitric oxide synthase gene via C/EBPβ and NF-κB in rat hepatocytes. Nitric Oxide 2013, 28, 47–56. [Google Scholar] [CrossRef]
- Chiu, P.Y.; Leung, H.Y.; Poon, M.K.; Lee, S.S.; Ko, K.M. Schisandrin B induced antioxidant response is partly mediated by cytochrome P-4502E1 catalyzed reaction in mouse liver. Mol. Cell. Biochem. 2006, 293, 87–92. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, Z.; Sun, J.; Pan, Q.; Sun, F.; Yan, Z.; Hu, X. Schisandrin B prevents doxorubicin induced chronic cardiotoxicity and enhances its anticancer activity in vivo. PLoS ONE 2011, 6, e28335. [Google Scholar] [CrossRef]
- Leong, P.K.; Wong, H.S.; Chen, J.; Chan, W.M.; Leung, H.Y.; Ko, K.M. Differential action between Schisandrin A and Schisandrin B in eliciting an anti-inflammatory action: The depletion of reduced glutathione and the induction of an antioxidant response. PLoS ONE 2016, 11, e0155879. [Google Scholar] [CrossRef] [PubMed]
- Taciak, B.; Białasek, M.; Braniewska, A.; Sas, Z.; Sawicka, P.; Kiraga, Ł.; Rygiel, T.; Król, M. Evaluation of phenotypic and functional stability of RAW 264.7 cell line through serial passages. PLoS ONE 2018, 13, e0198943. [Google Scholar] [CrossRef] [PubMed]
- Bauerfeld, C.P.; Rastogi, R.; Pirockinaite, G.; Lee, I.; Hüttemann, M.; Monks, B.; Birnbaum, M.J.; Franchi, L.; Nuñez, G.; Samavati, L. TLR4-mediated AKT activation is MyD88/TRIF dependent and critical for induction of oxidative phosphorylation and mitochondrial transcription factor A in murine macrophages. J. Immunol. 2012, 188, 2847–2857. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.H.; Yang, C.C.; Li, P.C.; Chen, W.C.; Chien, C.T. Therapeutic potential of traditional Chinese medicine on inflammatory diseases. J. Tradit. Complement. Med. 2013, 3, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Ip, S.P.; Poon, M.K.; Wu, S.S.; Che, C.T.; Ng, K.H.; Kong, Y.C.; Ko, K.M. Effect of schisandrin B on hepatic glutathione antioxidant system in mice: Protection against carbon tetrachloride toxicity. Planta Med. 1995, 61, 398–401. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.Y.; Kim, Y.H.; Bae, D.S.; Um, B.H.; Pan, C.H.; Kim, C.Y.; Lee, H.J.; Lee, J.K. Anti-inflammatory effects of gomisin N, gomisin J, and schisandrin C isolated from the fruit of Schisandra chinensis. Biosci. Biotechnol. Biochem. 2010, 74, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Laroux, F.S.; Pavlick, K.P.; Hines, I.N.; Kawachi, S.; Harada, H.; Bharwani, S.; Hoffman, J.M.; Grisham, M.B. Role of nitric oxide in inflammation. Acta Physiol. Scand. 2001, 173, 113–118. [Google Scholar] [CrossRef]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef]
- Medeiros, A.; Peres-Buzalaf, C.; Fortino-Verdan, F.; Serezani, C.H. Prostaglandin E2 and the suppression of phagocyte innate immune responses in different organs. Mediat. Inflamm. 2012, 2012, 327568. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Salvemini, D.; Marino, M.H. Inducible nitric oxide synthase and inflammation. Expert Opin. Investig. Drugs 1998, 7, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Guo, H.; Cheng, B.C.; Su, T.; Fu, X.Q.; Li, T.; Zhu, P.L.; Tse, K.W.; Pan, S.Y.; Yu, Z.L. Dingchuan tang essential oil inhibits the production of inflammatory mediators via suppressing the IRAK/NF-κB, IRAK/AP-1, and TBK1/IRF3 pathways in lipopolysaccharide-stimulated RAW264.7 cells. Drug Des. Dev. Ther. 2018, 12, 2731–2748. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-kappaB and IkappaB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shepherd, E.G.; Nelin, L.D. MAPK phosphatases—Regulating the immune response. Nat. Rev. Immunol. 2007, 7, 202–212. [Google Scholar] [CrossRef]
- Karin, M. The regulation of AP-1 activity by mitogen-activated protein kinases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 127–134. [Google Scholar] [CrossRef]
- Ozes, O.N.; Mayo, L.D.; Gustin, J.A.; Pfeffer, S.R.; Pfeffer, L.M.; Donner, D.B. NF-kappaB activation by tumour necrosis factor requires the Akt serine- threonine kinase. Nature 1999, 401, 82–85. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Wang, N.; Liang, H.; Zen, K. Molecular mechanisms that influence the macrophage m1-m2 polarization balance. Front. Immunol. 2014, 5, 614. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Rowe, D.C.; Barnes, B.J.; Caffrey, D.R.; Visintin, A.; Latz, E.; Monks, B.; Pitha, P.M.; Golenbock, D.T. LPS-TLR4 signaling to IRF-3/7 and NF-kappaB involves the toll adapters TRAM and TRIF. J. Exp. Med. 2003, 198, 1043–1055. [Google Scholar] [CrossRef]
- Bryan, N.S.; Grisham, M.B. Methods to detect nitric oxide and its Metabolites in biological samples. Free Radic. Biol. Med. 2007, 43, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.C.; Yu, H.; Su, T.; Fu, X.Q.; Guo, H.; Li, T.; Cao, H.H.; Tse, A.K.; Kwan, H.Y.; Yu, Z.L. A herbal formula comprising Rosae Multiflorae Fructus and Lonicerae Japonicae Flos inhibits the production of inflammatory mediators and the IRAK-1/TAK1 and TBK1/IRF3 pathways in RAW264.7 and THP-1 cells. J. Ethnopharmacol. 2015, 174, 195–199. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. | Retention Time (min) | Compounds | Content in Sample (mg/g) |
|---|---|---|---|
| 1 | 23.554 | Schisandrin | 303.03 |
| 2 | 33.907 | Schisandrol B | 89.86 |
| 3 | 64.140 | Schisantherin A | 44.93 |
| 4 | 81.137 | Schisandrin A | 57.47 |
| 5 | 90.831 | Schisandrin B | 103.45 |
| 6 | 93.303 | Schisandrin C | 7.84 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, G.; Cheng, B.C.-Y.; Zhao, H.; Fu, X.-Q.; Xie, R.; Zhang, S.-F.; Pan, S.-Y.; Zhang, Y. Schisandra Chinensis Lignans Suppresses the Production of Inflammatory Mediators Regulated by NF-κB, AP-1, and IRF3 in Lipopolysaccharide-Stimulated RAW264.7 Cells. Molecules 2018, 23, 3319. https://doi.org/10.3390/molecules23123319
Luo G, Cheng BC-Y, Zhao H, Fu X-Q, Xie R, Zhang S-F, Pan S-Y, Zhang Y. Schisandra Chinensis Lignans Suppresses the Production of Inflammatory Mediators Regulated by NF-κB, AP-1, and IRF3 in Lipopolysaccharide-Stimulated RAW264.7 Cells. Molecules. 2018; 23(12):3319. https://doi.org/10.3390/molecules23123319
Chicago/Turabian StyleLuo, Gan, Brian Chi-Yan Cheng, Hui Zhao, Xiu-Qiong Fu, Ran Xie, Shuo-Feng Zhang, Si-Yuan Pan, and Yi Zhang. 2018. "Schisandra Chinensis Lignans Suppresses the Production of Inflammatory Mediators Regulated by NF-κB, AP-1, and IRF3 in Lipopolysaccharide-Stimulated RAW264.7 Cells" Molecules 23, no. 12: 3319. https://doi.org/10.3390/molecules23123319