Cloning, Characterization, and Expression Analysis of Three FAD8 Genes Encoding a Fatty Acid Desaturase from Seeds of Paeonia ostii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Extraction of Seed Oil and Measurement by GC-MS
2.3. Cloning of Full-Length PoFAD8-1, PoFAD8-2, and PoFAD8-3 cDNA
2.4. Bioinformatic Analysis
2.5. Protein Subcellular Localization
2.6. Gene Expression Analysis
2.7. In Vitro Expression of FAD8-1 through Cell-Free Expression System
3. Results
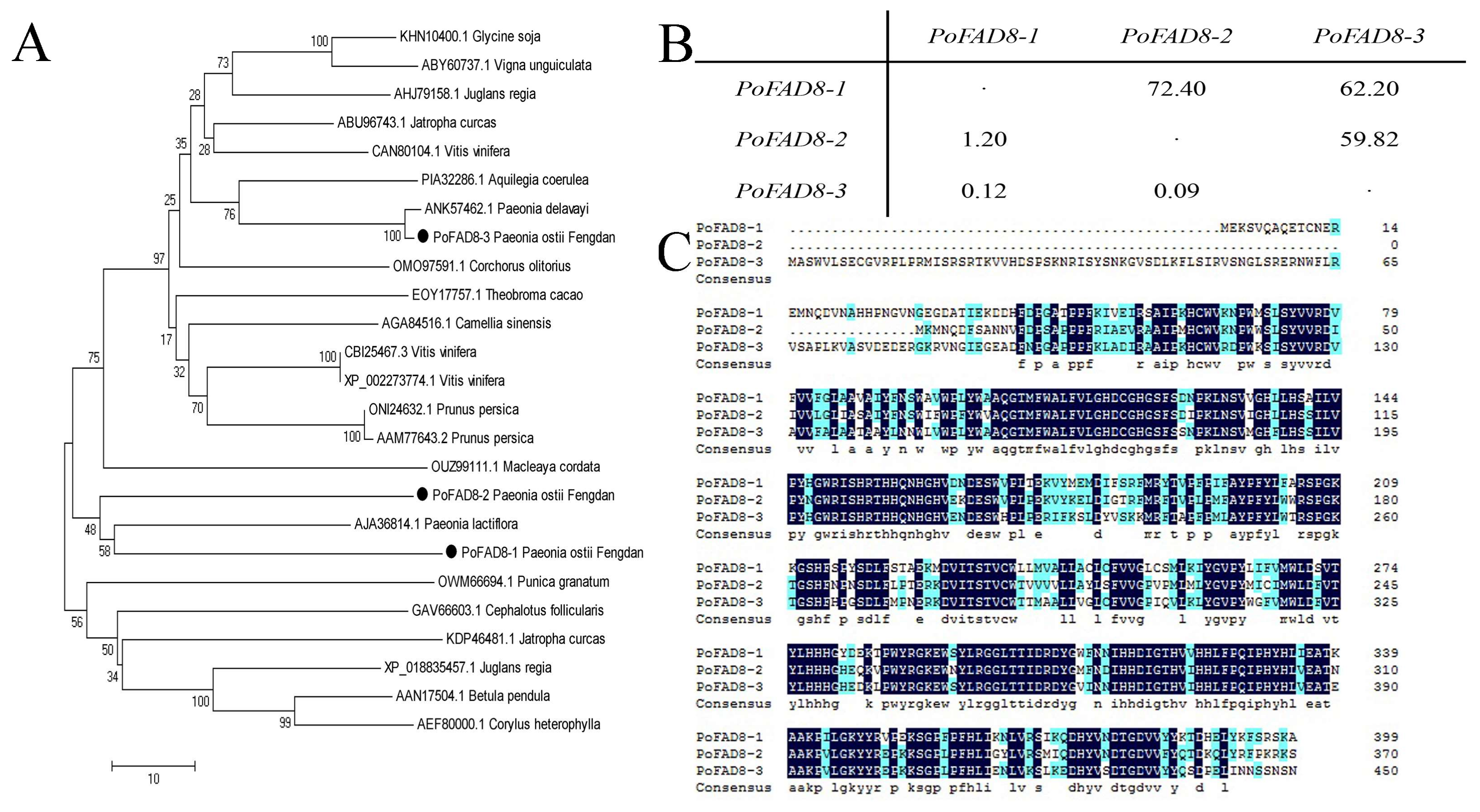
3.1. Cloning and Nucleotide Sequence Analysis of PoFAD8-1, PoFAD8-2, and PoFAD8-3
3.2. Bioinformatic Analysis of the PoFAD8-1, PoFAD8-2, and PoFAD8-3 Protein
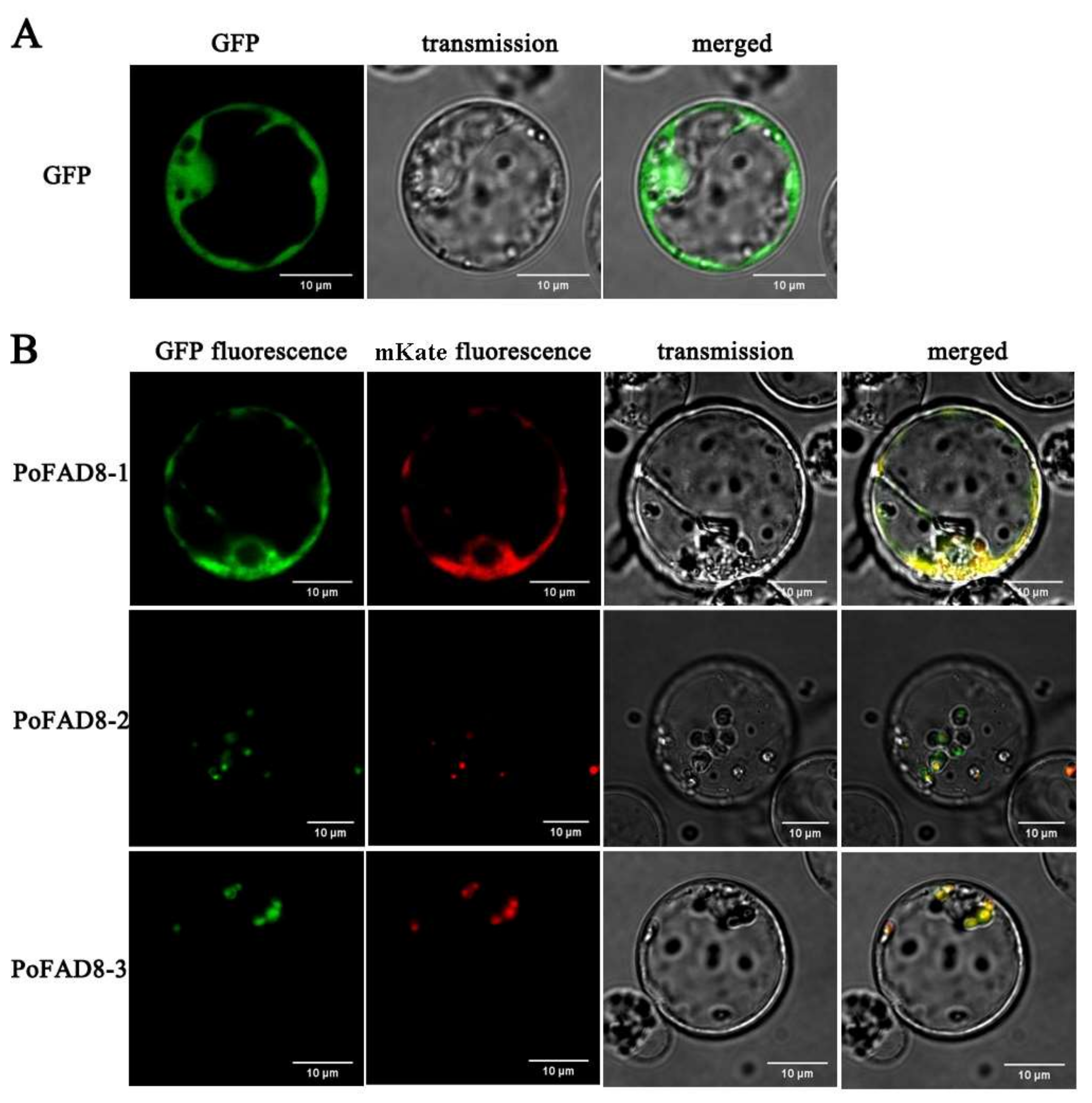
3.3. Subcellular Localization of the PoFAD8-1, PoFAD8-2, and PoFAD8-3 Protein
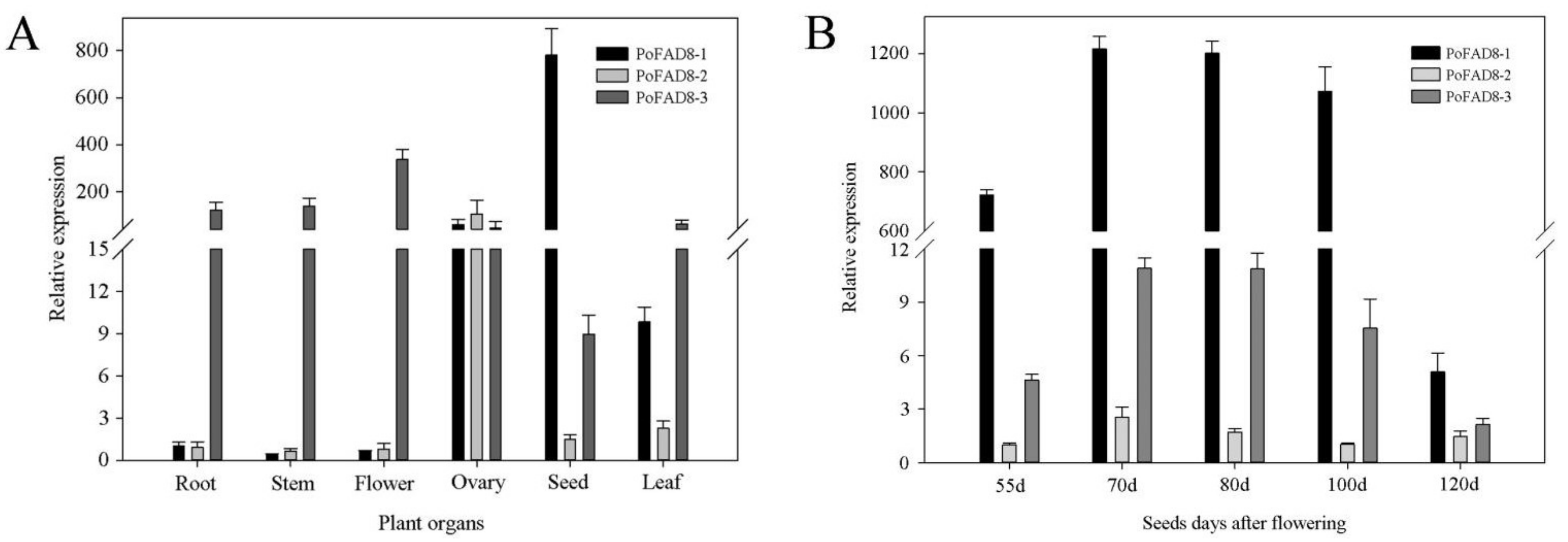
3.4. Expression Pattern of the PoFAD8-1, PoFAD8-2, and PoFAD8-3 Genes
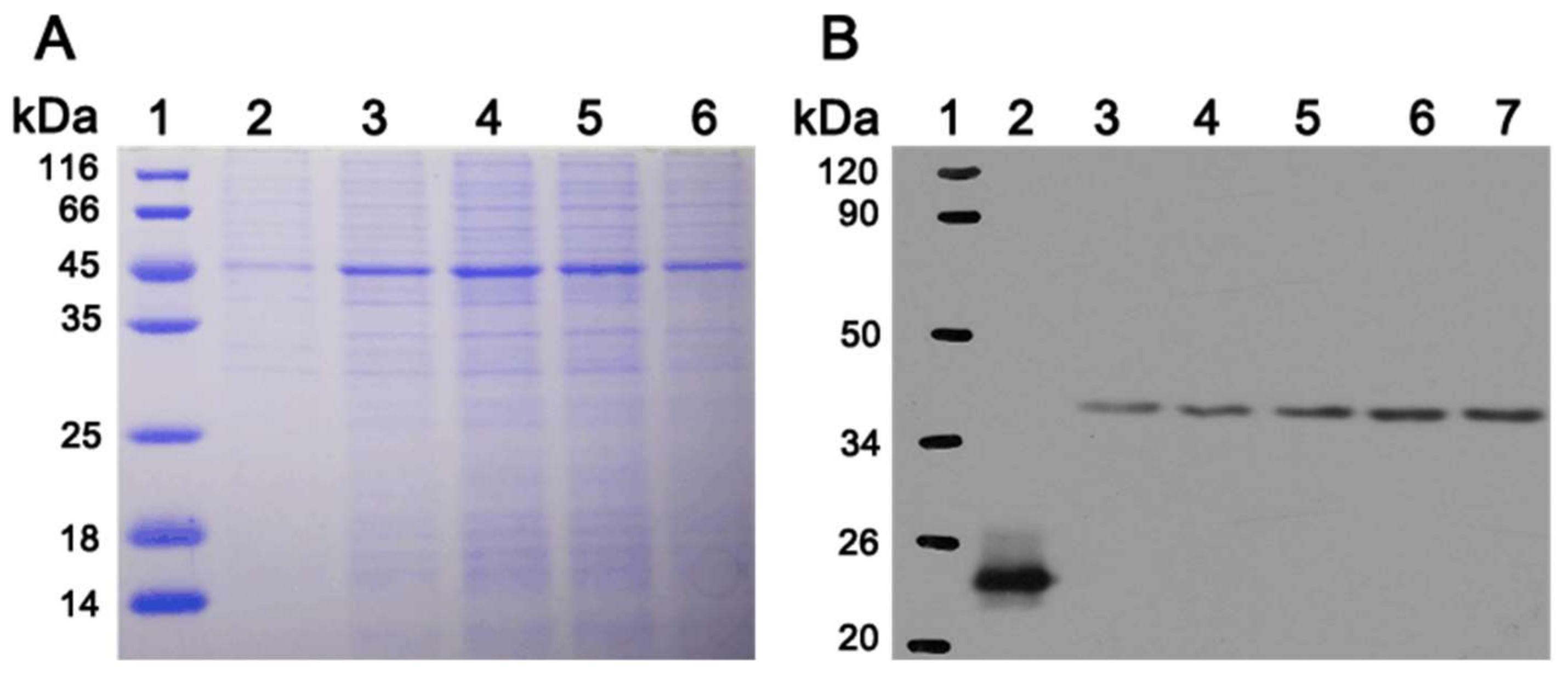
3.5. In Vitro Expression of PoFAD8-1 Gene
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cao, F.L.; Wang, H.L.; Yu, W.W.; Cheng, H. Advances in fatty acid desaturase and genes of higher plants. J. Nanjing For. Univ. Natl. Sci. Ed. 2012, 36, 125–132. [Google Scholar]
- Yazawa, H.; Iwahashi, H.; Kamisaka, Y.; Kimura, K.; Aki, T.; Ono, K.; Uemura, H. Heterologous production of dihomo-gamma-linolenic acid in Saccharomyces cerevisiae. Appl. Environ. Microb. 2007, 73, 6965–6971. [Google Scholar] [CrossRef] [PubMed]
- Ailhaud, G.; Massiera, F.; Weill, P.; Legrand, P.; Alessandri, J.M.; Guesnet, P. Temporal changes in dietary fats: Role of n − 6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog. Lipid Res. 2006, 45, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Teng, Y.B.; Li, B.; Meng, Q.W. Enhancement of low-temperature tolerance in transgenic tomato plants overexpressing Lefad7, through regulation of trienoic fatty acids. Photosynthetica 2013, 51, 238–244. [Google Scholar] [CrossRef]
- Wang, H.S.; Yu, C.; Tang, X.F.; Zhu, Z.J.; Ma, N.N.; Meng, Q.W. A tomato endoplasmic reticulum (ER)-type omega-3 fatty acid desaturase (LeFAD3) functions in early seedling tolerance to salinity stress. Plant Cell Rep. 2014, 33, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Tian, E.; Zeng, F.; Mackay, K.; Roslinsky, V.; Cheng, B. Detection and molecular characterization of two fad3 genes controlling linolenic acid content and development of allele-specific markers in yellow mustard (sinapis alba). PLoS ONE 2014, 9, e97430. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.; Arondel, V.; Iba, K.; Somerville, C. Cloning of a temperature-regulated gene encoding a chloroplast omega-3 desaturase from Arabidopsis thaliana. Plant Physiol. 1994, 106, 1615–1621. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.X.; Tang, G.Y.; Chen, G.; Bi, Y.P.; Shan, L. Polymorphism of AhFAD2 gene in peanut and its correlation with grain oleic acid/linoleic acid ratio. Acta Agron. Sin. 2011, 37, 415–423. [Google Scholar] [CrossRef]
- Somerville, C. Direct tests of the role of membrane lipid composition in low-temperature-induced photoinhibition and chilling sensitivity in plants and cyanobacteria. Proc. Natl. Acad. Sci. USA 1995, 92, 6215–6218. [Google Scholar] [CrossRef] [PubMed]
- Iba, K. Trienoic fatty acids and temperature tolerance of higher plants. In Abiotic Stress Tolerance in Plants; Springer Netherlands: Dordrecht, The Netherlands, 2006; pp. 61–68. [Google Scholar]
- Browse, J. The power of mutants for investigating jasmonate biosynthesis and signaling. Phytochemistry 2009, 70, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; Lapchyk, L.; Fukushige, H.; Hildebrand, D.; Klessig, D.; Kachroo, P. Plastidial fatty acid signaling modulates salicylic acid- and jasmonic acid-mediated defense pathways in the arabidopsis ssi2 mutant. Plant Cell 2003, 15, 2952–2965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Barg, R.; Yin, M.G.; Gueta-Dahan, Y.; Leikin-Frenkel, A.; Salts, Y.; Shabtai, S.; Ben-Hayyim, G. Modulated fatty acid desaturation via overexpression of two distinct omega-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Berberich, T.; Harada, M.; Sugawara, K.; Kodama, H.; Iba, K.; Kusano, T. Two maize genes encoding omega-3 fatty acid desaturase and their differential expression to temperature. Plant Mol. Biol. 1998, 36, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: Nutritional implications for chronic diseases. Biomed. Pharmacother. 2006, 60, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Wang, L.S.; Shu, Q.Y.; Wu, J.; Chen, L.G.; Shao, S.; Yin, D.D. Fatty acid composition of developing tree peony (paeonia section moutan DC.) seeds and transcriptome analysis during seed development. BMC Genom. 2015, 16, 208. [Google Scholar] [CrossRef] [PubMed]
- Ning, C.L.; Jiang, Y.; Meng, J.S.; Zhou, C.H.; Tao, J. Herbaceous peony seed oil: A rich source of unsaturated fatty acids and γ-tocopherol. Eur. J. Lipid Sci. Technol. 2015, 117, 532–542. [Google Scholar] [CrossRef]
- Biedermann, M.; Grob, K. Comprehensive two-dimensional gas chromatography for characterizing mineral oils in foods and distinguishing them from synthetic hydrocarbons. J. Chromatogr. A 2015, 1375, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Mravec, J.; Skupa, P.; Bailly, A.; Hoyerova, K.; Krecek, P.; Bielach, A.; Petrasek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.D. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, O.P.; Park, S.; Ilut, D.C.; Inmon, J.J.; Millhollon, J.C.; Liechty, Z.; Page, J.T.; Jenks, M.A.; Chapman, K.D.; Udall, J.A.; et al. Genome-wide analysis of the omega-3 fatty acid desaturase gene family in Gossypium. BMC Plant Biol. 2014, 14, 312. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, O.; Sakamoto, H.; Hashimoto, T.; Iba, K. A temperature-sensitive mechanism that regulates post-translational stability of a plastidial omega-3 fatty acid desaturase (FAD8) in Arabidopsis leaf tissues. J. Biol. Chem. 2005, 280, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Román, Á.; Andreu, V.; Hernández, M.L.; Lagunas, B.; Picorel, R.; Martínez-Rivas, J.M.; Alfonso, M. Contribution of the different omega-3 fatty acid desaturase genes to the cold response in soybean. J. Exp. Bot. 2012, 63, 4973–4982. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Ye, J.; Guo, W.D.; Wang, C.C.; Hu, H.T. Differences in cold tolerance and expression of two fatty acid desaturase genes in the leaves between fingered citron and its dwarf mutant. Trees 2012, 26, 1193–1201. [Google Scholar] [CrossRef]
- Iba, K.; Gibson, S.; Nishiuchi, T.; Fuse, T.; Nishimura, M.; Arondel, V.; Hugly, S.; Somerville, C. A gene encoding a chloroplast omega-3 fatty acid desaturase complements alterations in fatty acid desaturation and chloroplast copy number of the fad7 mutant of Arabidopsis thaliana. J. Biol. Chem. 1993, 268, 24099–24105. [Google Scholar] [PubMed]
- Nguyen, H.M.; Cuiné, S.; Beyly-Adriano, A.; Légeret, B.; Billon, E.; Auroy, P.; Beisson, F.; Peltier, G.; Li-Beisson, Y. The green microalga Chlamydomonas reinhardtii has a single ω-3 fatty acid desaturase that localizes to the chloroplast and impacts both plastidic and extraplastidic membrane lipids. Plant Physiol. 2013, 163, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.U.; Lee, K.R.; Shim, D.; Lee, J.H.; Chen, G.Q.; Hwang, S. Transcriptome analysis and identification of genes associated with ω-3 fatty acid biosynthesis in Perilla frutescens(L.) var. frutescens. BMC Genom. 2016, 17, 474. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Chen, B.; Win, A.N.; Fu, C.; Lian, J.; Liu, X.; Wang, R.; Zhang, X.; Chai, Y. Omega-3 fatty acid desaturase gene family from two ω-3 sources, Salvia hispanica and Perilla frutescens: Cloning, characterization and expression. PLoS ONE 2018, 13, e0191432. [Google Scholar] [CrossRef] [PubMed]
- Dyer, J.M.; Chapital, D.C.; Cary, J.W.; Pepperman, A.B. Chilling-sensitive, post-transcriptional regulation of a plant fatty acid desaturase expressed in yeast. Biochem. Biophs. Res. Commun. 2001, 282, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Ubiquitin | GACCTATACCAAGCCGAAG | CGTTCCAGCACCACAATC |
| qFAD8-1 | ACCTTAGAGGAGGGCTTACGACAAT | CGTAGTGGTCTTGCTTGATGCTCCT |
| qFAD8-2 | TAGTGGCATGAAGATGAATCAAGAT | CGCTTGCTATTAGTCCCAGAACCAC |
| qFAD8-3 | ATGATAGCCCCTCAAAGAATAGAAT | CGCAACATCCCTCACAACATAGC |
| Amino Acids | PoFAD8-1 | PoFAD8-2 | PoFAD8-3 | Amino Acids | PoFAD8-1 | PoFAD8-2 | PoFAD8-3 |
|---|---|---|---|---|---|---|---|
| Ala (A) | 22 | 14 | 24 | Leu (L) | 30 | 31 | 40 |
| Arg (R) | 14 | 16 | 24 | Lys (K) | 22 | 17 | 24 |
| Asn (N) | 15 | 14 | 20 | Met (M) | 11 | 12 | 9 |
| Asp (D) | 22 | 19 | 22 | Phe (F) | 23 | 22 | 20 |
| Cys (C) | 7 | 4 | 5 | Pro (P) | 23 | 28 | 31 |
| Gln (Q) | 7 | 8 | 5 | Ser (S) | 25 | 21 | 38 |
| Glu (E) | 17 | 10 | 18 | Thr (T) | 18 | 16 | 17 |
| Gly (G) | 25 | 23 | 30 | Trp (W) | 13 | 15 | 16 |
| His (H) | 28 | 24 | 28 | Tyr (Y) | 23 | 21 | 20 |
| Ile (I) | 20 | 22 | 20 | Val (V) | 35 | 34 | 39 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, J.; Chen, M.; Zhu, M.; Jiang, Y.; Meng, J.; Zhao, D.; Tao, J. Cloning, Characterization, and Expression Analysis of Three FAD8 Genes Encoding a Fatty Acid Desaturase from Seeds of Paeonia ostii. Molecules 2018, 23, 929. https://doi.org/10.3390/molecules23040929
Sun J, Chen M, Zhu M, Jiang Y, Meng J, Zhao D, Tao J. Cloning, Characterization, and Expression Analysis of Three FAD8 Genes Encoding a Fatty Acid Desaturase from Seeds of Paeonia ostii. Molecules. 2018; 23(4):929. https://doi.org/10.3390/molecules23040929
Chicago/Turabian StyleSun, Jing, Ming Chen, Mengyuan Zhu, Yu Jiang, Jiasong Meng, Daqiu Zhao, and Jun Tao. 2018. "Cloning, Characterization, and Expression Analysis of Three FAD8 Genes Encoding a Fatty Acid Desaturase from Seeds of Paeonia ostii" Molecules 23, no. 4: 929. https://doi.org/10.3390/molecules23040929