Monohexosylceramides from Rhizopus Species Isolated from Brazilian Caatinga: Chemical Characterization and Evaluation of Their Anti-Biofilm and Antibacterial Activities


 ,
,
Abstract
:
1. Introduction
2. Results
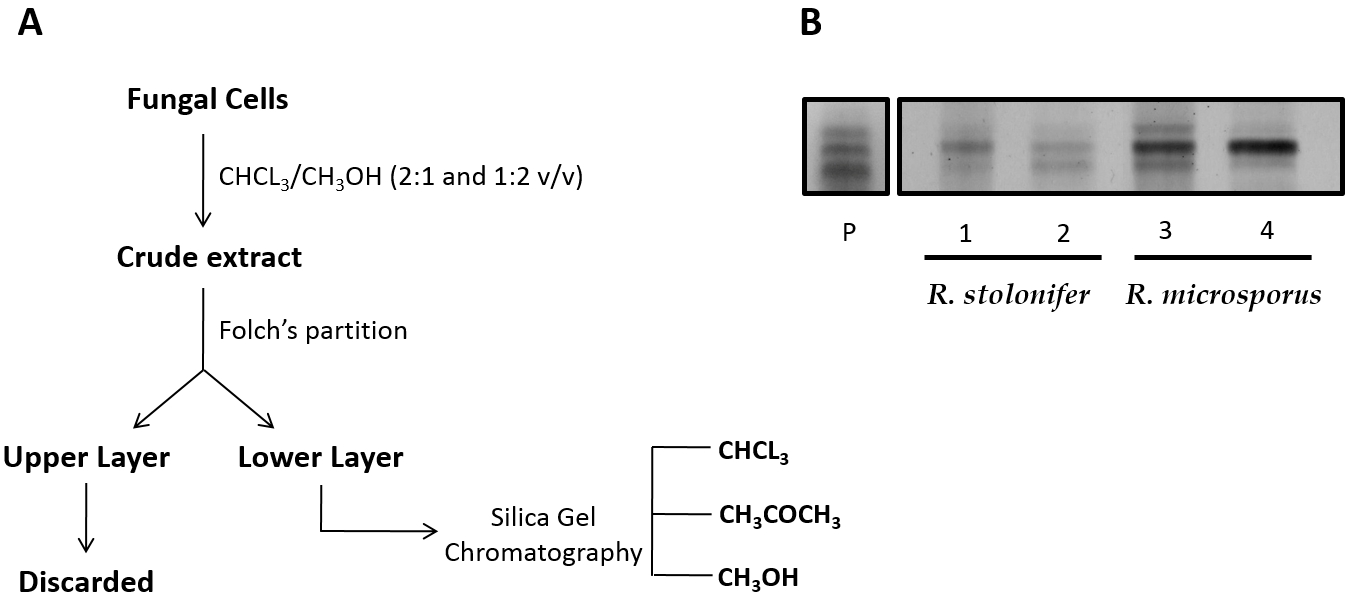
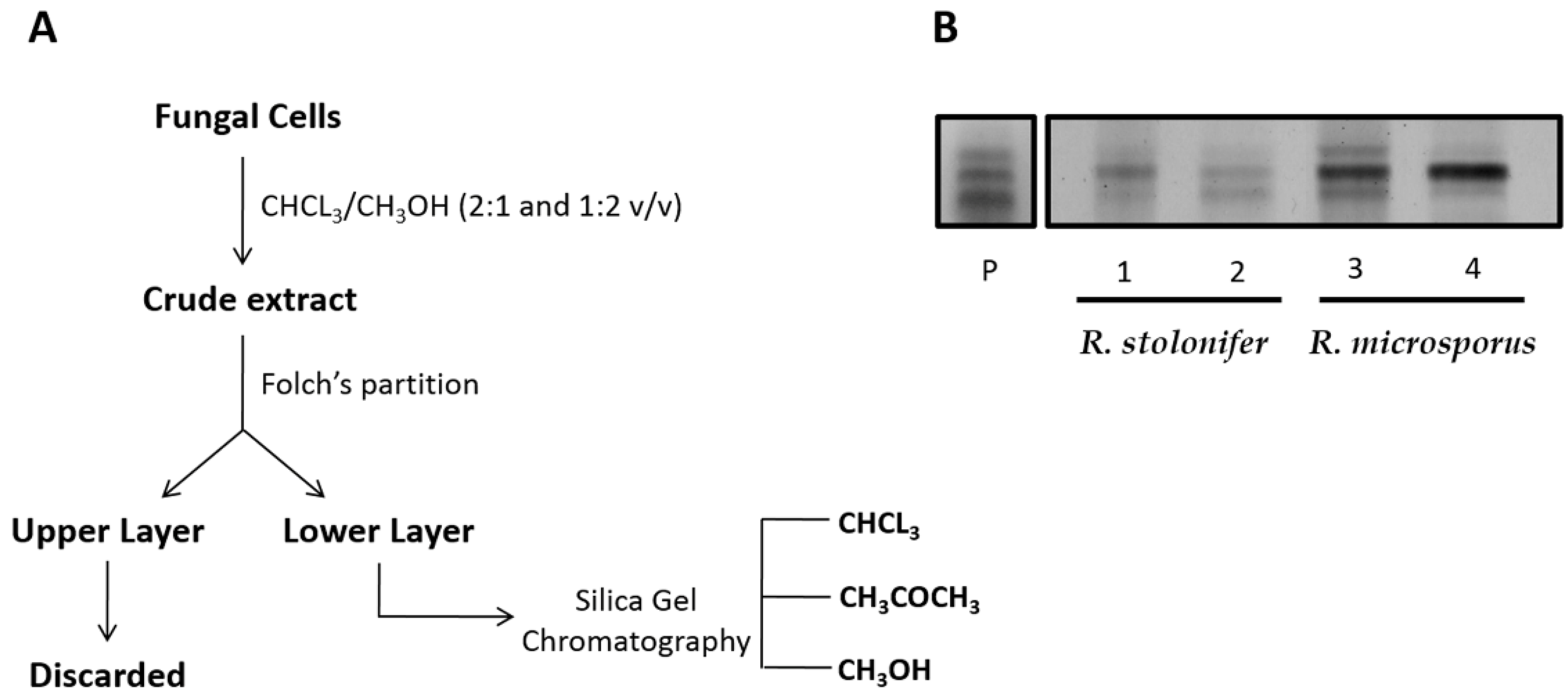
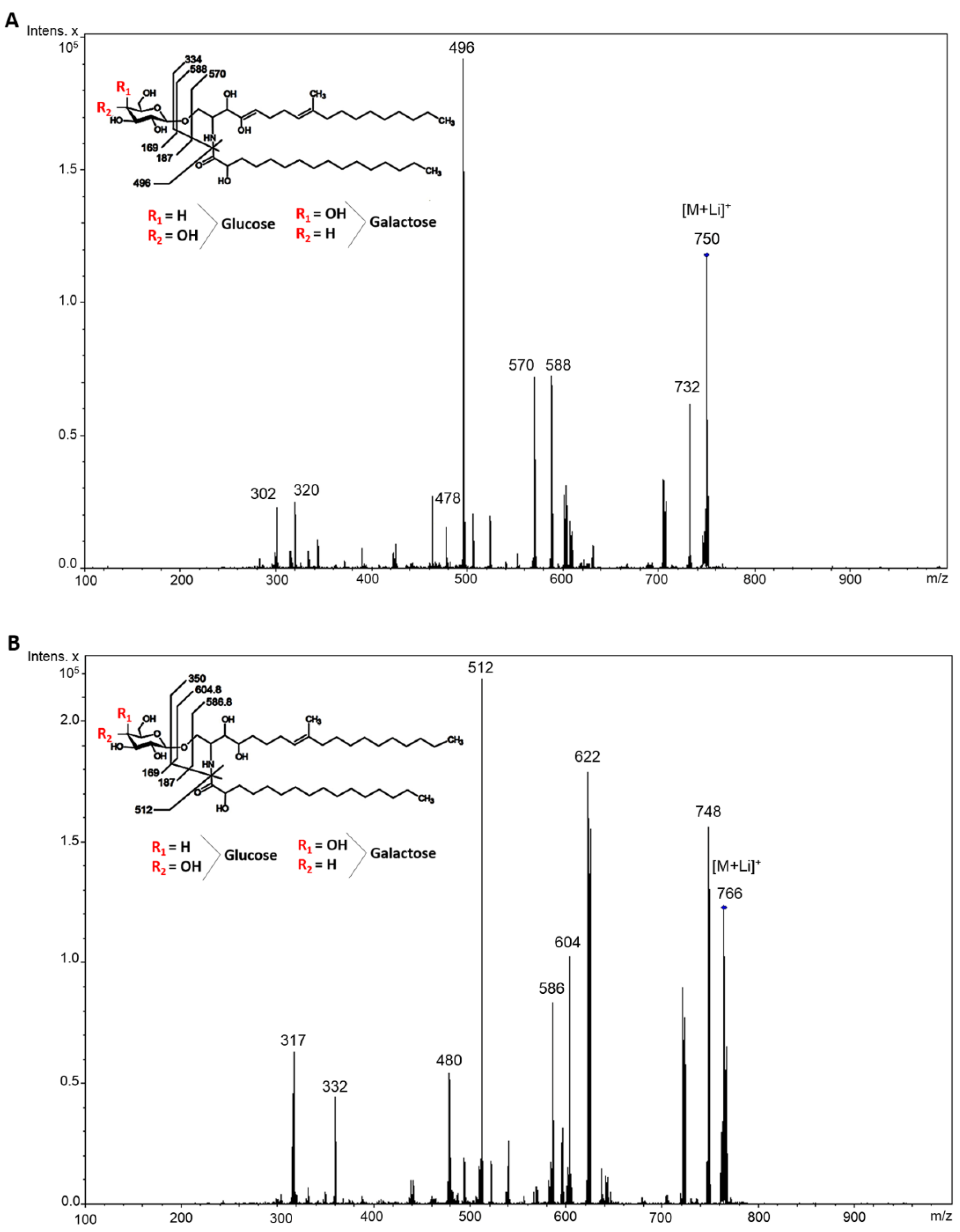
2.1. CMH Purification and Chemical Analysis
2.2. Antibacterial Activity Evaluation
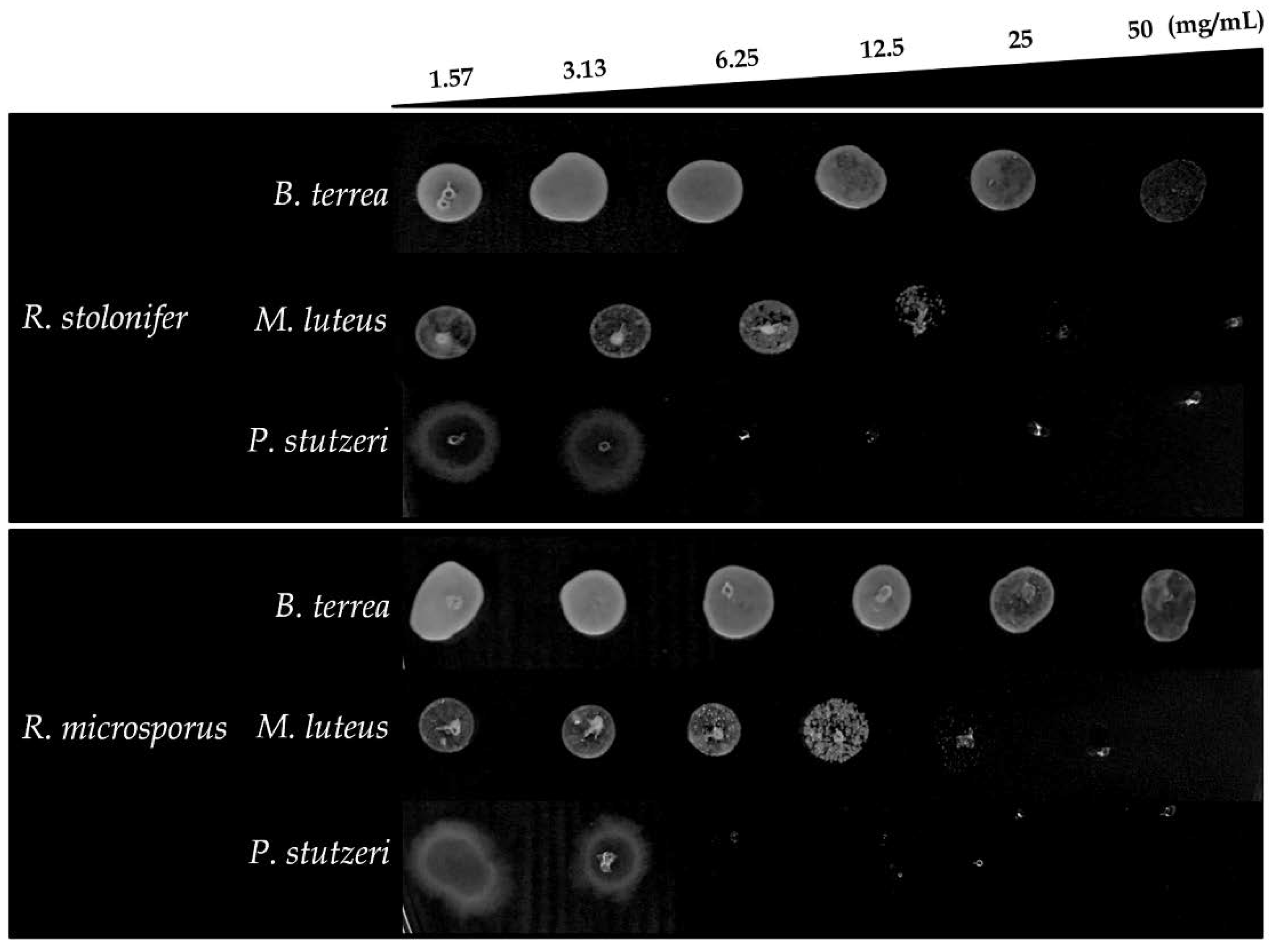
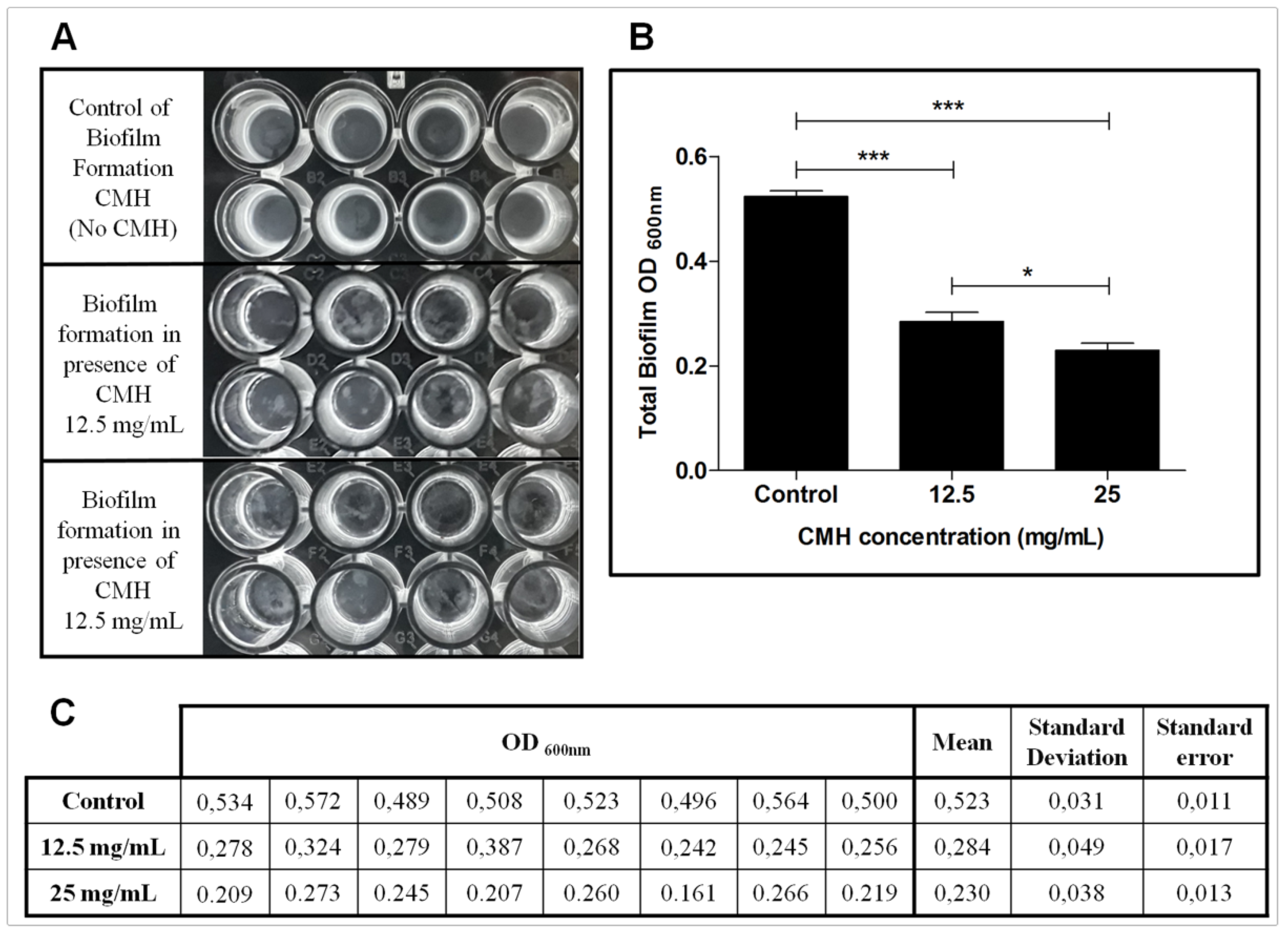
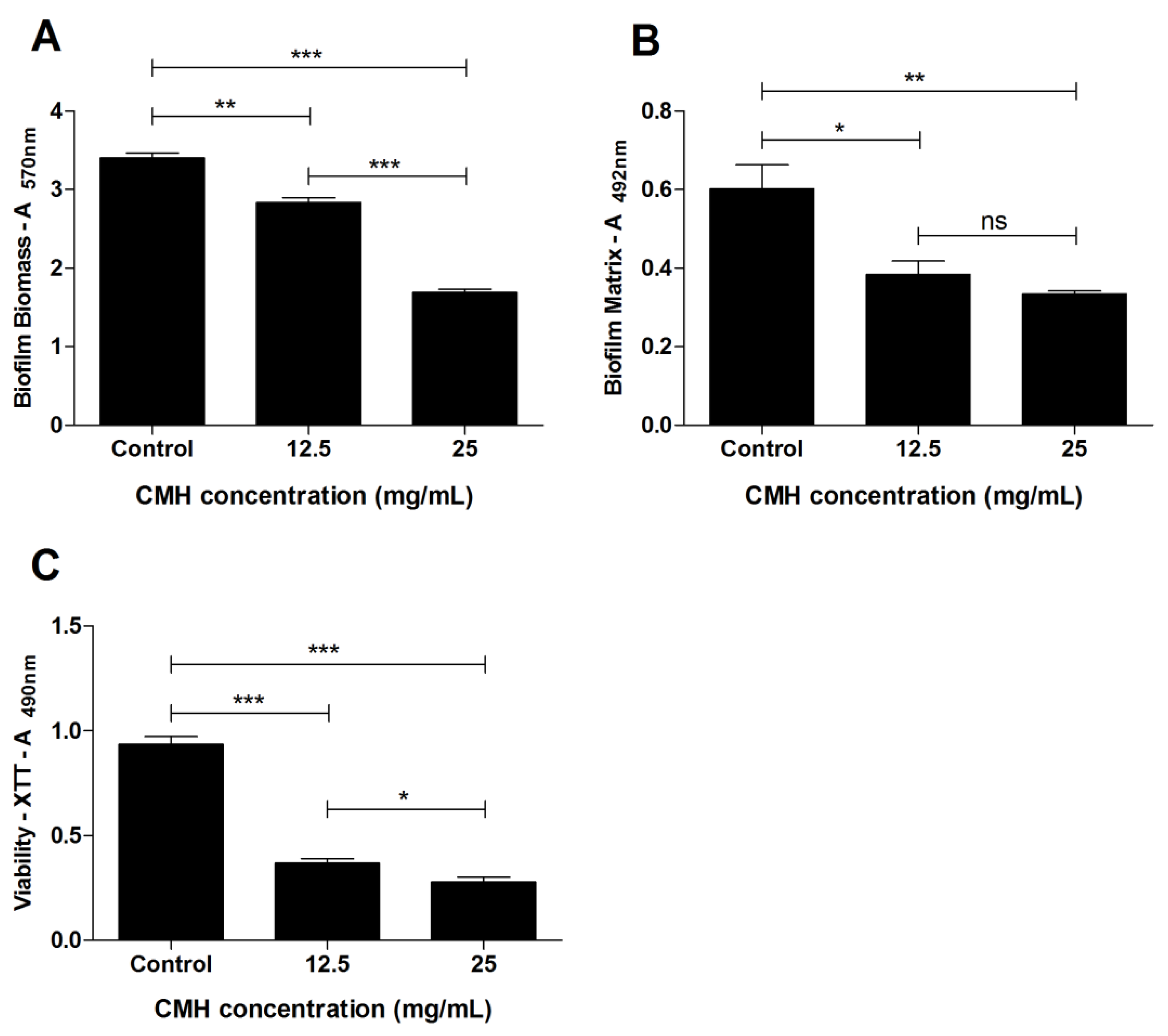
2.3. Evaluation of Anti-Biofilm Activity
3. Discussion
4. Material and Methods
4.1. Microorganisms and Culture Conditions
4.2. Extraction and Purification of CMH from R. stolonifer and R. microsporus
4.3. Sugar Analysis
4.4. ESI-MS Analysis
4.5. Antimicrobial Assay
4.5.1. Bacterial Strains
4.5.2. Antimicrobial Activity Assay
4.5.3. Determination of MIC and MBC
4.5.4. Effect of CMH on Inhibition of MRSA ATCC9393 Biofilm
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Azevedo Santiago, A.L.C.M.; dos Santos, P.J.P.; Maia, L.C. Mucorales from the semiarid of Pernambuco, Brazil. Braz. J. Microbiol. 2013, 44, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, D.X.; De Azevedo Santiago, A.L.C.M.; De Souza-Motta, C.M. Diversity of mucorales in natural and degraded semi-arid soils. Braz. J. Bot. 2016, 39, 1127–1133. [Google Scholar] [CrossRef]
- Lackner, M.; Caramalho, R.; Lass-Flörl, C. Laboratory diagnosis of mucormycosis: Current status and future perspectives. Future Microbiol. 2014, 9, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Ribes, J.A.; Vanover-Sams, C.L.; Baker, D.J. Zygomycetes in human disease. Clin. Microbiol. Rev. 2000, 13, 236–301. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.R.; Rodrigues, M.L.; Travassos, L.R.; Haido, R.M.T.; Wait, R.; Barreto-Bergter, E. Characterization of glucosylceramides in Pseudallescheria boydii and their involvement in fungal differentiation. Glycobiology 2002, 12, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Bergter, E.; Pinto, M.R.; Rodrigues, M.L. Structure and biological functions of fungal cerebrosides. An. Acad. Bras. Ciênc. 2004, 76, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Barreto-Bergter, E.; Sassaki, G.L.; de Souza, L.M. Structural analysis of fungal cerebrosides. Front. Microbiol. 2011, 2, 239. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.F.C.; Rodrigues, M.L.; Farias, S.E.; Almeida, I.C.; Pinto, M.R.; Barreto-Bergter, E. Glucosylceramides in colletotrichum gloeosporioides are involved in the differentiation of conidia into mycelial cells. FEBS Lett. 2004, 561, 137–143. [Google Scholar] [CrossRef]
- Nimrichter, L.; Cerqueira, M.D.; Leitão, E.A.; Miranda, K.; Nakayasu, E.S.; Almeida, S.R.; Almeida, I.C.; Alviano, C.S.; Barreto-Bergter, E.; Rodrigues, M.L. Structure, cellular distribution, antigenicity, and biological functions of Fonsecaea pedrosoi ceramide monohexosides. Infect. Immun. 2005, 73, 7860–7868. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Travassos, L.R.; Miranda, K.R.; Franzen, A.J.; Rozental, S.; de Souza, W.; Alviano, C.S.; Barreto-Bergter, E. Human antibodies against a purified glucosylceramide from Cryptococcus neoformans inhibit cell budding and fungal growth. Infect. Immun. 2000, 68, 7049–7060. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Cahoon, E.B.; Thokala, M.; Kaur, J.; Li, J.; Shah, D.M. Sphingolipid c-9 methyltransferases are important for growth and virulence but not for sensitivity to antifungal plant defensins in Fusarium graminearum. Eukaryot. Cell 2009, 8, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Rollin-Pinheiro, R.; Singh, A.; Barreto-Bergter, E.; Del Poeta, M. Sphingolipids as targets for treatment of fungal infections. Future Med. Chem. 2016, 8, 1469–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Takakuwa, N.; Ohnishi, M.; Oda, Y. Presence of glucosylceramide in yeast and its relation to alkali tolerance of yeast. Appl. Microbiol. Biotechnol. 2006, 71, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Thevissen, K.; Warnecke, D.C.; François, I.E.J.A.; Leipelt, M.; Heinz, E.; Ott, C.; Zähringer, U.; Thomma, B.P.H.J.; Ferket, K.K.A.; Cammue, B.P.A. Defensins from insects and plants interact with fungal glucosylceramides. J. Biol. Chem. 2004, 279, 3900–3905. [Google Scholar] [CrossRef] [PubMed]
- Cateni, F.; Zilic, J.; Falsone, G.; Scialino, G.; Banfi, E. New cerebrosides from Euphorbia peplis L.: Antimicrobial activity evaluation. Bioorgan. Med. Chem. Lett. 2003, 13, 4345–4350. [Google Scholar] [CrossRef]
- Derengowski, L.S.; De-Souza-Silva, C.; Braz, S.V.; Mello-De-Sousa, T.M.; Báo, S.N.; Kyaw, C.M.; Silva-Pereira, I. Antimicrobial effect of farnesol, a Candida albicans quorum sensing molecule, on Paracoccidioides brasiliensis growth and morphogenesis. Ann. Clin. Microbiol. Antimicrob. 2009, 8, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, R.G.; Wang, F.W.; Yang, Y.M.; Liu, Y.X.; Tan, R.X. Antibacterial and xanthine oxidase inhibitory cerebrosides from Fusarium sp. IFB-121, and endophytic fungus in quercus variabilis. Lipids 2004, 39, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-P.; Chen, Y.; Xia, B.; Wang, M.; Dong, Y.-F.; Feng, X. Two novel ceramides with a phytosphingolipid and a tertiary amide structure from Zephyranthes candida. Lipids 2009, 44, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, P.C.S.; Monteiro, A.S.; Marques, S.G.; Monteiro, S.G.; Monteiro-Neto, V.; Coqueiro, M.M.M.; Marques, A.C.G.; de Jesus Gomes Turri, R.; Santos, S.G.; Bomfim, M.R.Q. Phenotypic and molecular detection of the bla(KPC) gene in clinical isolates from inpatients at hospitals in São Luis, MA, Brazil. BMC Infect. Dis. 2016, 16, 737. [Google Scholar] [CrossRef] [PubMed]
- Rossi Gonçalves, I.; Ferreira, M.L.; Araujo, B.F.; Campos, P.A.; Royer, S.; Batistão, D.W.F.; Souza, L.P.; Brito, C.S.; Urzedo, J.E.; Gontijo-Filho, P.P.; et al. Outbreaks of colistin-resistant and colistin-susceptible KPC-producing klebsiella pneumoniae in a brazilian intensive care unit. J. Hosp. Infect. 2016, 94, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Tognim, M.C.B.; Cardoso, C.L. Nosocomial outbreaks in brazil: Can they be controlled? J. Hosp. Infect. 2016, 94, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Lima-e-Silva, A.A.; Silva-Filho, R.G.; Fernandes, H.M.Z.; Saramago, C.S.M.; Viana, A.S.; Souza, M.J.; Nogueira, E.M. Sub-inhibitory concentrations of rifampicin strongly stimulated biofilm production in S. aureus. Open Microbiol. J. 2017, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rojas, A.; Makarova, O.; Rolff, J. Antimicrobials, stress and mutagenesis. PLoS Pathog. 2014, 10, e1004445. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, A.; Xenophontos, E.; Karatsi, A.; Petrides, C.; Kleridou, M.; Zintilis, C. Insidious manifestation of pyogenic liver abscess caused by streptococcus intermedius and micrococcus luteus: A case report. Oxf. Med. Case Rep. 2016, 2016, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Bennasar, A.; Bosch, R.; García-Valdés, E.; Palleroni, N.J. Biology of Pseudomonas stutzeri. Microbiol. Mol. Biol. Rev. 2006, 70, 510–547. [Google Scholar] [CrossRef] [PubMed]
- Miltiadous, G.; Elisaf, M. Native valve endocarditis due to micrococcus luteus: A case report and review of the literature. J. Med. Case Rep. 2011, 5, 251. [Google Scholar] [CrossRef] [PubMed]
- Miron, D.; Keness, Y.; Bor, N.; Spiegel, R.; Horowitz, Y. Pseudomonas stutzeri knee arthritis in a child: Case report and review. J. Pediatr. Orthop. B 2007, 16, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, L.L.; Toledo, M.S.; Ferreira, F.A.S.; Straus, A.H.; Takahashi, H.K. Structural diversity and biological significance of glycosphingolipids in pathogenic and opportunistic fungi. Front. Cell. Infect. Microbiol. 2014, 4, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, L.V.G.; Nakayasu, E.S.; Gazos-Lopes, F.; Vallejo, M.C.; Matsuo, A.L.; Almeida, I.C.; Puccia, R. Characterization of cell wall lipids from the pathogenic phase of Paracoccidioides brasiliensis cultivated in the presence or absence of human plasma. PLoS ONE 2013, 8, e63372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, H.K.; Toledo, M.S.; Suzuki, E.; Tagliari, L.; Straus, A.H. Current relevance of fungal and trypanosomatid glycolipids and sphingolipids: Studies defining structures conspicuously absent in mammals. An. Acad. Bras. Ciênc. 2009, 81, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Glycosynapses: Microdomains controlling carbohydrate-dependent cell adhesion and signaling. An. Acad. Bras. Ciênc. 2004, 76, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S.-I. Structure and function of glycosphingolipids and sphingolipids: Recollections and future trends. Biochim. Biophys. Acta 2008, 1780, 325–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoki, K.; Uchiyama, R.; Yamauchi, S.; Katayama, T.; Itonori, S.; Sugita, M.; Hada, N.; Yamada-Hada, J.; Takeda, T.; Kumagai, H.; et al. Newly discovered neutral glycosphingolipids in aureobasidin a-resistant zygomycetes: Identification of a novel family of gala-series glycolipids with core galα1-6galβ1-6galβ sequences. J. Biol. Chem. 2004, 279, 32028–32034. [Google Scholar] [CrossRef] [PubMed]
- Weinstein, R.A. Controlling antimicrobial resistance in hospitals: Infection control and use of antibiotics. Emerg. Infect. Dis. 2001, 7, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Sánchez Ade, J.; Hernández-Sánchez, H.; Jaramillo-Flores, M.E. Biological activity of glycolipids produced by microorganisms: New trends and possible therapeutic alternatives. Microbiol. Res. 2013, 168, 22–32. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E.W. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Sambanthamoorthy, K.; Feng, X.; Patel, R.; Patel, S.; Paranavitana, C. Antimicrobial and antibiofilm potential of biosurfactants isolated from lactobacilli against multi-drug-resistant pathogens. BMC Microbiol. 2014, 14, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folch, J.; Lees, M.; Stanley, G.H.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Leitão, E.A.; Bittencourt, V.C.B.; Haido, R.M.T.; Valente, A.P.; Peter-Katalinic, J.; Letzel, M.; de Souza, L.M.; Barreto-Bergter, E. B-galactofuranose-containing O-linked oligosaccharides present in the cell wall peptidogalactomannan of aspergillus fumigatus contain immunodominant epitopes. Glycobiology 2003, 13, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Anal. Chem. 1965, 37, 1602–1604. [Google Scholar] [CrossRef]
- Calixto, R.O.R.; Rollin-Pinheiro, R.; da Silva, M.I.D.; Liporagi-Lopes, L.C.; Vieira, J.M.; Sassaki, G.L.; Barreto-Bergter, E. Structural analysis of glucosylceramides (GlcCer) from species of the pseudallescheria/scedosporium complex. Fungal Biol. 2016, 120, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Marinho, P.R.; Muricy, G.R.S.; Silva, M.F.L.; de Marval, M.G.; Laport, M.S. Antibiotic-resistant bacteria inhibited by extracts and fractions from brazilian marine sponges. Rev. Bras. Farmacogn. 2010, 20, 267–275. [Google Scholar] [CrossRef]
- Hili, P.; Evans, C.S.; Veness, R.G. Antimicrobial action of essential oils: The effect of dimethylsulphoxide on the activity of cinnamon oil. Lett. Appl. Microbiol. 1997, 24, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds monohexosylceramides (CMH) are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rhizopus Species | Bacteria Species | Inhibition (+) or No Inhibition (−) | ||
|---|---|---|---|---|
| Control | Folch Lower Layer | CMH | ||
| R. stolonifer | B. cepacea | − | − | − |
| B. terrea | − | ± | + | |
| E. coli | − | − | − | |
| M. luteus | − | ± | + | |
| S. aureus | − | − | − | |
| S. aureus MRSA | − | − | − | |
| P. stutzeri | − | + | + | |
| R. microsporus | B. cepacea | − | − | − |
| B. terrea | − | + | ± | |
| E. coli | − | − | − | |
| M. luteus | − | ± | ± | |
| S. aureus | − | − | − | |
| S. aureus MRSA | − | − | − | |
| P. stutzeri | − | + | + | |
| CMH Fraction | Bacteria | MIC50 | MBC |
|---|---|---|---|
| R. stolonifer | B. terrea | 6.25 mg/mL | >50 mg/mL |
| M. luteus | 6.25 mg/mL | 25 mg/mL | |
| P. stutzeri | 3.13 mg/mL | 6.25 mg/mL | |
| R. microsporus | B. terrea | 3.13 mg/mL | >50 mg/mL |
| M. luteus | 6.25 mg/mL | 25 mg/mL | |
| P. stutzeri | 3.12 mg/mL | 6.25 mg/mL | |
| Streptomycin/Penicillin | B. terrea | 0.004 mg/mL | - |
| M. luteus | 0.004 mg/mL | - | |
| P. stutzeri | 0.004 mg/mL | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vieira, E.R.; Xisto, M.I.D.d.S.; Pele, M.A.; Alviano, D.S.; Alviano, C.S.; Barreto-Bergter, E.; De Campos-Takaki, G.M. Monohexosylceramides from Rhizopus Species Isolated from Brazilian Caatinga: Chemical Characterization and Evaluation of Their Anti-Biofilm and Antibacterial Activities. Molecules 2018, 23, 1331. https://doi.org/10.3390/molecules23061331
Vieira ER, Xisto MIDdS, Pele MA, Alviano DS, Alviano CS, Barreto-Bergter E, De Campos-Takaki GM. Monohexosylceramides from Rhizopus Species Isolated from Brazilian Caatinga: Chemical Characterization and Evaluation of Their Anti-Biofilm and Antibacterial Activities. Molecules. 2018; 23(6):1331. https://doi.org/10.3390/molecules23061331
Chicago/Turabian StyleVieira, Edson Rodrigues, Mariana Ingrid Dutra da Silva Xisto, Milagre Américo Pele, Daniela Sales Alviano, Celuta Sales Alviano, Eliana Barreto-Bergter, and Galba Maria De Campos-Takaki. 2018. "Monohexosylceramides from Rhizopus Species Isolated from Brazilian Caatinga: Chemical Characterization and Evaluation of Their Anti-Biofilm and Antibacterial Activities" Molecules 23, no. 6: 1331. https://doi.org/10.3390/molecules23061331