
Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Fermentative Aroma Compounds
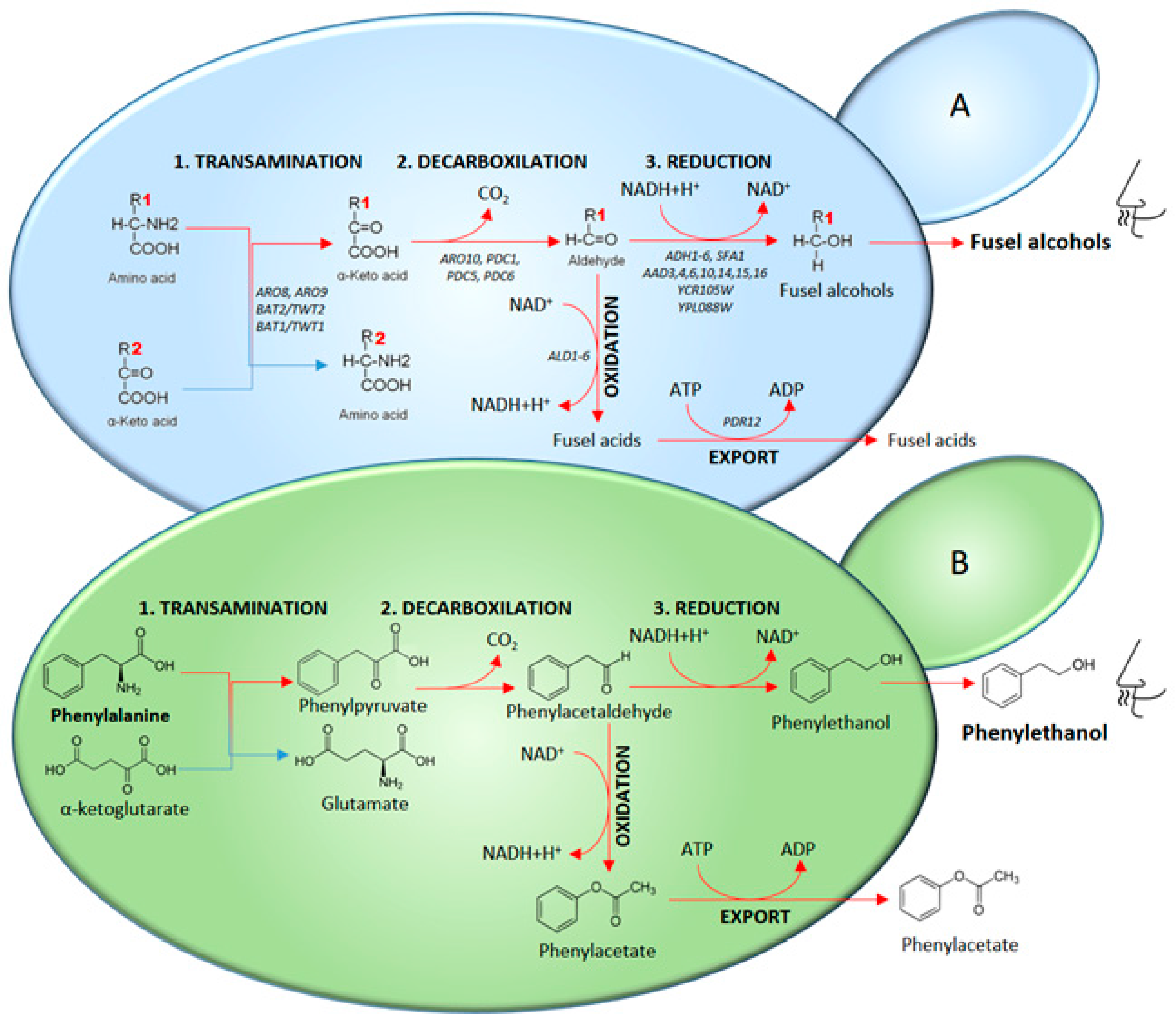
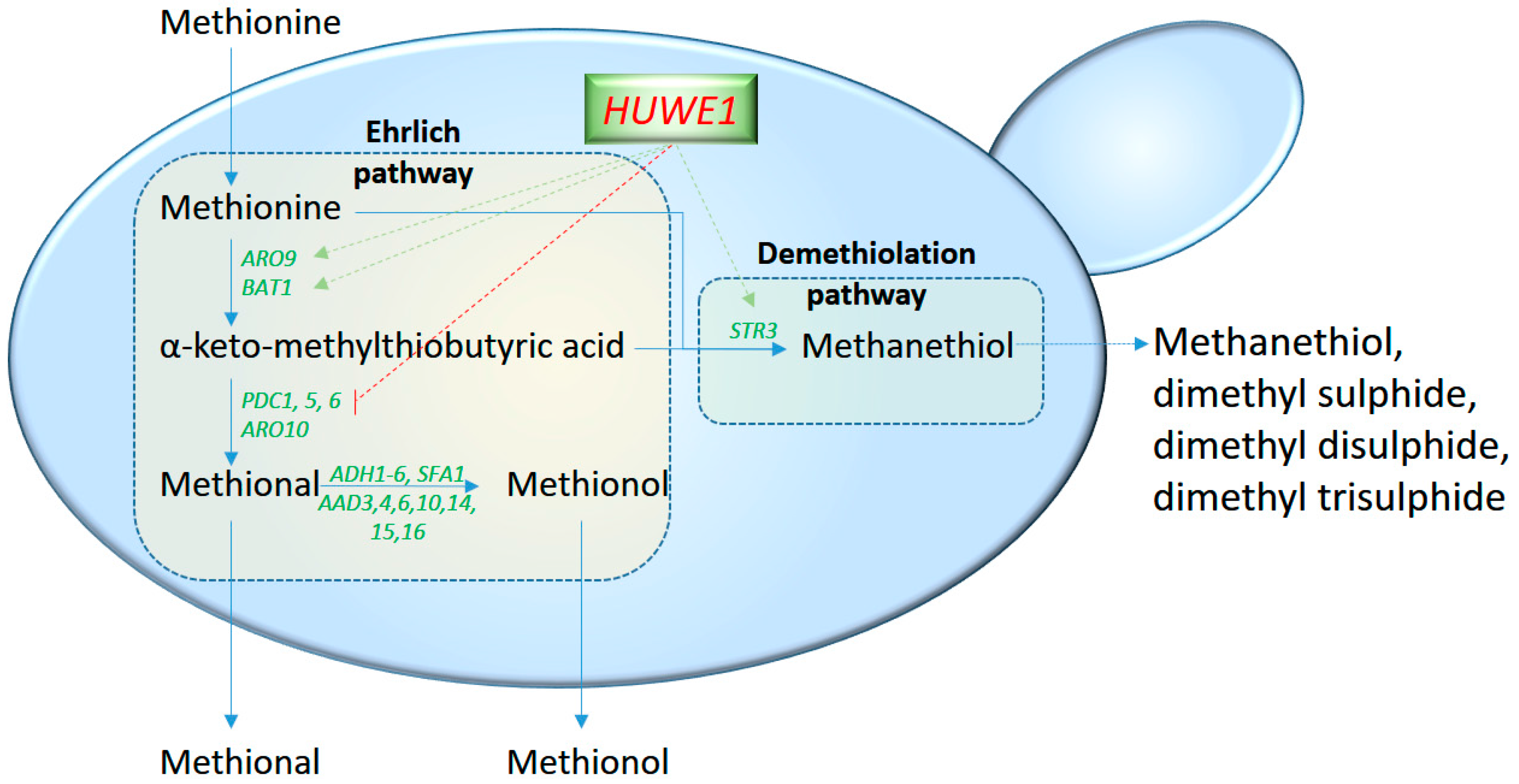
2.1. Higher Alcohols and the Ehrlich Pathway Regulation
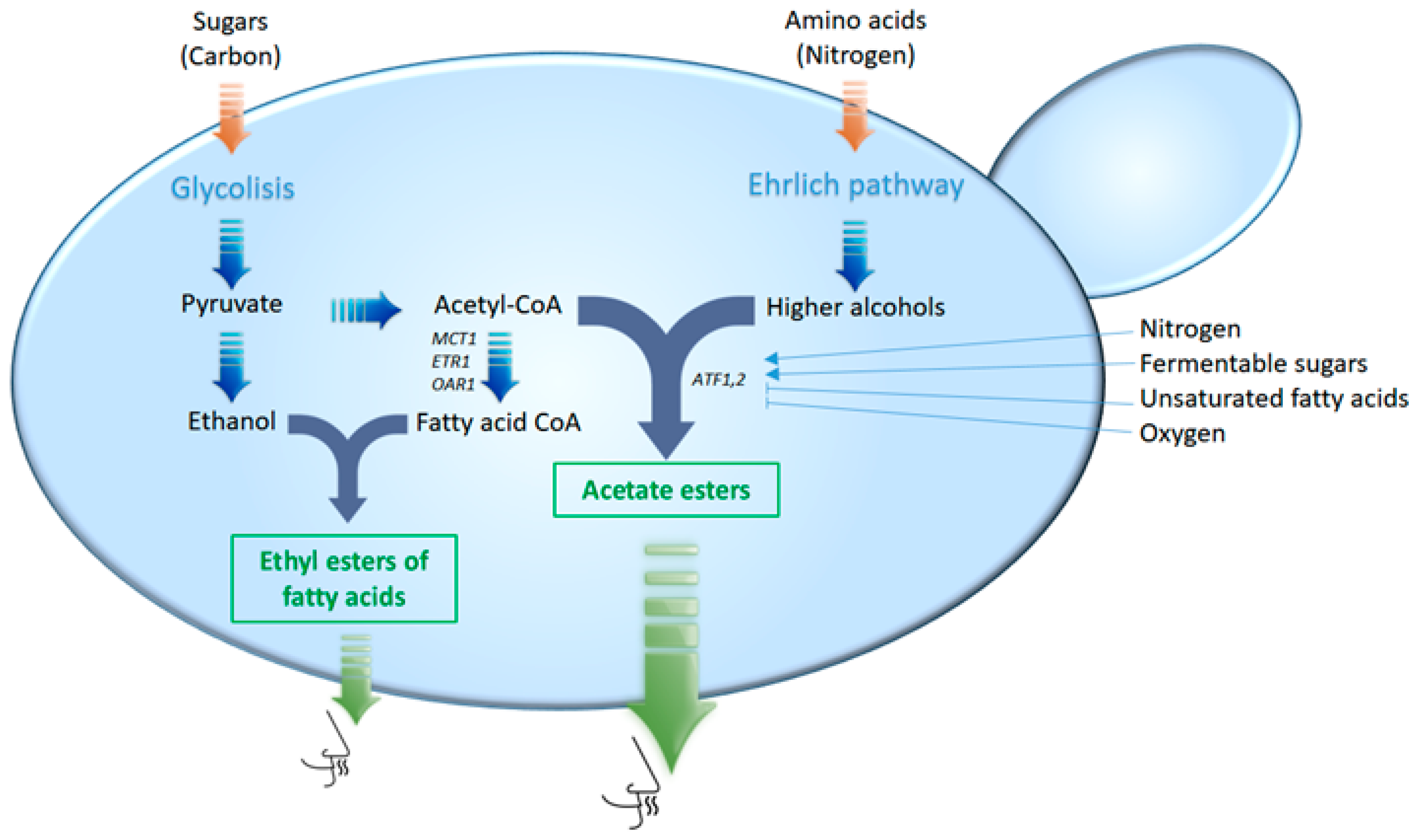
2.2. Esters
3. Wine Microorganisms and Varietal Aroma Compounds
3.1. Production of Volatile Sulphur Compounds
3.1.1. Undesirable Sulphur Compounds
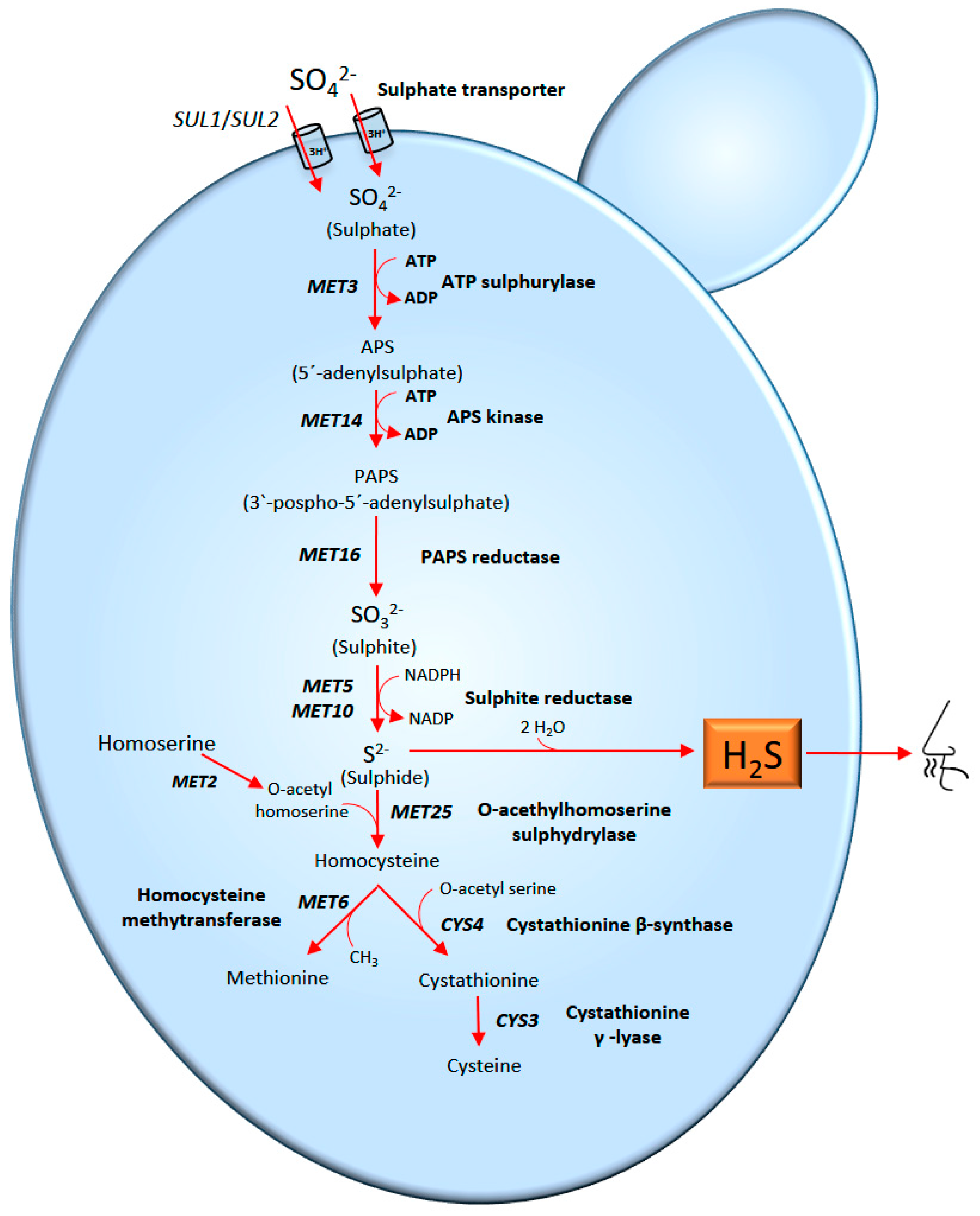
Hydrogen Sulphide
Dimethyl Sulphide and Methanethiol
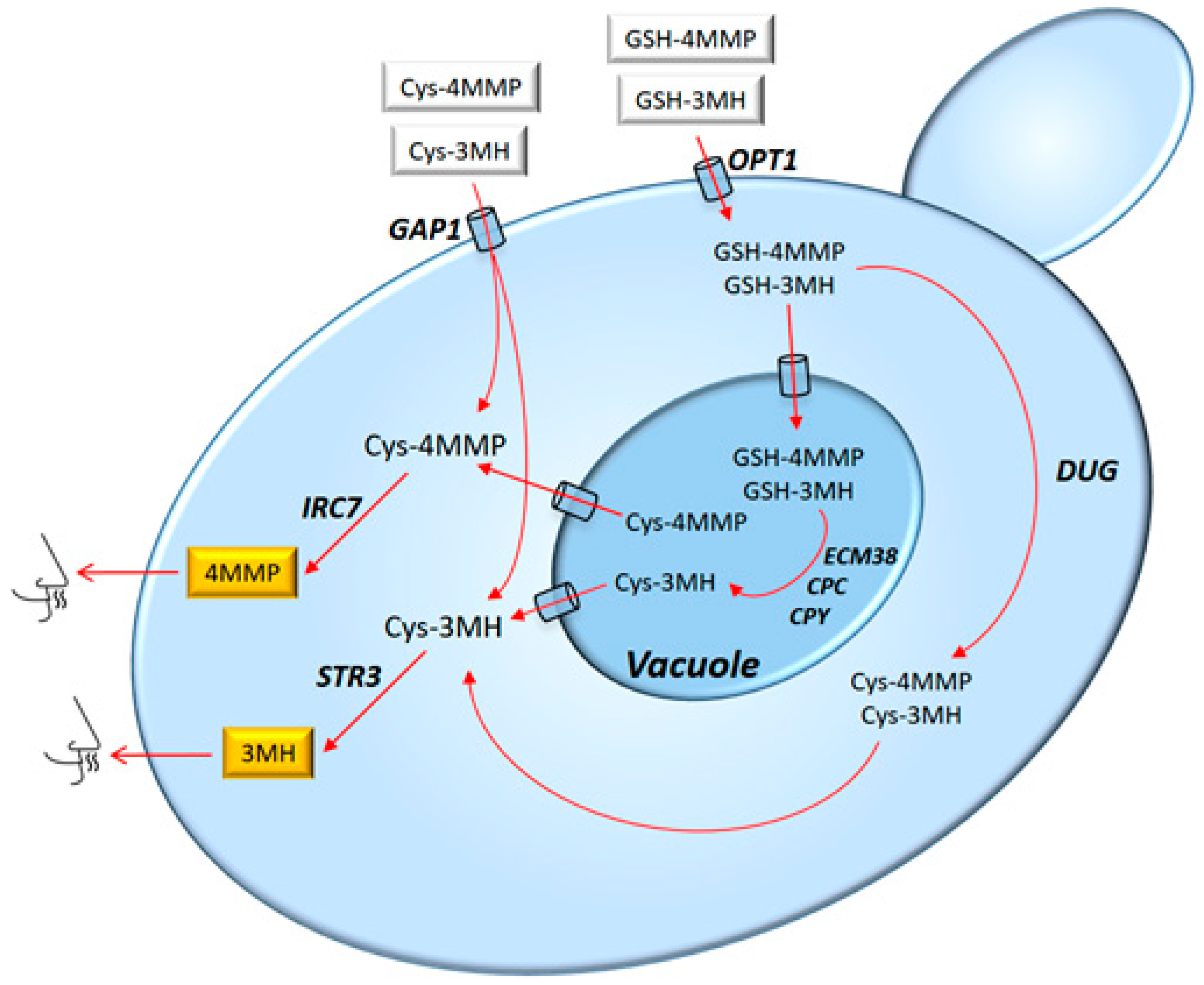
3.1.2. Desirable Sulphur Compounds
3.1.3. Metabolic and Gene Regulation
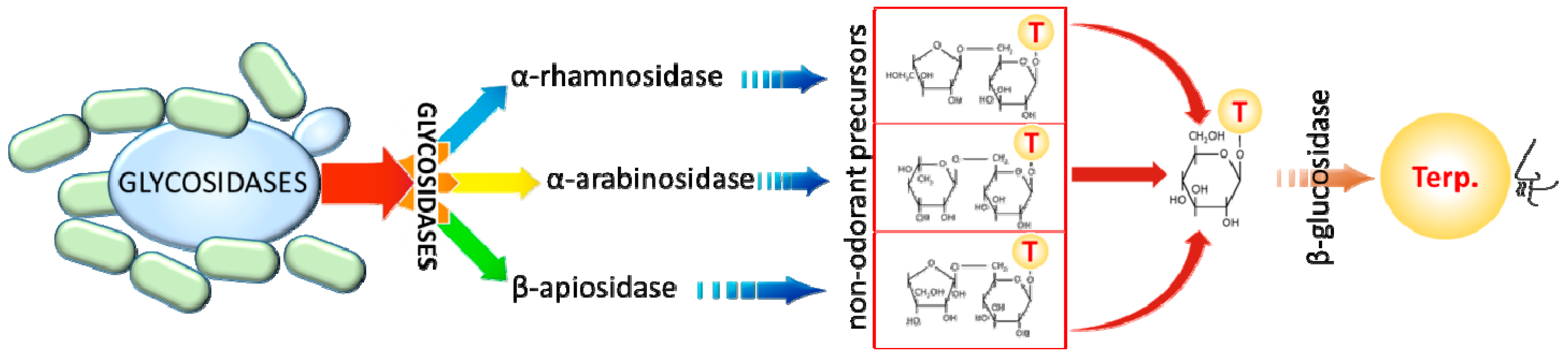
3.2. Wine Yeast and Bacteria Production of Monoterpenes
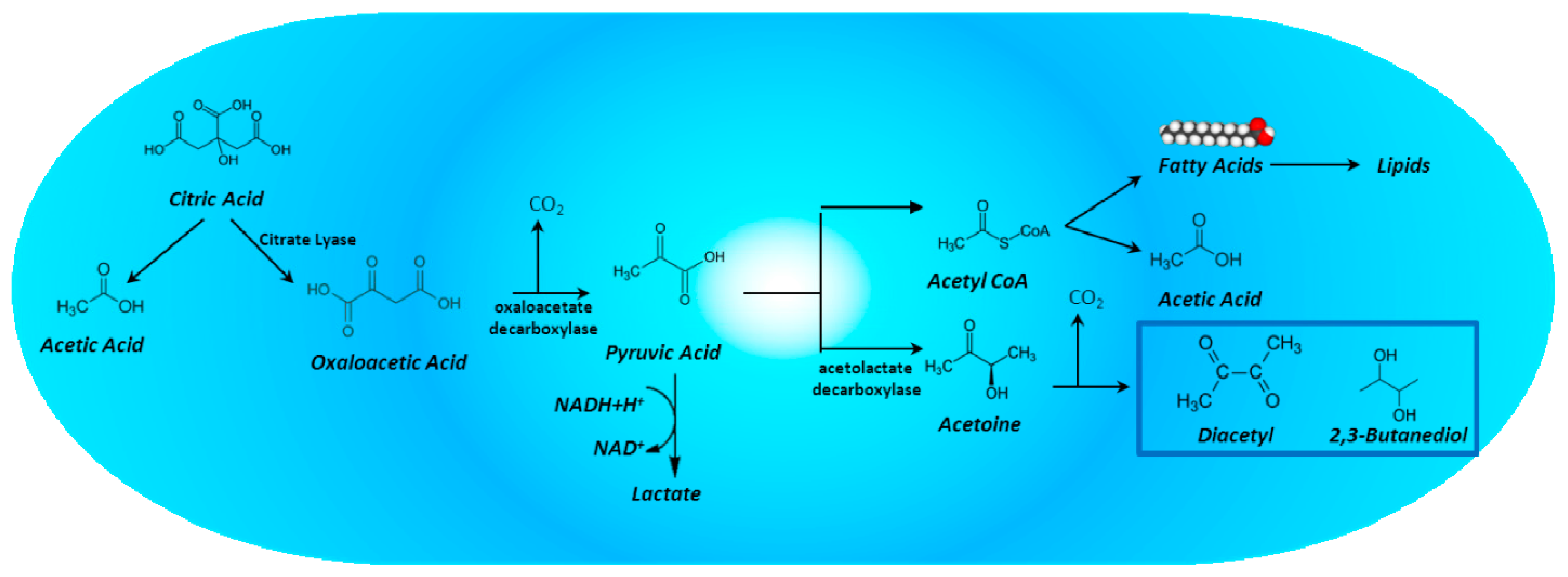
4. Citric Acid Degradation by Lactic Acid Bacteria and Its Impact on the Aromatic Quality of Wine
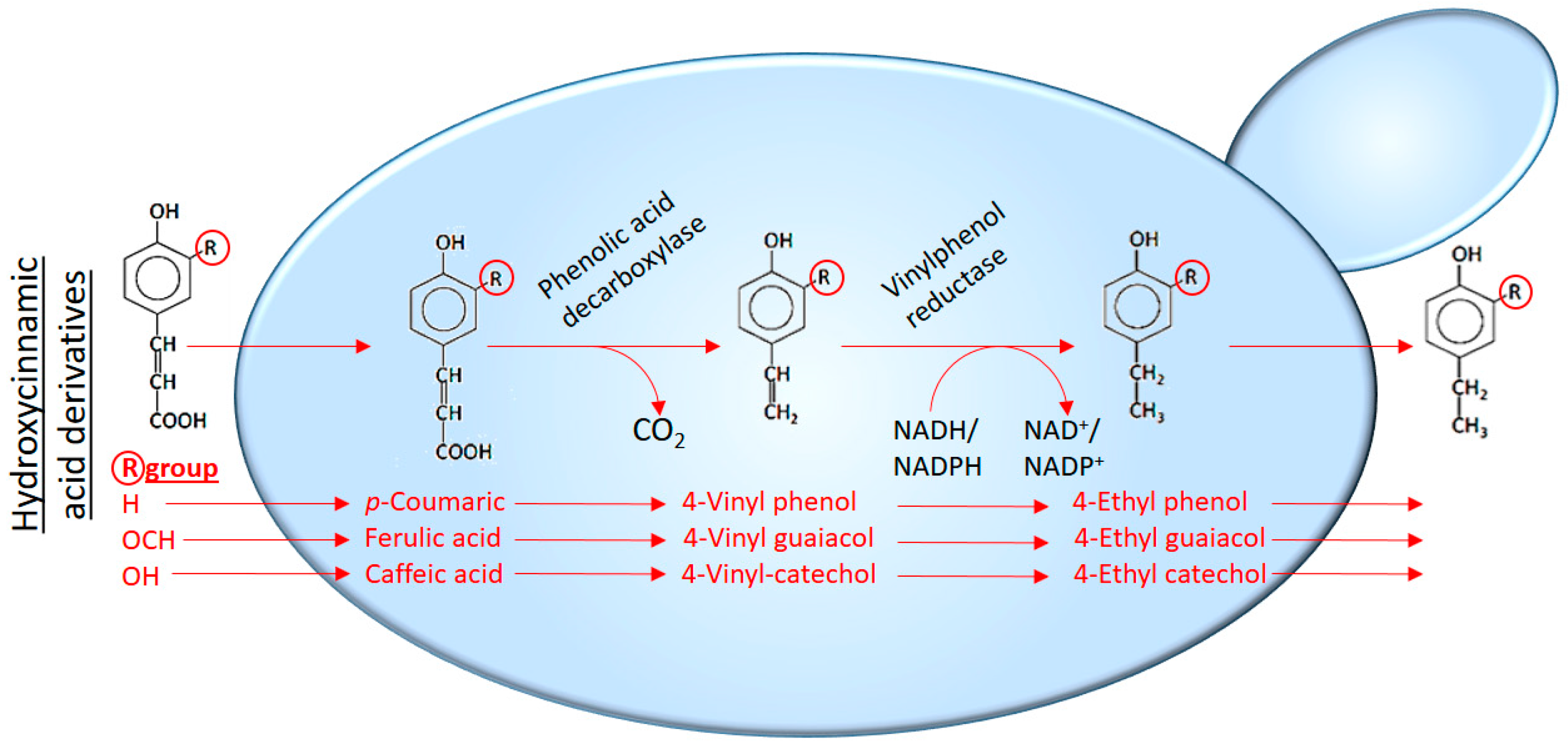
5. Brettanomyces/Dekkera bruxellensis and Volatile Phenols
6. Production of Off-Flavours by Lactic Acid Bacteria
7. Contribution of Oral Microbiota to Wine Aroma Perception
8. Future Perspectives and Conclusions
Acknowledgments
Conflicts of Interest
References
- Polaskova, P.; Herszage, J.; Ebeler, S.E. Wine flavor: Chemistry in a glass. Chem. Soc. Rev. 2008, 37, 2478–2489. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rousseaux, S.; Tourdot-Marechal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome, a dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2015, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial biogeography of wine grapes is conditioned by cultivar, vintage, and climate. Proc. Natl. Acad. Sci. USA 2014, 111, E139–E148. [Google Scholar] [CrossRef] [PubMed]
- Zarraonaindia, I.; Owens, S.M.; Weisenhorn, P.; West, K.; Hampton-Marcell, J.; Lax, S.; Bokulich, N.A.; Mills, D.A.; Martin, G.; Taghavi, S. The soil microbiome influences grapevine-associated microbiota. mBio 2015, 6, e02527-14. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.A.; van der Lelie, D.; Zarraonaindia, I. Microbial terroir for wine grapes. Proc. Natl. Acad. Sci. USA 2014, 111, 5–6. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the enzymatic basis of wine “flavorome”: A phylo-functional study of wine related yeast species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Pérez, J.M.; Campo, E.; San-Juan, F.; Coque, J.J.R.; Ferreira, V.; Hernández-Orte, P. Sensory and chemical characterisation of the aroma of prieto picudo rosé wines: The differential role of autochthonous yeast strains on aroma profiles. Food Chem. 2012, 133, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Sills, H.; Capozzi, V.; Romano, A.; Cappellin, L.; Spano, G.; Breniaux, M.; Lucas, P.; Biasioli, F. Advances in wine analysis by PTR-ToF-MS: optimization of the method and discrimination of wines fermented with different malolactic starters. Int. J. Mass. Spectrom. 2016, 397–398, 42–51. [Google Scholar] [CrossRef]
- Bartowsky, E.J. Oenococcus oeni and malolactic fermentation–moving into the molecular arena. Aust. J. Grape Wine Res. 2005, 11, 174–187. [Google Scholar] [CrossRef]
- du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food Bioprocess. Technol. 2011, 4, 876–906. [Google Scholar] [CrossRef]
- Berbegal, C.; Peña, N.; Russo, P.; Grieco, F.; Pardo, I.; Ferrer, S.; Spano, G.; Capozzi, V. Technological properties of Lactobacillus plantarum strains isolated from grape must fermentation. Food Microbiol. 2016, 57, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, M.V.; Polo, M.C. Winemaking biochemistry and microbiology: Current knowledge and future trends. Crit. Rev. Food Sci. Nutr. 2005, 45, 265–286. [Google Scholar] [CrossRef] [PubMed]
- Munoz, R.; Moreno-Arribas, M.; de las Rivas, B. Lactic acid bacteria. In Molecular Wine Microbiology, 1st ed.; Carrascosa, A.V., Muñoz, R., González, R., Eds.; Elsevier Academic Press: London, UK, 2011; pp. 191–226. [Google Scholar]
- Lonvaud-Funel, A. Effects of malolactic fermentation in wine quality. In Managing Wine Quality: Oenology and Wine Quality; Reynolds, A., Ed.; Woodhead Publishing Limited: Cambridge, UK, 2010. [Google Scholar]
- Tao, Y.; García, J.F.; Sun, D.-W. Advances in wine aging technologies for enhancing wine quality and accelerating wine aging process. Crit. Rev. Food Sci. Nutr. 2014, 54, 817–835. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Arribas, V.; Pueyo, E.; Polo, M.C. Peptides in musts and wines. Changes during the manufacture of cavas (sparkling wines). J. Agric. Food. Chem. 1996, 44, 3783–3788. [Google Scholar] [CrossRef]
- Alcaide-Hidalgo, J.M.; Moreno-Arribas, M.V.; Martín-Álvarez, P.J.; Polo, M.C. Influence of malolactic fermentation, postfermentative treatments and ageing with lees on nitrogen compounds of red wines. Food Chem. 2007, 103, 572–581. [Google Scholar] [CrossRef]
- Zeng, X.A.; Yu, S.J.; Zhang, L.; Chen, X.D. The effects of ac electric field on wine maturation. Innov. Food Sci. Emerg. 2008, 9, 463–468. [Google Scholar] [CrossRef]
- Loira, I.; Vejarano, R.; Morata, A.; Ricardo-da-Silva, J.; Laureano, O.; González, M.; Suárez-Lepe, J. Effect of Saccharomyces strains on the quality of red wines aged on lees. Food Chem. 2013, 139, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Juega, M.; Carrascosa, A.V.; Martinez-Rodriguez, A.J. Effect of short ageing on lees on the mannoprotein content, aromatic profile, and sensorial character of white wines. J. Food. Sci. 2015, 80, M384–M388. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderón, F.; Benito, S. Outlining the influence of non-conventional yeasts in wine ageing over lees. Yeast 2016, 33, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Suárez, R.; Suárez-Lepe, J.; Morata, A.; Calderón, F. The production of ethylphenols in wine by yeasts of the genera Brettanomyces and Dekkera: A review. Food Chem. 2007, 102, 10–21. [Google Scholar] [CrossRef]
- Carrau, F.M.; Medina, K.; Boido, E.; Farina, L.; Gaggero, C.; Dellacassa, E.; Versini, G.; Henschke, P.A. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 2005, 243, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Gamero, A.; Belloch, C.; Querol, A. Genomic and transcriptomic analysis of aroma synthesis in two hybrids between Saccharomyces cerevisiae and S. kudriavzevii in winemaking conditions. Microb. Cell. Fact. 2015, 14, 128. [Google Scholar] [CrossRef] [PubMed]
- Janssens, L.; De Pooter, H.; Schamp, N.; Vandamme, E. Production of flavours by microorganisms. Process. Biochem 1992, 27, 195–215. [Google Scholar] [CrossRef]
- Krings, U.; Berger, R. Biotechnological production of flavours and fragrances. Appl. Microbiol. Biotechnol. 1998, 49, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stribny, J.; Gamero, A.; Pérez-Torrado, R.; Querol, A. Saccharomyces kudriavzevii and Saccharomyces uvarum differ from Saccharomyces cerevisiae during the production of aroma-active higher alcohols and acetate esters using their amino acidic precursors. Int. J. Food. Microbiol. 2015, 205, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- The Yeasts: A Taxonomic Study; Kurtzman, C.P.; Fell, J.W.; Boekhout, T. (Eds.) Elsevier: Amsterdam, The Netherlands, 2011.
- Esteve-Zarzoso, B.; Manzanares, P.; Ramon, D.; Querol, A. The role of non-Saccharomyces yeasts in industrial winemaking. Int. Microbiol. 1998, 1, 143–148. [Google Scholar] [PubMed]
- García, V.; Vásquez, H.; Fonseca, F.; Manzanares, P.; Viana, F.; Martínez, C.; Ganga, M. Effects of using mixed wine yeast cultures in the production of chardonnay wines. Rev. Argent. Microbiol. 2010, 42, 226–229. [Google Scholar] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food. Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Gamero, A.; Quintilla, R.; Groenewald, M.; Alkema, W.; Boekhout, T.; Hazelwood, L. High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiol. 2016, 60, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, C.; Mendes-Faia, A.; Lage, P.; Mira, N.P.; Mendes-Ferreira, A. Genomic expression program of Saccharomyces cerevisiae along a mixed-culture wine fermentation with Hanseniaspora guilliermondii. Microb. Cell. Fact. 2015, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Ebeler, S.E. Analytical chemistry: Unlocking the secrets of wine flavor. Food. Rev. Int. 2001, 17, 45–64. [Google Scholar] [CrossRef]
- Rapp, A.; Versini, G. Influence of nitrogen compounds in grapes on aroma compounds of wines. Dev. Food Sci. 1995, 37, 1659–1694. [Google Scholar]
- Tao, Y.; Li, H.; Wang, H.; Zhang, L. Volatile compounds of young cabernet sauvignon red wine from Changli county (China). J. Food Compos. Anal. 2008, 21, 689–694. [Google Scholar] [CrossRef]
- Rankine, B. Formation of higher alcohols by wine yeasts, and relationship to taste thresholds. J. Sci. Food Agric. 1967, 18, 583–589. [Google Scholar] [CrossRef]
- Giudici, P.; Romano, P.; Zambonelli, C. A biometric study of higher alcohol production in Saccharomyces cerevisiae. Can. J. Microbiol. 1990, 36, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Schulthess, D.; Ettlinger, L. Influence of the concentration of branched chain amino acids on the formation of fusel alcohols. J. Inst. Brew. 1978, 84, 240–243. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2015, 31, 277–293. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Navascues, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, F. Über die bedingungen der fuselölbildung und über ihren zusammenhang mit dem eiweissaufbau der hefe. Ber. Dtsch. Chem. Ges. 1907, 40, 1027–1047. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.-M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef]
- Cooper, T.G. Transmitting the signal of excess nitrogen in Saccharomyces cerevisiae from the tor proteins to the gata factors: Connecting the dots. FEMS Microbiol. Rev. 2002, 26, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef]
- Lilly, M.; Bauer, F.F.; Styger, G.; Lambrechts, M.G.; Pretorius, I.S. The effect of increased branched-chain amino acid transaminase activity in yeast on the production of higher alcohols and on the flavour profiles of wine and distillates. FEMS Yeast. Res. 2006, 6, 726–743. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jia, K.-Z.; Xia, S.-T.; Xu, Y.-H.; Liu, R.-S.; Li, H.-M.; Tang, Y.-J. Regulating Ehrlich and demethiolation pathways for alcohols production by the expression of ubiquitin-protein ligase gene HUWE1. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Sáenz-Navajas, M.-P.; Ballester, J.; Fernández-Zurbano, P.; Ferreira, V.; Peyron, D.; Valentin, D. Wine quality perception: A sensory point of view. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé-Suáldea, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 119–138. [Google Scholar]
- Marais, J.; Van Rooyen, P.; Plessis, C. Objective quality rating of pinotage wine. Vitis 1979, 18, 31–39. [Google Scholar]
- Lee, S.-J.; Rathbone, D.; Asimont, S.; Adden, R.; Ebeler, S.E. Dynamic changes in ester formation during chardonnay juice fermentations with different yeast inoculation and initial brix conditions. Am. J. Enol Vitic. 2004, 55, 346–354. [Google Scholar]
- Garofolo, A.; Piracci, A. Evolution des esters des acides gras pendant la conservation des vins: Constantes d'équilibre et énergies d’activation. Bulletin de l'OIV 1994, 67, 225–245. [Google Scholar]
- Stribny, J.; Querol, A.; Pérez-Torrado, R. Differences in enzymatic properties of the Saccharomyces kudriavzevii and Saccharomyces uvarum alcohol acetyltransferases and their impact on aroma-active compounds production. Front. Microbiol. 2016, 7, 897. [Google Scholar] [CrossRef] [PubMed]
- Malcorps, P.; Dufour, J.P. Short-chain and medium-chain aliphatic-ester synthesis in Saccharomyces cerevisiae. Eur. J. Biochem. 1992, 210, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Beltran, G.; Novo, M.; Leberre, V.; Sokol, S.; Labourdette, D.; Guillamon, J.-M.; Mas, A.; François, J.; Rozes, N. Integration of transcriptomic and metabolic analyses for understanding the global responses of low-temperature winemaking fermentations. FEMS Yeast. Res. 2006, 6, 1167–1183. [Google Scholar] [CrossRef] [PubMed]
- Nagasawa, N.; Bogaki, T.; Iwamatsu, A.; Hamachi, M.; Kumagai, C. Cloning and Nucleotide Sequence of the Alcohol Acetyltransferase II Gene (ATF2) from Saccharomyces cerevisiae Kyokai No. 7. Biosci. Biotechnol. Biochem. 1998, 62, 1852–1857. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, M.; Pretorius, I. Yeast and its importance to wine aroma. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar]
- Sadoudi, M.; Tourdot-Marechal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacon, J.J.; Ballester, J.; Vichi, S.; Guerin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.; Mendes, F.; Hogg, T.; Vasconcelos, I. Alcohols, esters and heavy sulphur compounds production by pure and mixed cultures of apiculate wine yeasts. Int. J. Food Microbiol. 2005, 103, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Lleixà, J.; Martín, V.; Portillo, M.d.C.; Carrau, F.; Beltran, G.; Mas, A. Comparison of fermentation and wines produced by inoculation of Hanseniaspora vineae and Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 338. [Google Scholar] [CrossRef] [PubMed]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food. Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [PubMed]
- Iranzo, J.U.; Perez, A.B.; Canas, P.I. Study of the oenological characteristicsand enzymatic activities of wine yeasts. Food Microbiol. 1998, 15, 399–406. [Google Scholar] [CrossRef]
- Pozo-Bayón, M.; G-Alegría, E.; Polo, M.; Tenorio, C.; Martín-Álvarez, P.; Calvo De La Banda, M.; Ruiz-Larrea, F.; Moreno-Arribas, M. Wine volatile and amino acid composition after malolactic fermentation: Effect of Oenococcus oeni and Lactobacillus plantarum starter cultures. J. Agric. Food. Chem. 2005, 53, 8729–8735. [Google Scholar] [CrossRef] [PubMed]
- Antalick, G.; Perello, M.-C.; de Revel, G. Characterization of fruity aroma modifications in red wines during malolactic fermentation. J. Agric. Food. Chem. 2012, 60, 12371–12383. [Google Scholar] [CrossRef] [PubMed]
- Sumby, K.; Grbin, P.; Jiranek, V. Characterization of EstCOo8 and EstC34, intracellular esterases, from the wine-associated lactic acid bacteria Oenococcus oeni and Lactobacillus hilgardii. J. Appl. Microbiol. 2013, 114, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martín, F.; Seseña, S.; Izquierdo, P.M.; Palop, M.L. Esterase activity of lactic acid bacteria isolated from malolactic fermentation of red wines. Int. J. Food Microbiol. 2013, 163, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Costello, P.; Siebert, T.; Solomon, M.; Bartowsky, E. Synthesis of fruity ethyl esters by acyl coenzyme a: Alcohol acyltransferase and reverse esterase activities in Oenococcus oeni and Lactobacillus plantarum. J. Appl. Microbiol. 2013, 114, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Torres, M.; Reverón, I.; Mancheño, J.M.; de las Rivas, B.; Muñoz, R. Characterization of a feruloyl esterase from Lactobacillus plantarum. Appl. Environ. Microbiol. 2013, 79, 5130–5136. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Torres, M.; Barcenilla, J.M.; Mancheño, J.M.; de las Rivas, B.; Muñoz, R. Characterization of a versatile arylesterase from Lactobacillus plantarum active on wine esters. J. Agric. Food Chem. 2014, 62, 5118–5125. [Google Scholar] [CrossRef] [PubMed]
- Mestres, M.; Busto, O.; Guasch, J. Analysis of organic sulfur compounds in wine aroma. J. Chromatogr A 2000, 881, 569–581. [Google Scholar] [CrossRef]
- Tominaga, T.; Darriet, P.; Dubourdieu, D. Identification of 3-mercaptohexyl acetate in sauvignon wine, a powerful aromatic compound exhibiting box-tree odor. Vitis 1996, 35, 207–210. [Google Scholar]
- Pripis-Nicolau, L.; Revel, G.d.; Bertrand, A.; Lonvaud-Funel, A. Methionine catabolism and production of volatile sulphur compounds by oenococcus oeni. J. Appl. Microbiol. 2004, 96, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Moreira, N.; Mendes, F.; Pereira, O.; de Pinho, P.G.; Hogg, T.; Vasconcelos, I. Volatile sulphur compounds in wines related to yeast metabolism and nitrogen composition of grape musts. Anal. Chim. Acta 2002, 458, 157–167. [Google Scholar] [CrossRef]
- Smith, M.; Bekker, M.; Smith, P.; Wilkes, E. Sources of volatile sulfur compounds in wine. Aust. J. Grape Wine Res. 2015, 21, 705–712. [Google Scholar] [CrossRef]
- Rauhut, D. Yeasts-production of sulfur compounds. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Reading, UK, 1993; Volume 6, pp. 183–223. [Google Scholar]
- Amerine, M.A.; Berg, H.V.; Kunkee, R.E.; Ough, C.S.; Singleton, V.L.; Webb, A.D. The Technology of Wine Making, 4th ed.; AVI Publishing Company: Westport, CT, USA, 1980. [Google Scholar]
- Spiropoulos, A.; Bisson, L.F. Met17 and hydrogen sulfide formation in Saccharomyces cerevisiae. Appl Environ. Microbiol. 2000, 66, 4421–4426. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Curtin, C.D.; Varela, C.; Pretorius, I.S. Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 2012, 96, 601–618. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.; Pretorius, I. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Mendes-Ferreira, A.; Mendes-Faia, A.; Leao, C. Survey of hydrogen sulphide production by wine yeasts. J. Food Protect. 2002, 65, 1033–1037. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.; Loza-Tavera, H.; Hernández-Navarro, A.; Moreno-Sánchez, R. Sulfur assimilation and glutathione metabolism under cadmium stress in yeast, protists and plants. FEMS Microbiol. Rev. 2005, 29, 653–671. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Surdin-Kerjan, Y. Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Res. 1997, 61, 503–532. [Google Scholar]
- Wysocki, R.; Tamás, M.J. How Saccharomyces cerevisiae copes with toxic metals and metalloids. FEMS Microbiol. Rev. 2010, 34, 925–951. [Google Scholar] [CrossRef] [PubMed]
- Blaiseau, P.L.; Thomas, D. Multiple transcriptional activation complexes tether the yeast activator met4 to DNA. EMBO J. 1998, 17, 6327–6336. [Google Scholar] [CrossRef] [PubMed]
- Natarajan, K.; Meyer, M.R.; Jackson, B.M.; Slade, D.; Roberts, C.; Hinnebusch, A.G.; Marton, M.J. Transcriptional profiling shows that Gcn4p is a master regulator of gene expression during amino acid starvation in yeast. Mol. Cell. Biol. 2001, 21, 4347–4368. [Google Scholar] [CrossRef] [PubMed]
- Linderholm, A.L.; Olineka, T.L.; Hong, Y.; Bisson, L.F. Allele diversity among genes of the sulfate reduction pathway in wine strains of Saccharomyces cerevisiae. Am. J. Enol. Vitic. 2006, 57, 431–440. [Google Scholar]
- Segurel, M.A.; Razungles, A.J.; Riou, C.; Salles, M.; Baumes, R.L. Contribution of dimethyl sulfide to the aroma of syrah and grenache noir wines and estimation of its potential in grapes of these varieties. J. Agric. Food. Chem. 2004, 52, 7084–7093. [Google Scholar] [CrossRef] [PubMed]
- Kinzurik, M.I.; Herbst-Johnstone, M.; Gardner, R.C.; Fedrizzi, B. Evolution of volatile sulfur compounds during wine fermentation. J. Agric. Food. Chem. 2015, 63, 8017–8024. [Google Scholar] [CrossRef] [PubMed]
- Perpète, P.; Duthoit, O.; De Maeyer, S.; Imray, L.; Lawton, A.I.; Stavropoulos, K.E.; Gitonga, V.W.; Hewlins, M.J.; Dickinson, J.R. Methionine catabolism in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Zamora, F. El potencial de oxidorreducción en enología. ACE: Revista de enología 2010, 2, 114. [Google Scholar]
- Vallet, A.; Santarelli, X.; Lonvaud-Funel, A.; De Revel, G.; Cabanne, C. Purification of an alcohol dehydrogenase involved in the conversion of methional to methionol in Oenococcus oeni IOEB 8406. Appl. Microbiol. Biotechnol. 2009, 82, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Darriet, P.; Tominaga, T.; Lavigne, V.; Boidron, J.N.; Dubourdieu, D. Identification of a powerful aromatic component of Vitis vinifera L. Var. Sauvignon wines: 4-mercapto-4-methylpentan-2-one. Flavour Frag. J. 1995, 10, 385–392. [Google Scholar] [CrossRef]
- Dubourdieu, D.; Tominaga, T.; Masneuf, I.; des Gachons, C.P.; Murat, M.L. The role of yeasts in grape flavor development during fermentation: The example of sauvignon blanc. Am. J. Enol. Vitic. 2006, 57, 81–88. [Google Scholar]
- Tominaga, T.; Masneuf, I.; Dubourdieu, D. Mise en evidence d'un S-conjugue de la cysteine, precurseur d'arome du sauvignon. J. Int. Sci. Vigne Vin. 1995, 29, 227–232. [Google Scholar]
- Tominaga, T.; Peyrot des Gachons, C.; Dubourdieu, D. A new type of flavor precursors in Vitis vinifera l. Cv. Sauvignon blanc: S-cysteine conjugates. J. Agric. Food Chem. 1998, 46, 5215–5219. [Google Scholar] [CrossRef]
- Fedrizzi, B.; Pardon, K.H.; Sefton, M.A.; Elsey, G.M.; Jeffery, D.W. First identification of 4-S-glutathionyl-4-methylpentan-2-one, a potential precursor of 4-mercapto-4-methylpentan-2-one, in sauvignon blanc juice. J. Agric. Food Chem. 2009, 57, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.S.; Swiegers, J.H.; Elsey, G.M.; Siebert, T.E.; Bartowsky, E.J.; Fleet, G.H.; Pretorius, I.S.; de Barros Lopes, M.A. Variation in 4-mercapto-4-methyl-pentan-2-one release by Saccharomyces cerevisiae commercial wine strains. FEMS Microbiol. Lett. 2004, 240, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Roland, A.; Schneider, R.; Razungles, A.; Cavelier, F. Varietal thiols in wine: Discovery, analysis and applications. Chem. Rev. 2011, 111, 7355–7376. [Google Scholar] [CrossRef] [PubMed]
- Maggu, M.; Winz, R.; Kilmartin, P.A.; Trought, M.C.; Nicolau, L. Effect of skin contact and pressure on the composition of sauvignon blanc must. J. Agric. Food Chem. 2007, 55, 10281–10288. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Gardner, R.C. Yeast genes required for conversion of grape precursors to varietal thiols in wine. FEMS Yeast Res. 2015, 15, fov034. [Google Scholar] [CrossRef] [PubMed]
- Subileau, M.; Schneider, R.; Salmon, J.-M.; Degryse, E. Nitrogen catabolite repression modulates the production of aromatic thiols characteristic of sauvignon blanc at the level of precursor transport. FEMS Yeast Res. 2008, 8, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Cordente, A.G.; Capone, D.L.; Curtin, C.D. Unravelling glutathione conjugate catabolism in Saccharomyces cerevisiae: The role of glutathione/dipeptide transporters and vacuolar function in the release of volatile sulfur compounds 3-mercaptohexan-1-ol and 4-mercapto-4-methylpentan-2-one. Appl. Microbiol. Biotechnol. 2015, 99, 9709–9722. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.S.; Klein, M.; Swiegers, J.H.; Hayasaka, Y.; Elsey, G.M.; Fleet, G.H.; Hoj, P.B.; Pretorius, I.S.; de Barros Lopes, M.A. Genetic determinants of volatile-thiol release by Saccharomyces cerevisiae during wine fermentation. Appl. Environ. Microbiol. 2005, 71, 5420–5426. [Google Scholar] [CrossRef] [PubMed]
- Roncoroni, M.; Santiago, M.; Hooks, D.O.; Moroney, S.; Harsch, M.J.; Lee, S.A.; Richards, K.D.; Nicolau, L.; Gardner, R.C. The yeast IRC7 gene encodes a β-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 2011, 28, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Belda, I.; Ruiz, J.; Navascués, E.; Marquina, D.; Santos, A. Improvement of aromatic thiol release through the selection of yeasts with increased β-lyase activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Holt, S.; Cordente, A.G.; Curtin, C. Saccharomyces cerevisiae STR3 and yeast cystathionine beta-lyase enzymes: The potential for engineering increased flavor release. Bioeng. Bugs 2012, 3, 178–180. [Google Scholar] [PubMed]
- Thibon, C.; Marullo, P.; Claisse, O.; Cullin, C.; Dubourdieu, D.; Tominaga, T. Nitrogen catabolic repression controls the release of volatile thiols by Saccharomyces cerevisiae during wine fermentation. FEMS Yeast Res. 2008, 8, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Murat, M.-L.; Tominaga, T.; Dubourdieu, D. Assessing the aromatic potential of Cabernet Sauvignon and Merlot musts used to produce rose wine by assaying the cysteinylated precursor of 3-mercaptohexan-1-ol. J. Agric. Food Chem. 2001, 49, 5412–5417. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, C.; du Toit, W.J. A comprehensive review on sauvignon blanc aroma with a focus on certain positive volatile thiols. Food Res. Int. 2012, 45, 287–298. [Google Scholar] [CrossRef]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in sauvignon blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; Moine, V.; Thibon, C.; Bely, M. Enhanced 3-sulfanylhexan-1-ol production in sequential mixed fermentation with Torulaspora delbrueckii/ Saccharomyces cerevisiae reveals a situation of synergistic interaction between two industrial strains. Front. Microbiol. 2016, 7, 293. [Google Scholar] [CrossRef] [PubMed]
- Baumes, R. Wine aroma precursors. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, C., Eds.; Springer: Cham, Switzerland, 2009; pp. 251–274. [Google Scholar]
- Maicas, S.; Mateo, J.J. Hydrolysis of terpenyl glycosides in grape juice and other fruit juices: A review. Appl. Microbiol. Biotechnol. 2005, 67, 322–335. [Google Scholar] [CrossRef] [PubMed]
- Marais, J. Terpenes in the aroma of grapes and wines: A review. S. Afr. J. Enol. Vitic. 1983, 4, 49–60. [Google Scholar]
- Swiegers, J.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Mateo, J.; Jiménez, M. Monoterpenes in grape juice and wines. J. Chromatogr. A 2000, 881, 557–567. [Google Scholar] [CrossRef]
- Park, S.K.; Morrison, J.C.; Adams, D.O.; Noble, A.C. Distribution of free and glycosidically bound monoterpenes in the skin and mesocarp of muscat of alexandria grapes during development. J. Agric. Food Chem. 1991, 39, 514–518. [Google Scholar] [CrossRef]
- Gunata, Y.; Bayonove, C.; Baumes, R.; Cordonnier, R. The aroma of grapes. Extraction and determination of free and glycosidically bound fractions of some grape aroma components. J. Chromatogr. A 1985, 331, 83–90. [Google Scholar] [CrossRef]
- Voirin, S.G.; Baumes, R.L.; Bitteur, S.M.; Gunata, Z.Y.; Bayonove, C.L. Novel monoterpene disaccharide glycosides of Vitis vinifera grapes. J. Agric. Food Chem. 1990, 38, 1373–1378. [Google Scholar] [CrossRef]
- Mateo, J.J.; di Stefano, R. Description of the β-glucosidase activity of wine yeasts. Food Microbiol. 1997, 14, 583–591. [Google Scholar] [CrossRef]
- Van Rensburg, P.; Pretorius, I. Enzymes in winemaking: Harnessing natural catalysts for efficient biotransformations-a review. S. Afr. J. Enol. Vitic. 2000, 21, 52–73. [Google Scholar]
- Gunata, Z.; Bitteur, S.; Brillouet, J.-M.; Bayonove, C.; Cordonnier, R. Sequential enzymic hydrolysis of potentially aromatic glycosides from grape. Carbohydr. Res. 1988, 184, 139–149. [Google Scholar] [CrossRef]
- Delcroix, A.; Günata, Z.; Sapis, J.-C.; Salmon, J.M.; Bayonove, C. Glycosidase activities of three enological yeast strains during winemaking: Effect on the terpenol content of muscat wine. Am. J. Enol. Vitic. 1994, 45, 291–296. [Google Scholar]
- Quatrini, P.; Marineo, S.; Puglia, A.M.; Restuccia, C.; Caggia, C.; Randazzo, C.L.; Spagna, G.; Barbagallo, R.N.; Palmeri, R.; Giudici, P. Partial sequencing of the β-glucosidase-encoding gene of yeast strains isolated from musts and wines. Ann. Microbiol. 2008, 58, 503–508. [Google Scholar] [CrossRef]
- Lopez, M.C.; Mateo, J.J.; Maicas, S. Screening of beta-glucosidase and beta-xylosidase activities in four non-Saccharomyces yeast isolates. J. Food Sci. 2015, 80, C1696–C1704. [Google Scholar] [CrossRef] [PubMed]
- Rosi, I.; Vinella, M.; Domizio, P. Characterization of β-glucosidase activity in yeasts of oenological origin. J. Appl. Bacteriol. 1994, 77, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, L.; Espinosa, J.; Fernandez-Gonzalez, M.; Briones, A. β-glucosidase activity in a Saccharomyces cerevisiae wine strain. Int. J. Food Microbiol. 2003, 80, 171–176. [Google Scholar] [CrossRef]
- Riou, C.; Salmon, J.-M.; Vallier, M.-J.; Günata, Z.; Barre, P. Purification, characterization, and substrate specificity of a novel highly glucose-tolerant β-glucosidase from Aspergillus oryzae. Appl. Environ. Microbiol. 1998, 64, 3607–3614. [Google Scholar] [PubMed]
- Margolles-Clark, E.; Tenkanen, M.; Luonteri, E.; Penttilä, M. Three α-galactosidase genes of Trichoderma reesei cloned by expression in yeast. Eur. J. Biochem. 1996, 240, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Zietsman, A.J.; de Klerk, D.; van Rensburg, P. Coexpression of α-l-arabinofuranosidase and β-glucosidase in Saccharomyces cerevisiae. FEMS Yeast Res. 2011, 11, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Mateo, J.J.; Peris, L.; Ibanez, C.; Maicas, S. Characterization of glycolytic activities from non- Saccharomyces yeasts isolated from bobal musts. J. Ind. Microbiol. Biotechnol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Maicas, S.; Mateo, J.J. Enzyme contribution of non-Saccharomyces yeasts to wine production. Univ. J. Microbiol. Res. 2015, 3, 17–25. [Google Scholar]
- Sabel, A.; Martens, S.; Petri, A.; König, H.; Claus, H. Wickerhamomyces anomalus AS1: A new strain with potential to improve wine aroma. Ann. Microbiol. 2014, 64, 483–491. [Google Scholar] [CrossRef]
- Mendes Ferreira, A.; Climaco, M.C.; Mendes Faia, A. The role of non-Saccharomyces species in releasing glycosidic bound fraction of grape aroma components—A preliminary study. J. Appl Microbiol 2001, 91, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, A.; Bartowsky, E.; Jiranek, V. A survey of glycosidase activities of commercial wine strains of Oenococcus oeni. Int. J. Food Microbiol. 2005, 105, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Boido, E.; Lloret, A.; Medina, K.; Carrau, F.; Dellacassa, E. Effect of β-glycosidase activity of Oenococcus oeni on the glycosylated flavor precursors of Tannat wine during malolactic fermentation. J. Agric. Food Chem. 2002, 50, 2344–2349. [Google Scholar] [CrossRef] [PubMed]
- Ugliano, M.; Genovese, A.; Moio, L. Hydrolysis of wine aroma precursors during malolactic fermentation with four commercial starter cultures of Oenococcus oeni. J. Agric. Food Chem. 2003, 51, 5073–5078. [Google Scholar] [CrossRef] [PubMed]
- D’Incecco, N.; Bartowsky, E.; Kassara, S.; Lante, A.; Spettoli, P.; Henschke, P. Release of glycosidically bound flavour compounds of chardonnay by Oenococcus oeni during malolactic fermentation. Food Microbiol. 2004, 21, 257–265. [Google Scholar] [CrossRef]
- Lerm, E.; Engelbrecht, L.; Du Toit, M. Selection and characterisation of Oenococcus oeni and Lactobacillus plantarum south african wine isolates for use as malolactic fermentation starter cultures. S. Afr. J. Enol. Vitic. 2011, 32. [Google Scholar]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; García-Ruiz, A.; Muñoz-González, C.; Bartolomé, B.; Moreno-Arribas, M.V. Selection and technological potential of Lactobacillus plantarum bacteria suitable for wine malolactic fermentation and grape aroma release. LWT-Food Sci. Technol. 2016, 73, 557–566. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P.A. The ‘buttery’attribute of wine—diacetyl—desirability, spoilage and beyond. Int. J. Food Microbiol. 2004, 96, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Martineau, B.; Henick-Kling, T.; Acree, T. Reassessment of the influence of malolactic fermentation on the concentration of diacetyl in wines. Am. J. Enol. Vitic. 1995, 46, 385–388. [Google Scholar]
- Nielsen, J.C.; Richelieu, M. Control of flavor development in wine during and after malolactic fermentation by Oenococcus oeni. Appl. Environ. Microbiol. 1999, 65, 740–745. [Google Scholar] [PubMed]
- Mills, D.A.; Rawsthorne, H.; Parker, C.; Tamir, D.; Makarova, K. Genomic analysis of Oenococcus oeni psu-1 and its relevance to winemaking. FEMS Microbiol Rev. 2005, 29, 465–475. [Google Scholar] [PubMed]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Oelofse, A.; Pretorius, I.; Du Toit, M. Significance of Brettanomyces and Dekkera during winemaking: A synoptic review. S. Afr. J. Enol. Vitic. 2008, 29, 128–144. [Google Scholar]
- Steensels, J.; Daenen, L.; Malcorps, P.; Derdelinckx, G.; Verachtert, H.; Verstrepen, K.J. Brettanomyces yeasts—From spoilage organisms to valuable contributors to industrial fermentations. Int. J. Food Microbiol. 2015, 206, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Edlin, D.A.; Narbad, A.; Dickinson, J.R.; Lloyd, D. The biotransformation of simple phenolic compounds by Brettanomyces anomalus. FEMS Microbiol. Lett. 1995, 125, 311–315. [Google Scholar] [CrossRef]
- Godoy, L.; Martínez, C.; Carrasco, N.; Ganga, M.A. Purification and characterization of a p-coumarate decarboxylase and a vinylphenol reductase from Brettanomyces bruxellensis. Int. J. Food Microbiol. 2008, 127, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Sangorrín, M.; García, V.; Lopes, C.; Sáez, J.; Martínez, C.; Ganga, M. Molecular and physiological comparison of spoilage wine yeasts. J. Appl. Microbiol. 2013, 114, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Montealegre, R.R.; Peces, R.R.; Vozmediano, J.C.; Gascueña, J.M.; Romero, E.G. Phenolic compounds in skins and seeds of ten grape Vitis vinifera varieties grown in a warm climate. J. Food Compos. Anal. 2006, 19, 687–693. [Google Scholar] [CrossRef]
- Bautista-Ortín, A.; Fernández-Fernández, J.; López-Roca, J.; Gómez-Plaza, E. The effects of enological practices in anthocyanins, phenolic compounds and wine colour and their dependence on grape characteristics. J. Food Compos. Anal. 2007, 20, 546–552. [Google Scholar] [CrossRef]
- Blondin, B.; Ratomahenina, R.; Arnaud, A.; Galzy, P. Purification and properties of the β-glucosidase of a yeast capable of fermenting cellobiose to ethanol: Dekkera intermedia van der walt. Eur. J. Appl. Microbiol. Biotechnol. 1983, 17, 1–6. [Google Scholar] [CrossRef]
- Moon, H.; Kim, J.-S.; Oh, K.-K.; Kim, S.W.; Hong, S.-I. Kinetic modeling of simultaneous saccharification and fermentation for ethanol production using steam-exploded wood with glucose- and cellobiose-fermenting yeast, Brettanomyces custersii. J. Microbiol. Biotechnol. 2001, 11, 598–606. [Google Scholar]
- Vanderhaegen, B.; Neven, H.; Coghe, S.; Verstrepen, K.; Derdelinckx, G.; Verachtert, H. Bioflavoring and beer refermentation. Appl. Microbiol. Biotechnol. 2003, 62, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Daenen, L.; Saison, D.; Sterckx, F.; Delvaux, F.; Verachtert, H.; Derdelinckx, G. Screening and evaluation of the glucoside hydrolase activity in Saccharomyces and Brettanomyces brewing yeasts. J. Appl. Microbiol. 2008, 104, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Dweck, H.K.; Ebrahim, S.A.; Farhan, A.; Hansson, B.S.; Stensmyr, M.C. Olfactory proxy detection of dietary antioxidants in Drosophila. Curr. Biol. 2015, 25, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Sturm, M.E.; Assof, M.; Fanzone, M.; Martinez, C.; Ganga, M.; Jofré, V.; Ramirez, M.; Combina, M. Relation between coumarate decarboxylase and vinylphenol reductase activity with regard to the production of volatile phenols by native Dekkera bruxellensis strains under ‘wine-like’conditions. Int. J. Food Microbiol. 2015, 206, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Pagliara, D.; Piccininno, T.; Tarantino, F.; Ciardulli, W.; Malfeito-Ferreira, M.; Loureiro, V. The effect of sugar concentration and temperature on growth and volatile phenol production by Dekkera bruxellensis in wine. FEMS Yeast Res. 2008, 8, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.; Pereira-da-Silva, S.; Tavares, M.; Malfeito-Ferreira, M.; Loureiro, V. Factors affecting the production of 4-ethylphenol by the yeast Dekkera bruxellensis in enological conditions. Food Microbiol. 2003, 20, 377–384. [Google Scholar] [CrossRef]
- Vigentini, I.; Romano, A.; Compagno, C.; Merico, A.; Molinari, F.; Tirelli, A.; Foschino, R.; Volonterio, G. Physiological and oenological traits of different Dekkera/Brettanomyces bruxellensis strains under wine-model conditions. FEMS Yeast Res. 2008, 8, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Agnolucci, M.; Rea, F.; Sbrana, C.; Cristani, C.; Fracassetti, D.; Tirelli, A.; Nuti, M. Sulphur dioxide affects culturability and volatile phenol production by Brettanomyces/Dekkera bruxellensis. Int. J. Food Microbiol. 2010, 143, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Laforgue, R.; Lonvaud-Funel, A. Hydroxycinnamic acid decarboxylase activity of brettanomyces bruxellensis involved in volatile phenol production: Relationship with cell viability. Food Microbiol. 2012, 32, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Serpaggi, V.; Remize, F.; Recorbet, G.; Gaudot-Dumas, E.; Sequeira-Le Grand, A.; Alexandre, H. Characterization of the “viable but nonculturable”(VBNC) state in the wine spoilage yeast Brettanomyces. Food Microbiol. 2012, 30, 438–447. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Di Toro, M.R.; Grieco, F.; Michelotti, V.; Salma, M.; Lamontanara, A.; Russo, P.; Orrù, L.; Alexandre, H.; Spano, G. Viable but not culturable (VBNC) state of Brettanomyces bruxellensis in wine: New insights on molecular basis of VBNC behaviour using a transcriptomic approach. Food Microbiol. 2016, 59, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Carmona, L.; Varela, J.; Godoy, L.; Ganga, M.A. Comparative proteome analysis of Brettanomyces bruxellensis under hydroxycinnamic acid growth. Electron. J. Biotechnol. 2016, 23, 37–43. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E. Bioconversions of ferulic acid, an hydroxycinnamic acid. Crit. Rev. Microbiol. 2006, 32, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Cavin, J.; Andioc, V.; Etievant, P.; Divies, C. Ability of wine lactic acid bacteria to metabolize phenol carboxylic acids. Am. J. Enol. Vitic. 1993, 44, 76–80. [Google Scholar]
- de las Rivas, B.; Rodríguez, H.; Curiel, J.A.; Landete, J.M.; Munoz, R. Molecular screening of wine lactic acid bacteria degrading hydroxycinnamic acids. J. Agric. Food Chem. 2008, 57, 490–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, P.J.; Lee, T.H.; Henschke, P. Ability of lactic acid bacteria to produce N-heterocycles causing mousy off-flavour in wine. Aust. J. Grape Wine Res. 2001, 7, 160–167. [Google Scholar] [CrossRef]
- Costello, P.J.; Henschke, P.A. Mousy off-flavor of wine: Precursors and biosynthesis of the causative N-heterocycles 2-ethyltetrahydropyridine, 2-acetyltetrahydropyridine, and 2-acetyl-1-pyrroline by Lactobacillus hilgardii DSM 20176. J. Agric. Food Chem 2002, 50, 7079–7087. [Google Scholar] [CrossRef] [PubMed]
- Pozo-Bayón, M.Á.; Muñoz-González, C.; Esteban-Fernández, A. Wine preference and wine aroma perception. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé-Suáldea, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 139–162. [Google Scholar]
- The Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [Green Version]
- Dewhirst, F.E.; Chen, T.; Izard, J.; Paster, B.J.; Tanner, A.C.; Yu, W.-H.; Lakshmanan, A.; Wade, W.G. The human oral microbiome. J. Bacteriol. 2010, 192, 5002–5017. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Fernández, A.; Rocha-Alcubilla, N.; Muñoz-González, C.; Moreno-Arribas, M.V.; Pozo-Bayón, M.Á. Intra-oral adsorption and release of aroma compounds following in-mouth wine exposure. Food Chem. 2016, 205, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Mayr, C.M.; Parker, M.; Baldock, G.A.; Black, C.A.; Pardon, K.H.; Williamson, P.O.; Herderich, M.J.; Francis, I.L. Determination of the importance of in-mouth release of volatile phenol glycoconjugates to the flavor of smoke-tainted wines. J. Agric. Food Chem. 2014, 62, 2327–2336. [Google Scholar] [CrossRef] [PubMed]
- Buettner, A. Influence of human saliva on odorant concentrations. 2. Aldehydes, alcohols, 3-alkyl-2-methoxypyrazines, methoxyphenols, and 3-hydroxy-4, 5-dimethyl-2 (5H)-furanone. J. Agric. Food Chem. 2002, 50, 7105–7110. [Google Scholar] [CrossRef] [PubMed]
- Buettner, A.; Beer, A.; Hannig, C.; Settles, M.; Schieberle, P. Physiological and analytical studies on flavor perception dynamics as induced by the eating and swallowing process. Food Qual. Prefer. 2002, 13, 497–504. [Google Scholar] [CrossRef]
- Lasekan, O. A comparative analysis of the influence of human salivary enzymes on odorant concentration in three palm wines. Molecules 2013, 18, 11809–11823. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, C.; Cueva, C.; Pozo-Bayón, M.Á.; Moreno-Arribas, M.V. Ability of human oral microbiota to produce wine odorant aglycones from odourless grape glycosidic aroma precursors. Food Chem 2015, 187, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Carrau, F.; Gaggero, C.; Aguilar, P.S. Yeast diversity and native vigor for flavor phenotypes. Trends Biotechnol. 2015, 33, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Borneman, A.R.; Pretorius, I.S.; Chambers, P.J. Comparative genomics: A revolutionary tool for wine yeast strain development. Curr. Opin. Biotechnol. 2013, 24, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Bartowsky, E.J.; Borneman, A.R. Genomic variations of Oenococcus oeni strains and the potential to impact on malolactic fermentation and aroma compounds in wine. Appl. Microbiol. Biotechnol. 2011, 92, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Tronchoni, J.; Quirós, M.; Morales, P. Genetic improvement and genetically modified microorganisms. In Wine Safety, Consumer Preference, and Human Health; Moreno-Arribas, M.V., Bartolomé-Suáldea, B., Eds.; Springer: Cham, Switzerland, 2016; pp. 71–96. [Google Scholar]
- Pretorius, I.S. Synthetic genome engineering forging new frontiers for wine yeast. Crit. Rev. Biotechnol. 2016, 37, 112–136. [Google Scholar] [CrossRef] [PubMed]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement. Molecules 2017, 22, 189. https://doi.org/10.3390/molecules22020189
Belda I, Ruiz J, Esteban-Fernández A, Navascués E, Marquina D, Santos A, Moreno-Arribas MV. Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement. Molecules. 2017; 22(2):189. https://doi.org/10.3390/molecules22020189
Chicago/Turabian StyleBelda, Ignacio, Javier Ruiz, Adelaida Esteban-Fernández, Eva Navascués, Domingo Marquina, Antonio Santos, and M. Victoria Moreno-Arribas. 2017. "Microbial Contribution to Wine Aroma and Its Intended Use for Wine Quality Improvement" Molecules 22, no. 2: 189. https://doi.org/10.3390/molecules22020189