
Curcumin Inhibits LIN-28A through the Activation of miRNA-98 in the Lung Cancer Cell Line A549

 , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
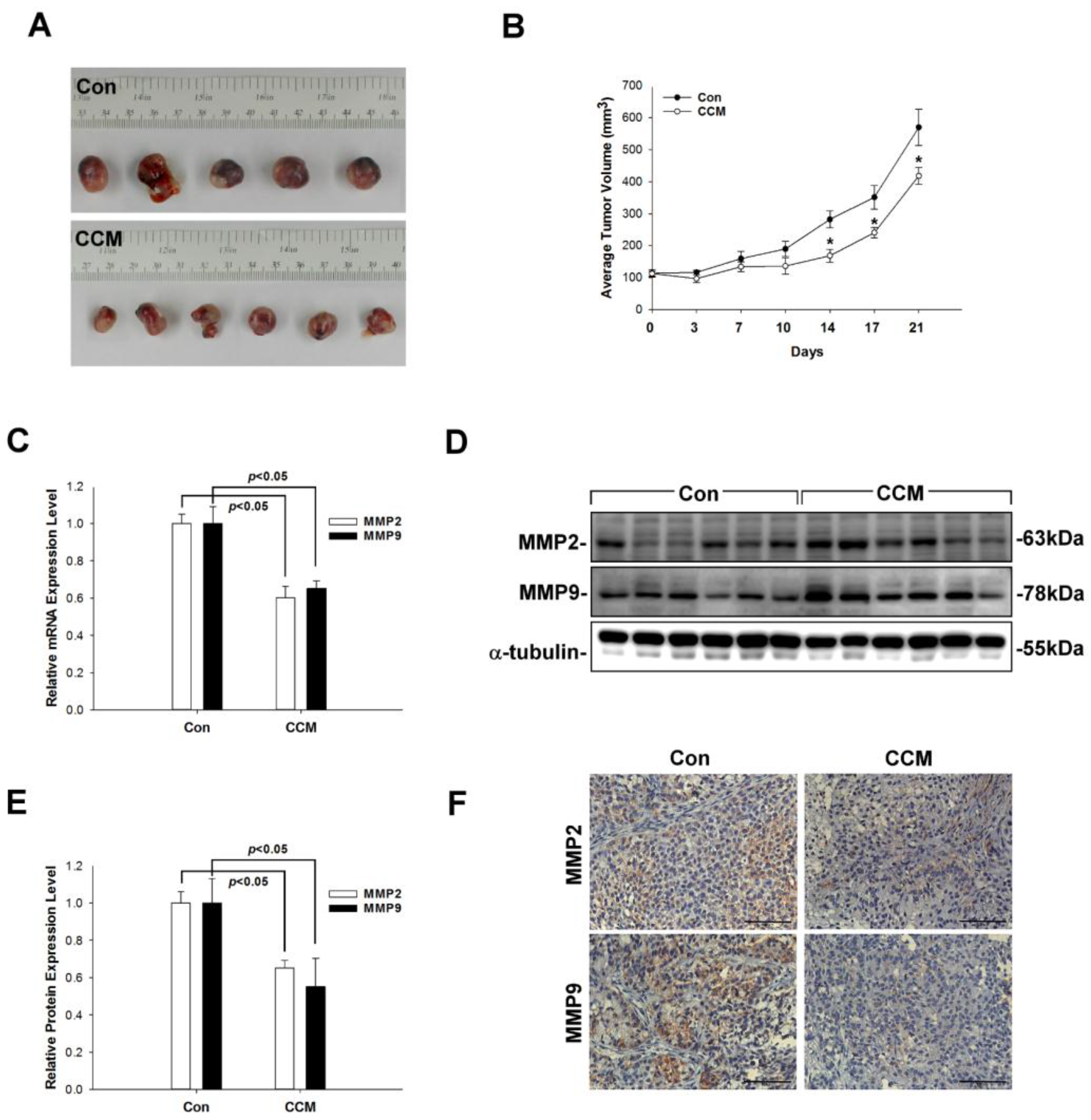
2.1. Curcumin Suppresses MMP2/9 Levels and Lung Cancer Growth In Vivo
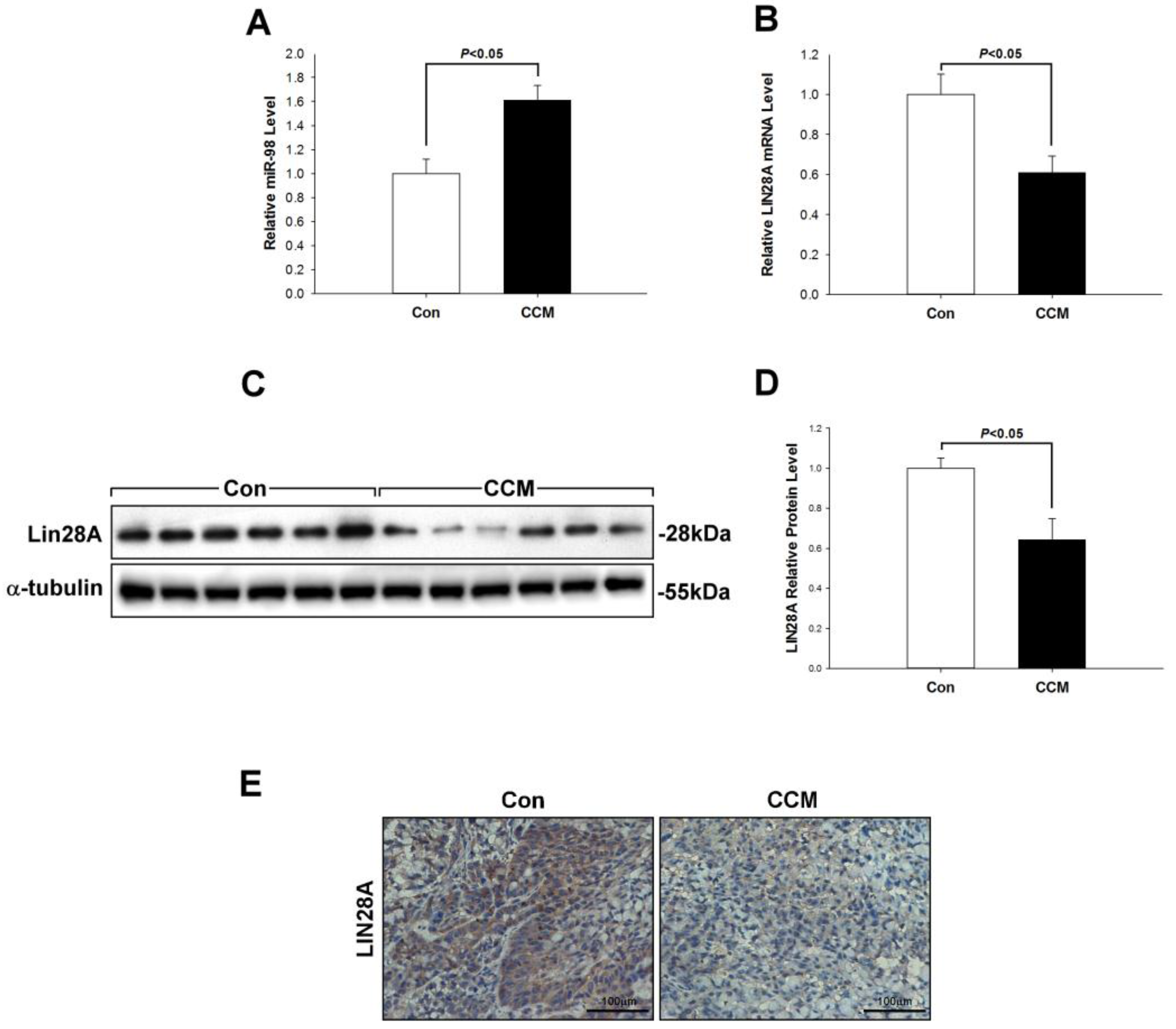
2.2. Curcumin Induces miR-98 Upregulation and LIN28A Downregulation In Vivo
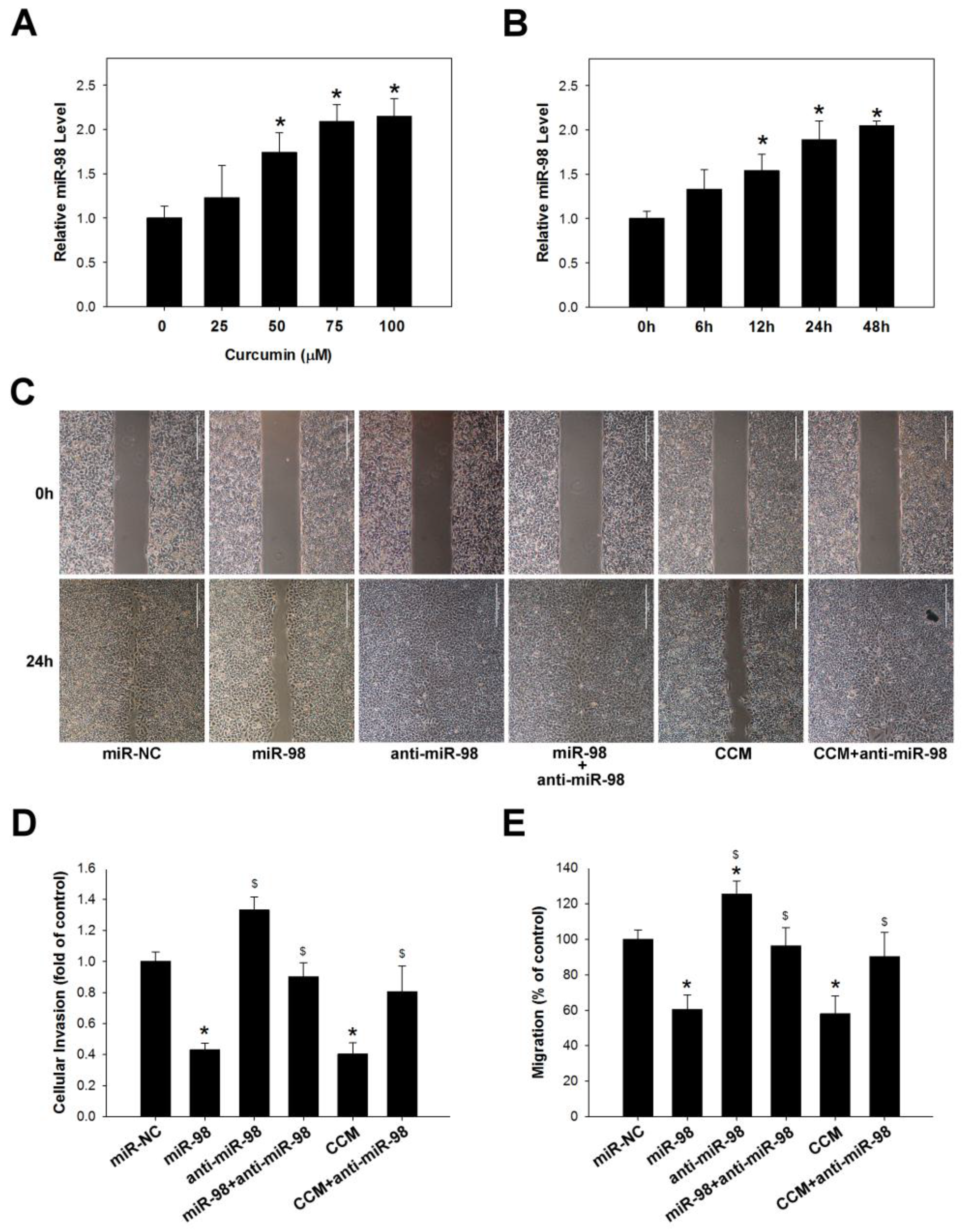
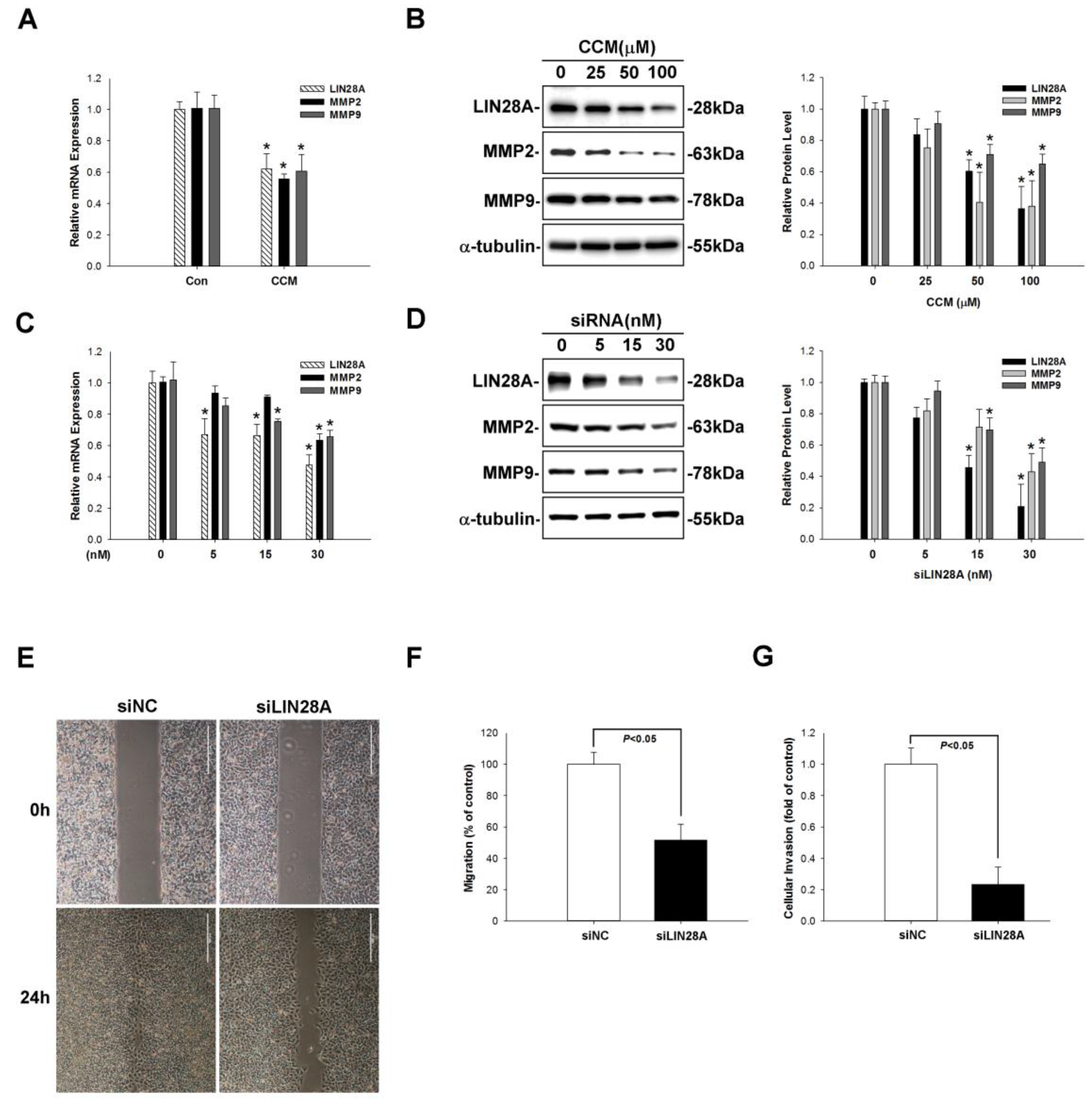
2.3. Curcumin Inhibits Lung Cancer Cell Migration and Invasion via Upregulation of miR-98
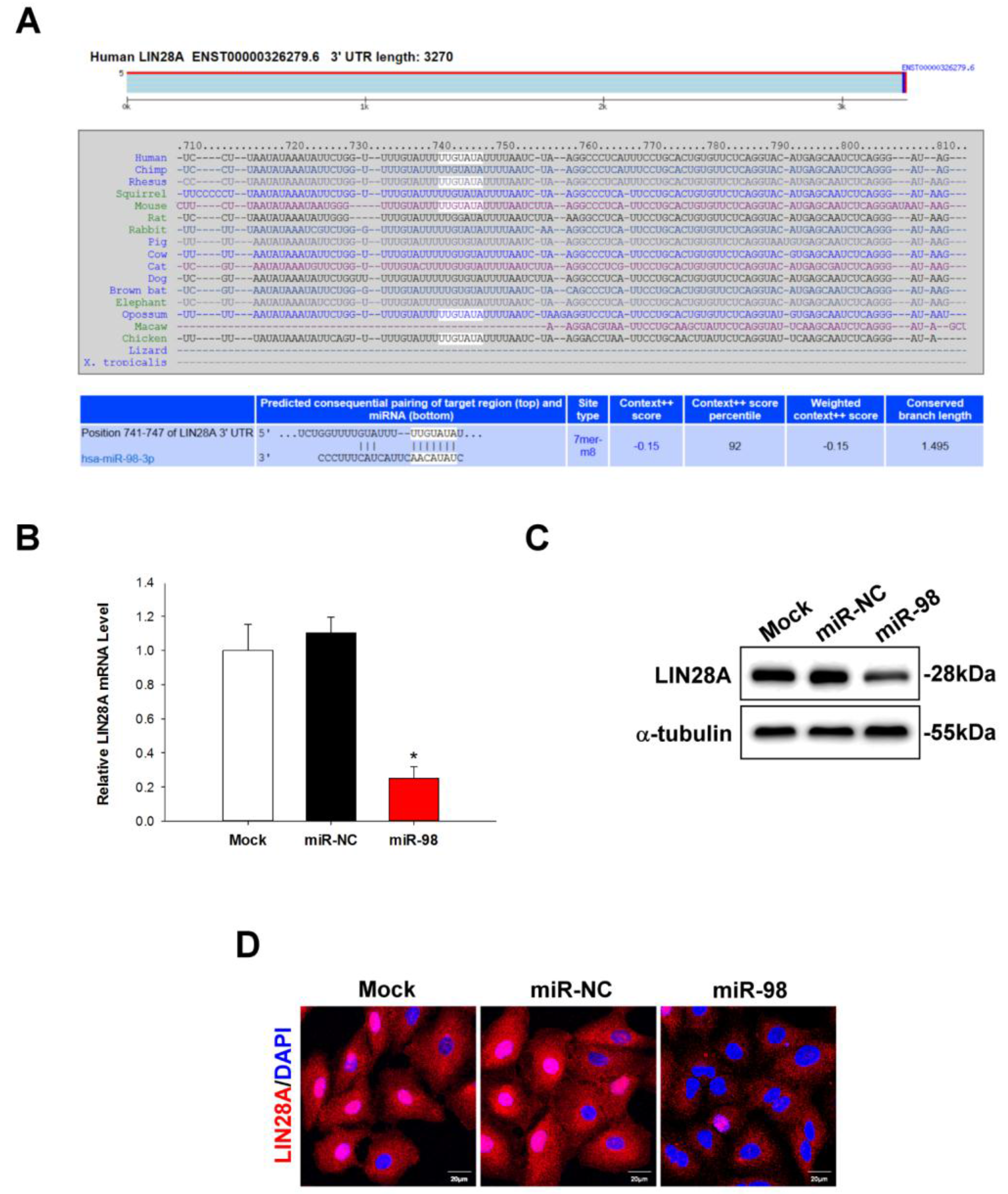
2.4. LIN28A Is a Downstream Target of miR-98 in Human Lung Cancer Cells
2.5. Curcumin Inhibits LIN28A-Mediated MMP2/9 Expression and Lung Cancer Metastasis
3. Discussion
4. Materials and Methods
4.1. Culture of A549 Cells
4.2. Immunohistochemical Staining
4.3. Immunocytochemistry
4.4. Migration Analysis
4.5. Cell Invasion Assay
4.6. Quantitative Real-Time PCR
4.7. Cell Transfection with miRNA-98 Mimic and Inhibitor
4.8. Western Blotting
4.9. Animal Study
4.10. Statistical Analyses
5. Conclusions
Acknowledgments
Author contributions
Conflicts of Interest
References
- Crawford, S.E.; Stellmach, V.; Ranalli, M.; Huang, X.; Huang, L.; Volpert, O.; De Vries, G.H.; Abramson, L.P.; Bouck, N. Pigment epithelium-derived factor (PEDF) in neuroblastoma: A multifunctional mediator of Schwann cell antitumor activity. J. Cell Sci. 2001, 114, 4421–4428. [Google Scholar] [PubMed]
- Jacob, A.; Prekeris, R. The regulation of MMP targeting to invadopodia during cancer metastasis. Front. Cell Dev. Biol. 2015, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lee, C.G. MicroRNA and cancer-focus on apoptosis. J. Cell. Mol. Med. 2009, 13, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Peters, S.; Bexelius, C.; Munk, V.; Leighl, N. The impact of brain metastasis on quality of life, resource utilization and survival in patients with non-small-cell lung cancer. Cancer Treat. Rev. 2016, 45, 139–162. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Wu, Y.; Zhang, Q.; Gao, G.; Zhang, C.; Xiang, Y. Identification of four serum microRNAs from a genome-wide serum microRNA expression profile as potential non-invasive biomarkers for endometrioid endometrial cancer. Oncol. Lett. 2013, 6, 261–267. [Google Scholar] [PubMed]
- Xiang, Q.; Tang, H.; Yu, J.; Yin, J.; Yang, X.; Lei, X. MicroRNA-98 sensitizes cisplatin-resistant human lung adenocarcinoma cells by up-regulation of HMGA2. Pharmazie 2013, 68, 274–281. [Google Scholar] [PubMed]
- Zhu, W.; Huang, Y.; Pan, Q.; Xiang, P.; Xie, N.; Yu, H. MicroRNA-98 Suppress Warburg Effect by Targeting HK2 in Colon Cancer Cells. Dig. Dis. Sci. 2017, 62, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Schageman, J.J.; Subauste, M.C.; Saber, B.; Hammond, S.M.; Prudkin, L.; Wistuba, II.; Ji, L.; Roth, J.A.; Minna, J.D.; et al. miR-93, miR-98, and miR-197 regulate expression of tumor suppressor gene FUS1. Mol. Cancer Res. 2009, 7, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Wendler, A.; Keller, D.; Albrecht, C.; Peluso, J.J.; Wehling, M. Involvement of let-7/miR-98 microRNAs in the regulation of progesterone receptor membrane component 1 expression in ovarian cancer cells. Oncol. Rep. 2010, 25, 273–279. [Google Scholar] [CrossRef]
- Du, Y.; Li, Y.; Lv, H.; Zhou, S.; Sun, Z.; Wang, M. miR-98 suppresses tumor cell growth and metastasis by targeting IGF1R in oral squamous cell carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 12252. [Google Scholar] [PubMed]
- Ting, H.J.; Messing, J.; Yasmin-Karim, S.; Lee, Y.F. Identification of microRNA-98 as a therapeutic target inhibiting prostate cancer growth and a biomarker induced by vitamin D. J. Biol. Chem. 2013, 288, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Hou, L.; Huang, Y. EZH2-specific microRNA-98 inhibits human ovarian cancer stem cell proliferation via regulating the pRb-E2F pathway. Tumour Biol. 2014, 35, 7239–7247. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.Y.; Chen, Q.J.; Wei, Y.; Wang, Y.L.; Wang, Z.W.; Xu, K.; He, Y.; Ma, H.B. Upregulation of microRNA-98 increases radiosensitivity in esophageal squamous cell carcinoma. J. Radiat. Res. 2016, 57, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-D.; Yuan, Y.; Zhuang, C.-W.; Li, B.-L.; Gong, D.-J.; Wang, S.-G.; Zeng, Z.-Y.; Cheng, H.-Z. MicroRNA-98 and microRNA-214 post-transcriptionally regulate enhancer of zeste homolog 2 and inhibit migration and invasion in human esophageal squamous cell carcinoma. Mol. Cancer 2012, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Zou, B.; Liu, L.; Cui, K.; Gao, J.; Yuan, S.; Cong, N. MicroRNA-98 acts as a tumor suppressor in hepatocellular carcinoma via targeting SALL4. Oncotarget 2016, 7, 74059–74073. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, X.; Shi, J. MiR-98 inhibits cell proliferation and invasion of non-small cell carcinoma lung cancer by targeting PAK1. Int. J. Clin. Exp. Med. 2015, 8, 20135. [Google Scholar] [PubMed]
- Sterenczak, K.A.; Eckardt, A.; Kampmann, A.; Willenbrock, S.; Eberle, N.; Länger, F.; Kleinschmidt, S.; Hewicker-Trautwein, M.; Kreipe, H.; Nolte, I. HMGA1 and HMGA2 expression and comparative analyses of HMGA2, Lin28 and let-7 miRNAs in oral squamous cell carcinoma. BMC Cancer 2014, 14, 694. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zhao, J.; Hu, W.; Yang, G.; Yu, H.; Wang, R.; Wang, L.; Zhang, G.; Fu, W.; Dai, L.; et al. Disturbance of the let-7/LIN28 double-negative feedback loop is associated with radio- and chemo-resistance in non-small cell lung cancer. PLoS ONE 2017, 12, e0172787. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.R.; Daley, G.Q. Lin28: A microRNA regulator with a macro role. Cell 2010, 140, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Parasramka, M.A.; Dashwood, W.M.; Wang, R.; Abdelli, A.; Bailey, G.S.; Williams, D.E.; Ho, E.; Dashwood, R.H. MicroRNA profiling of carcinogen-induced rat colon tumors and the influence of dietary spinach. Mol. Nutr. Food Res. 2012, 56, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Newman, M.A.; Thomson, J.M.; Hammond, S.M. Lin-28 interaction with the Let-7 precursor loop mediates regulated microRNA processing. RNA 2008, 14, 1539–1549. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.X.; Zhang, D.Z.; Fang, X.J.; Sun, P.S.; Xue, H.C. Let-7a mimic attenuates CCL18 induced breast cancer cell metastasis through Lin 28 pathway. Biomed. Pharmacother. 2016, 78, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.H.; Zhu, H. Lin28 and let-7 in cell metabolism and cancer. Transl. Pediatr. 2015, 4, 4–11. [Google Scholar] [PubMed]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [PubMed]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell. 2006, 9, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Coban, D.; Milenkovic, D.; Chanet, A.; Khallou-Laschet, J.; Sabbe, L.; Palagani, A.; Vanden Berghe, W.; Mazur, A.; Morand, C. Dietary curcumin inhibits atherosclerosis by affecting the expression of genes involved in leukocyte adhesion and transendothelial migration. Mol. Nutr. Food Res. 2012, 56, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Yuan, W.; Li, S.; Gupta, S.C. Curcumin-free turmeric exhibits anti-inflammatory and anticancer activities: Identification of novel components of turmeric. Mol. Nutr. Food Res. 2013, 57, 1529–1542. [Google Scholar] [CrossRef] [PubMed]
- López-Lázaro, M. Anticancer and carcinogenic properties of curcumin: Considerations for its clinical development as a cancer chemopreventive and chemotherapeutic agent. Mol. Nutr. Food Res. 2008, 52, S103–S127. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.F.; Fu, Y.S.; Liao, Y.J.; Xia, W.J.; Chen, Y.C.; Zeng, Y.X.; Kung, H.F.; Xie, D. Curcumin induces down-regulation of EZH2 expression through the MAPK pathway in MDA-MB-435 human breast cancer cells. Eur. J. Pharmacol. 2010, 637, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-Q.; Chai, K.-Q.; Zhu, X.-M.; Jiang, H.; Wang, X.; Xue, Q.; Zheng, A.-H.; Zhou, H.-Y.; Chen, Y.; Chen, X.-C. Anti-cancer effects of curcumin on lung cancer through the inhibition of EZH2 and NOTCH1. Oncotarget 2016, 7, 26535. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.R.; Liu, P.L.; Chen, Y.H.; Chou, S.H.; Cheng, Y.J.; Hwang, J.J.; Chong, I.W. Curcumin inhibits non-small cell lung cancer cells metastasis through the adiponectin/NF-kappab/MMPs signaling pathway. PLoS ONE 2015, 10, e0144462. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Pedone, C.; Majeed, M.; Sahebkar, A.H. Curcumin and lung cancer: The role of microRNAs. Curr. Pharm. Des. 2017, 23, 1–5. [Google Scholar]
- Bao, B.; Ali, S.; Banerjee, S.; Wang, Z.; Logna, F.; Azmi, A.S.; Kong, D.; Ahmad, A.; Li, Y.; Padhye, S.; et al. Curcumin analogue CDF inhibits pancreatic tumor growth by switching on suppressor microRNAs and attenuating EZH2 expression. Cancer Res. 2012, 72, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Bao, B.; Ali, S.; Ahmad, A.; Azmi, A.S.; Li, Y.; Banerjee, S.; Kong, D.; Sethi, S.; Aboukameel, A.; Padhye, S.B.; et al. Hypoxia-induced aggressiveness of pancreatic cancer cells is due to increased expression of VEGF, IL-6 and miR-21, which can be attenuated by CDF treatment. PLoS ONE 2012, 7, e50165. [Google Scholar] [CrossRef] [PubMed]
- Teiten, M.H.; Dicato, M.; Diederich, M. Curcumin as a regulator of epigenetic events. Mol. Nutr. Food Res. 2013, 57, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, V.A.; Cleys, E.R.; Da Silveira, J.C.; Spillman, M.A.; Winger, Q.A.; Bouma, G.J. High LIN28A Expressing Ovarian Cancer Cells Secrete Exosomes That Induce Invasion and Migration in HEK293 Cells. Biomed. Res. Int. 2015, 2015, 701390. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Xu, W.; Song, J.; Liang, Y.; Fu, W.; Zhu, X.C.; Li, C.; Peng, J.-S.; Zheng, J.-N. Overexpression of Lin28 inhibits the proliferation, migration and cell cycle progression and induces apoptosis of BGC-823 gastric cancer cells. Oncol. Rep. 2015, 33, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Fingleton, B. Matrix metalloproteinases: Roles in cancer and metastasis. Front. Biosci. 2005, 11, 479–491. [Google Scholar] [CrossRef]
- Wang, T.; He, Y.; Zhu, Y.; Chen, M.; Weng, M.; Yang, C.; Zhang, Y.; Ning, N.; Zhao, R.; Yang, W. Comparison of the expression and function of Lin28A and Lin28B in colon cancer. Oncotarget 2016, 7, 79605–79616. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Zhao, L.; Feng, X.; Xu, C.; Li, C.; Niu, Y. Lin28A activates androgen receptor via regulation of c-myc and promotes malignancy of ER−/Her2+ breast cancer. Oncotarget 2016, 7, 60407–60418. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Lv, H.; Wu, F.; Wang, C.; Li, T.; Lv, G.; Tang, L.; Guo, L.; Tang, S.; Cao, D.; et al. Musashi 2 contributes to the stemness and chemoresistance of liver cancer stem cells via LIN28A activation. Cancer Lett. 2017, 384, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Fu, T.Y.; Chang, C.C.; Lin, C.T.; Lai, C.H.; Peng, S.Y.; Ko, Y.J.; Tang, P.C. Let-7b-mediated suppression of basigin expression and metastasis in mouse melanoma cells. Exp. Cell Res. 2011, 317, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, S.; Shen, H.; Chen, W.; Xu, H.; Chen, X.; Sun, D.; Zhong, S.; Zhao, J.; Tang, J. Curcumin inhibits cancer progression through regulating expression of microRNAs. Tumour Biol. 2017, 39, 1010428317691680. [Google Scholar] [CrossRef] [PubMed]
- Tzatsos, A.; Paskaleva, P.; Lymperi, S.; Contino, G.; Stoykova, S.; Chen, Z.; Wong, K.K.; Bardeesy, N. Lysine-specific demethylase 2B (KDM2B)-let-7-enhancer of zester homolog 2 (EZH2) pathway regulates cell cycle progression and senescence in primary cells. J. Biol. Chem. 2011, 286, 33061–33069. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Tang, Y.; Cui, H.; Zhao, X.; Luo, X.; Pan, W.; Huang, X.; Shen, N. Let-7/miR-98 regulate Fas and Fas-mediated apoptosis. Genes Immun. 2011, 12, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Qi, J.; Reyes, J.M.; Li, L.; Rao, P.K.; Li, F.; Lin, C.Y.; Perry, J.A.; Lawlor, M.A.; Federation, A. Oncogenic deregulation of EZH2 as an opportunity for targeted therapy in lung cancer. Cancer Discov. 2016, 6, 1006–1021. [Google Scholar] [CrossRef] [PubMed]
- Wassef, M.; Rodilla, V.; Teissandier, A.; Zeitouni, B.; Gruel, N.; Sadacca, B.; Irondelle, M.; Charruel, M.; Ducos, B.; Michaud, A.; et al. Impaired PRC2 activity promotes transcriptional instability and favors breast tumorigenesis. Genes Dev. 2015, 29, 2547–2562. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.C.; Chen, Y.L.; Yuan, R.H.; Pan, H.W.; Yang, W.C.; Hsu, H.C.; Jeng, Y.M. Lin-28B expression promotes transformation and invasion in human hepatocellular carcinoma. Carcinogenesis 2010, 31, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; May, L.; Milsom, C.; Anderson, G.M.; Weitz, J.I.; Luyendyk, J.P.; Broze, G.; Mackman, N.; Rak, J. Contribution of host-derived tissue factor to tumor neovascularization. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1975–1981. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Zhang, J.J.; Hua, L.; Yao, K.H.; Chen, J.T.; Ren, X.Q. MicroRNA-98 suppresses cell proliferation, migration and invasion by targeting collagen triple helix repeat containing 1 in hepatocellular carcinoma. Mol. Med. Rep. 2016, 13, 2639–2644. [Google Scholar] [PubMed]
- Fan, Y.; Ye, M.; Wu, L.; Lv, S.; Wu, M.; Xiao, B.; Liao, C.; Ji, Q.; Chai, Y.; Zhu, X. Overexpression of miR-98 inhibits cell invasion in glioma cell lines via downregulation of IKKε. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3593–3604. [Google Scholar] [PubMed]
- Liu, W.; Xiao, P.; Wu, H.; Wang, L.; Kong, D.; Yu, F. MicroRNA-98 plays a suppressive role in non-small cell lung cancer through inhibition of SALL4 protein expression. Oncol. Res. 2016. [Google Scholar] [CrossRef]
- Abbas Momtazi, A.; Sahebkar, A. Difluorinated curcumin: A promising curcumin analogue with improved anti-tumor activity and pharmacokinetic profile. Curr. Pharm. Des. 2016, 22, 4386–4397. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Z.; Han, Z.; Tian, N. Paclitaxel-sensitization enhanced by curcumin involves down-regulation of nuclear factor-κ B and Lin28 in Hep3B cells. J. Recept. Signal. Transduct. Res. 2015, 35, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.A.; Jiang, W.G. Loss of tight junction barrier function and its role in cancer metastasis. Biochim. Biophys. Acta. 2009, 1788, 872–891. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.W.; Bernardo, M.M.; Pietila, M.; Gervasi, D.C.; Toth, M.; Kotra, L.P.; Massova, I.; Mobashery, S.; Fridman, R. Characterization of the monomeric and dimeric forms of latent and active matrix metalloproteinase-9. Differential rates for activation by stromelysin 1. J. Biol. Chem. 2000, 275, 2661–2668. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Bian, S.; Li, J.; He, J.; Chen, H.; Ge, L.; Jiao, Z.; Zhang, Y.; Peng, W.; Du, F.; Mo, Y.; Gong, A. MeCP2 suppresses LIN28A expression via binding to its methylated-CpG islands in pancreatic cancer cells. Oncotarget 2016, 7, 14476–14485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Chen, J.T.; Hua, L.; Yao, K.H.; Wang, C.Y. miR-98 inhibits hepatocellular carcinoma cell proliferation via targeting EZH2 and suppressing Wnt/beta-catenin signaling pathway. Biomed. Pharmacother. 2017, 85, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.F.; Liu, P.L.; Chong, I.W.; Liu, Y.P.; Chen, Y.H.; Ku, P.M.; Li, C.Y.; Chen, H.H.; Chiang, H.C.; Wang, C.L.; et al. Pigment Epithelium-Derived Factor Mediates Autophagy and Apoptosis in Myocardial Hypoxia/Reoxygenation Injury. PLoS ONE 2016, 11, e0156059. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the Curcumin compounds are not available from authors. |





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-L.; Chang, J.-M.; Chong, I.-W.; Hung, Y.-L.; Chen, Y.-H.; Huang, W.-T.; Kuo, H.-F.; Hsieh, C.-C.; Liu, P.-L. Curcumin Inhibits LIN-28A through the Activation of miRNA-98 in the Lung Cancer Cell Line A549. Molecules 2017, 22, 929. https://doi.org/10.3390/molecules22060929
Liu W-L, Chang J-M, Chong I-W, Hung Y-L, Chen Y-H, Huang W-T, Kuo H-F, Hsieh C-C, Liu P-L. Curcumin Inhibits LIN-28A through the Activation of miRNA-98 in the Lung Cancer Cell Line A549. Molecules. 2017; 22(6):929. https://doi.org/10.3390/molecules22060929
Chicago/Turabian StyleLiu, Wei-Lun, Jia-Ming Chang, Inn-Wen Chong, Yi-Li Hung, Yung-Hsiang Chen, Wen-Tsung Huang, Hsuan-Fu Kuo, Chong-Chao Hsieh, and Po-Len Liu. 2017. "Curcumin Inhibits LIN-28A through the Activation of miRNA-98 in the Lung Cancer Cell Line A549" Molecules 22, no. 6: 929. https://doi.org/10.3390/molecules22060929