
Phenylethanol Glycosides from Cistanche tubulosa Suppress Hepatic Stellate Cell Activation and Block the Conduction of Signaling Pathways in TGF-β1/smad as Potential Anti-Hepatic Fibrosis Agents


Abstract
:1. Introduction

2. Results
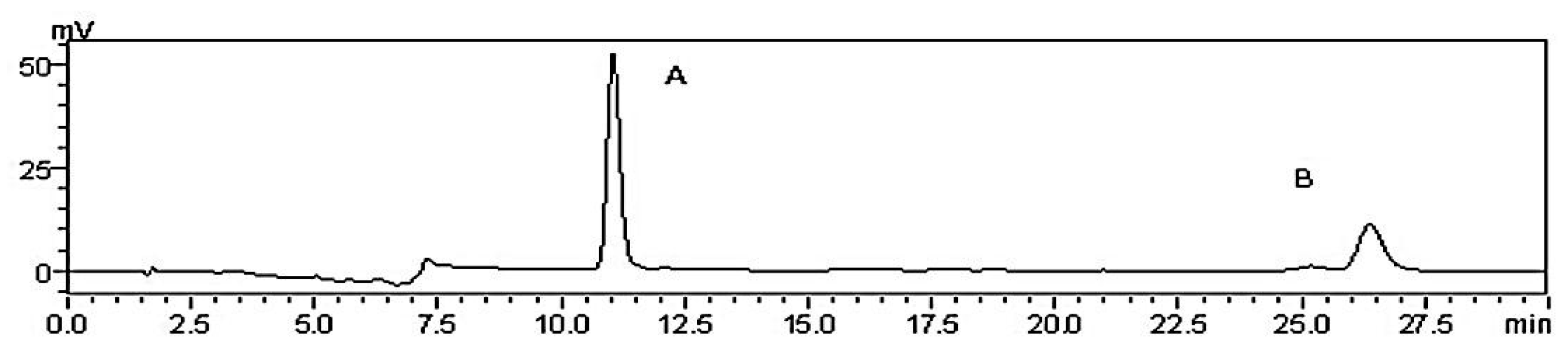
2.1. Quantitative Determination of CPhGs
2.2. Inhibitory Activities of CPhGs, Echinacoside and Acteoside on HSC-T6

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (μg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 500 | 250 | 125 | 62.5 | 31.25 | 15.625 | 7.8125 | 3.90625 | Control | |
| CPhGs OD | 0.18 | 0.23 | 0.30 | 0.45 | 0.50 | 0.57 | 0.62 | 0.67 | 0.68 |
| IR (%) | 73.40 | 66.40 | 55.76 | 33.42 | 26.93 | 16.32 | 9.02 | 1.10 | |
| Echinacoside OD | 0.29 | 0.33 | 0.35 | 0.36 | 0.42 | 0.46 | 0.49 | 0.51 | 0.52 |
| IR (%) | 44.19 | 37.57 | 31.91 | 30.90 | 18.95 | 11.71 | 6.33 | 1.44 | |
| acteoside OD | 0.22 | 0.26 | 0.31 | 0.49 | 0.65 | 0.73 | 0.80 | 1.03 | 1.78 |
| IR (%) | 87.41 | 85.13 | 82.48 | 72.20 | 63.60 | 58.83 | 54.83 | 42.13 | |

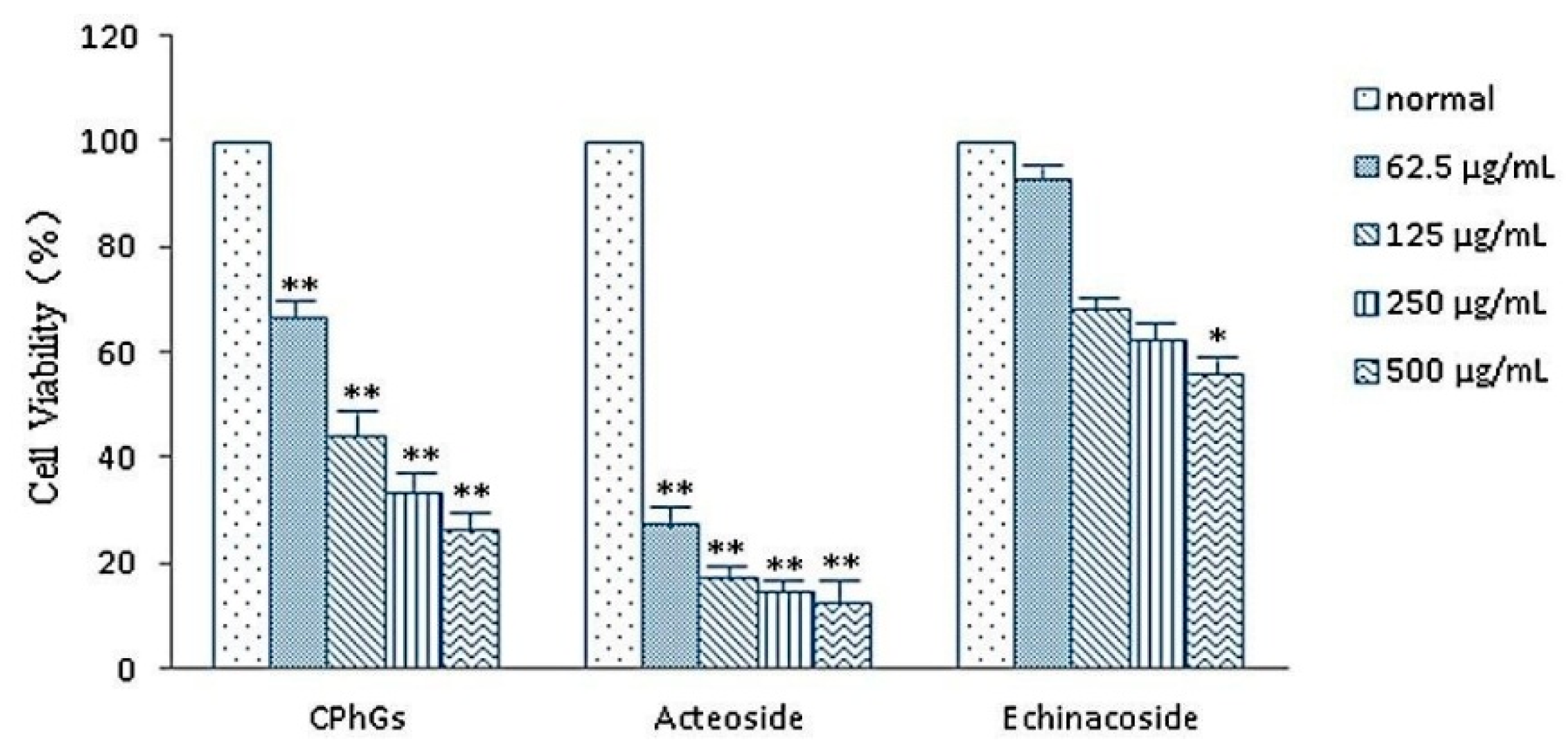
2.3. Cell Toxicity of CPhGs, Echinacoside and Acteoside on HSC
| Group | n | LDH (U/L) |
|---|---|---|
| Control | 4 | 226.22 ± 26.82 |
| CPhGs 100 μg/mL | 4 | 240.58 ± 17.75 * |
| 50 μg/mL | 4 | 239.21 ± 2.11 * |
| 25 μg/mL | 4 | 230.50 ± 6.64 * |
| Echinacoside 500 μg/mL | 4 | 240.01 ± 4.39 * |
| 250 μg/mL | 4 | 236.56 ± 13.06 * |
| 125 μg/mL | 4 | 233.02 ± 5.85 * |
| Acteoside 6 μg/mL | 4 | 240.77 ± 6.43 * |
| 3 μg/mL | 4 | 240.13 ± 10.04 * |
| 1.5 μg/mL | 4 | 234.99 ± 7.92 * |
2.4. Effect of CPhGs, Echinacoside and Acteoside on HSC Proliferation Induced by TGF-β1
| Group | Absorbance Inhibition (OD) (%) | Group | Absorbance Inhibition (OD) (%) | Group | Absorbance Inhibition (OD) (%) | |||
|---|---|---|---|---|---|---|---|---|
| Control TGF-β1 | 0.91 ± 0.05 1.30 ± 0.05 ** | Control TGF-β1 | 0.53 ± 0.06 0.74 ± 0.05 ** | Control TGF-β1 | 0.80 ± 0.07 0.96 ± 0.03 ** | |||
| TC 100 μg/mL | 1.14 ± 0.14 # | 41.60 | TE 500 μg/mL | 0.61 ± 0.01 ## | 55.58 | TA 6.0 μg/mL | 0.87 ± 0.03 ## | 53.44 |
| TC 50 μg/mL | 1.15 ± 0.05 # | 38.40 | TE 250 μg/mL | 0.67 ± 0.03 # | 30.89 | TA 3.0 μg/mL | 0.89 ± 0.04 # | 45.50 |
| TC 25 μg/mL | 1.22 ± 0.08 | 19.68 | TE 125 μg/mL | 0.68 ± 0.02 # | 25.83 | TA 1.5 μg/mL | 0.93 ± 0.04 | 20.92 |
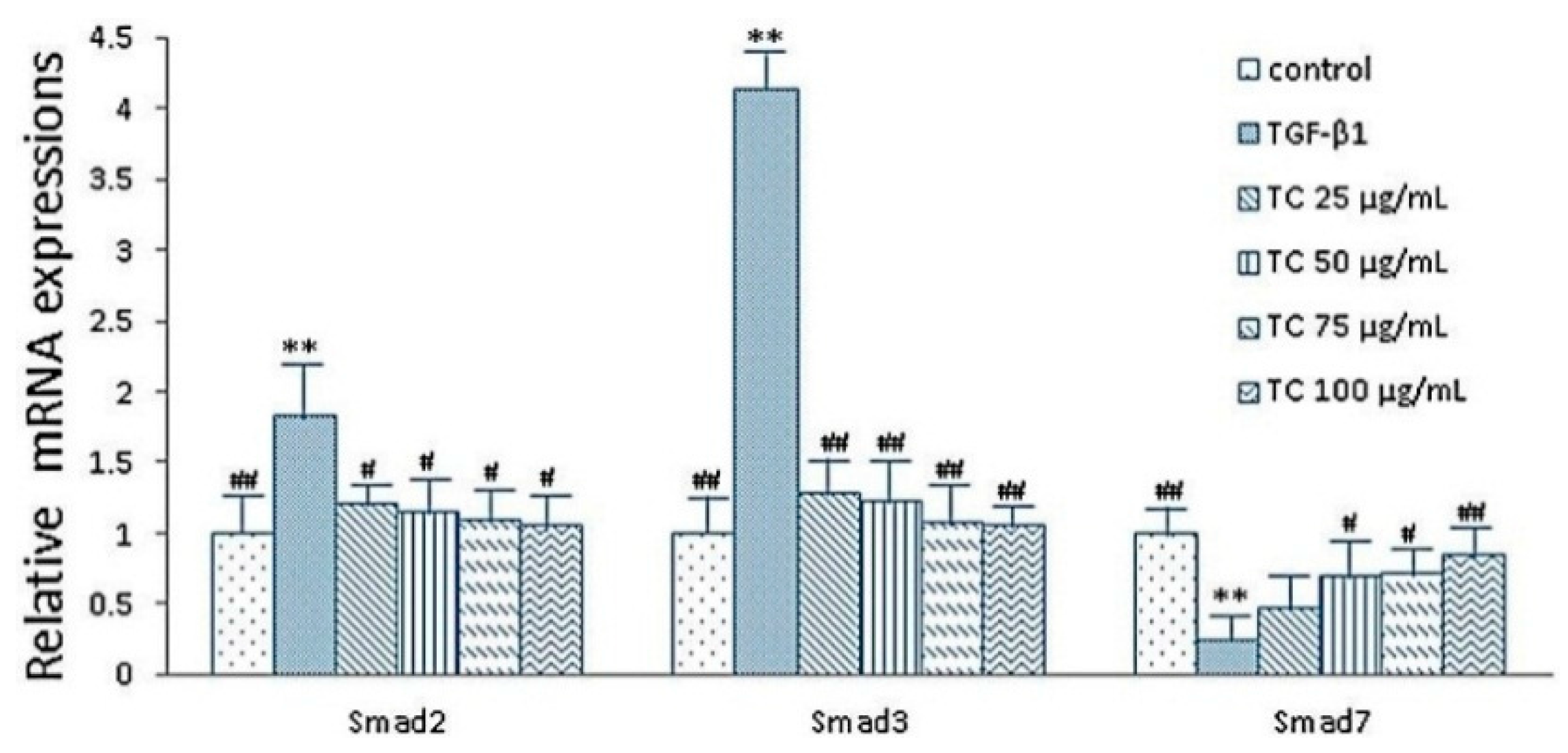
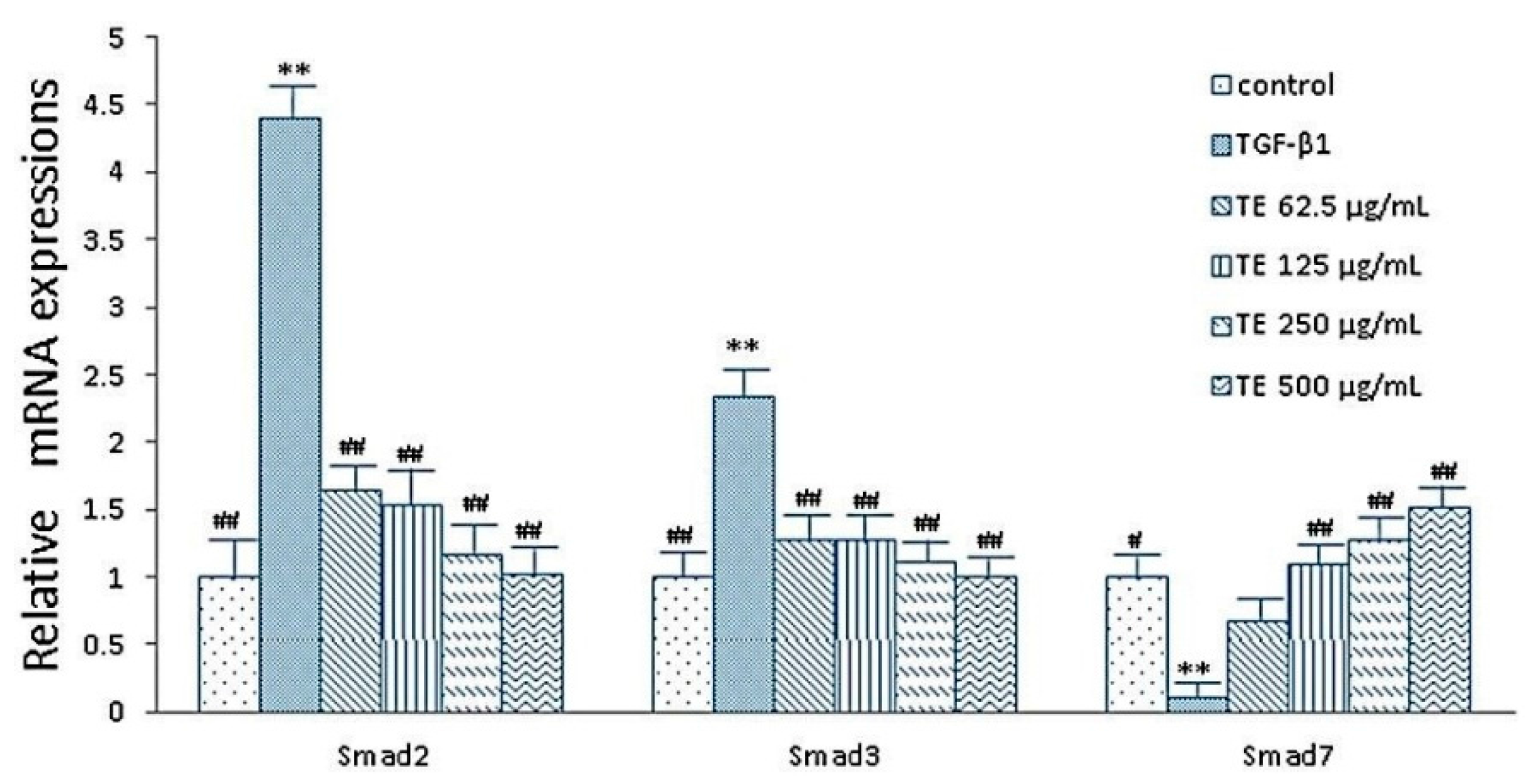
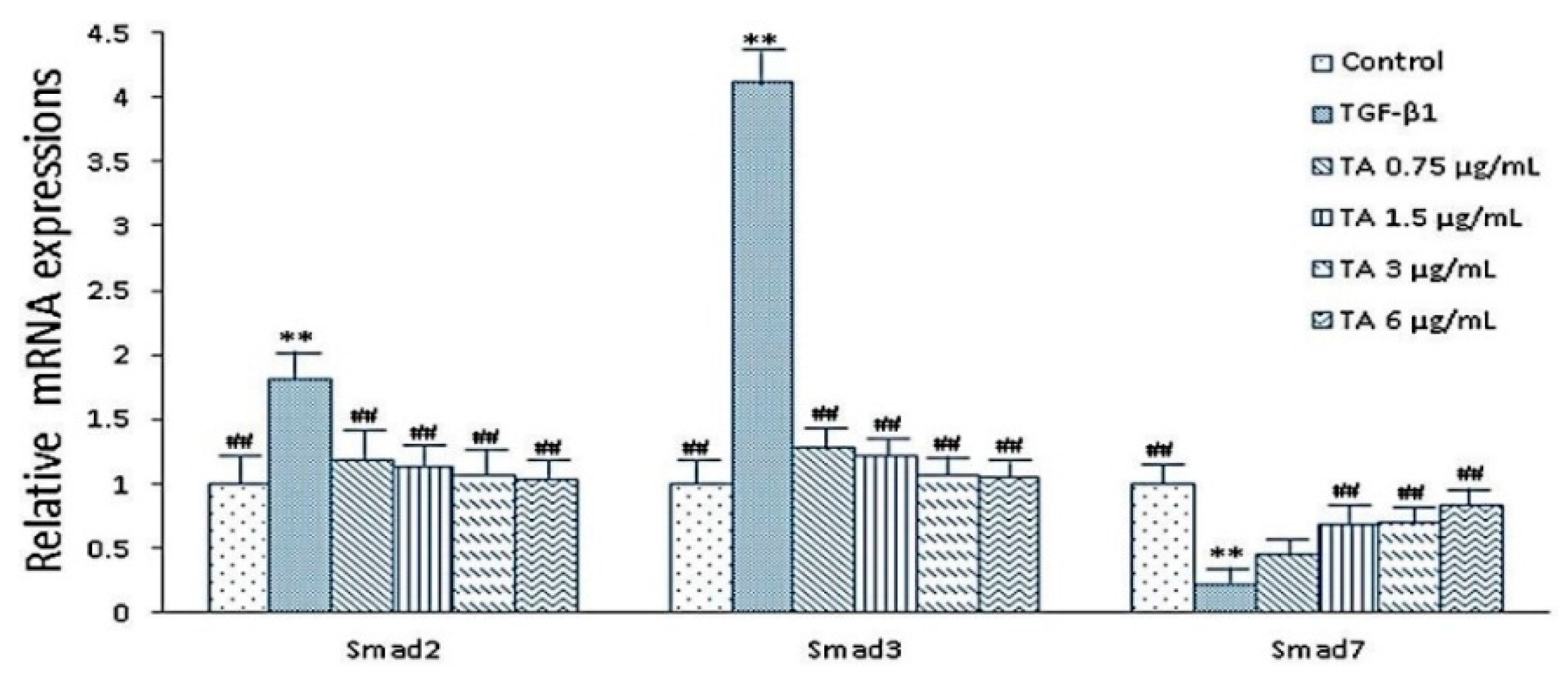
2.5. Expressions of smad2, smad3 and smad7 mRNA after CPhGs, echinacoside and acteoside intervention on HSCs



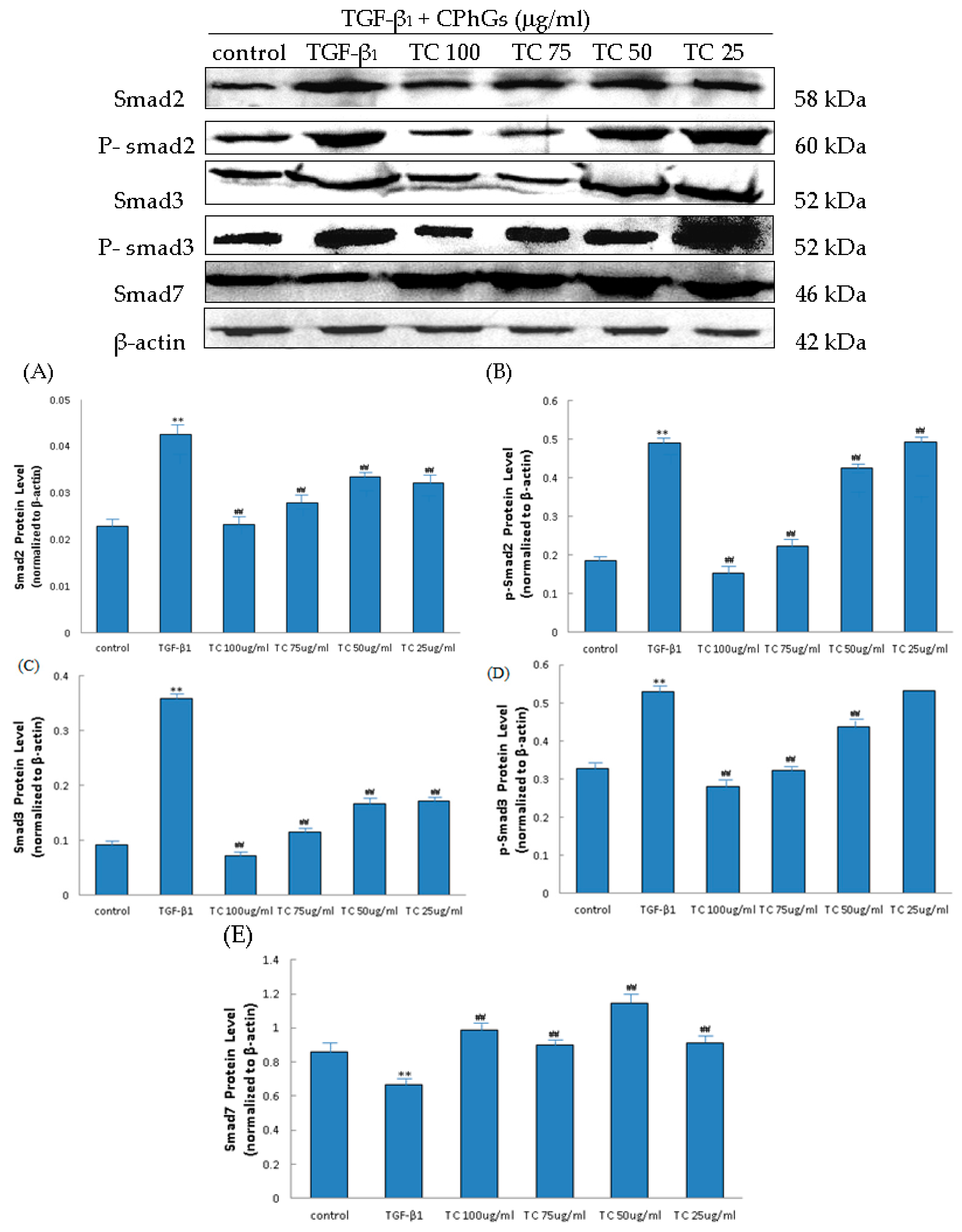
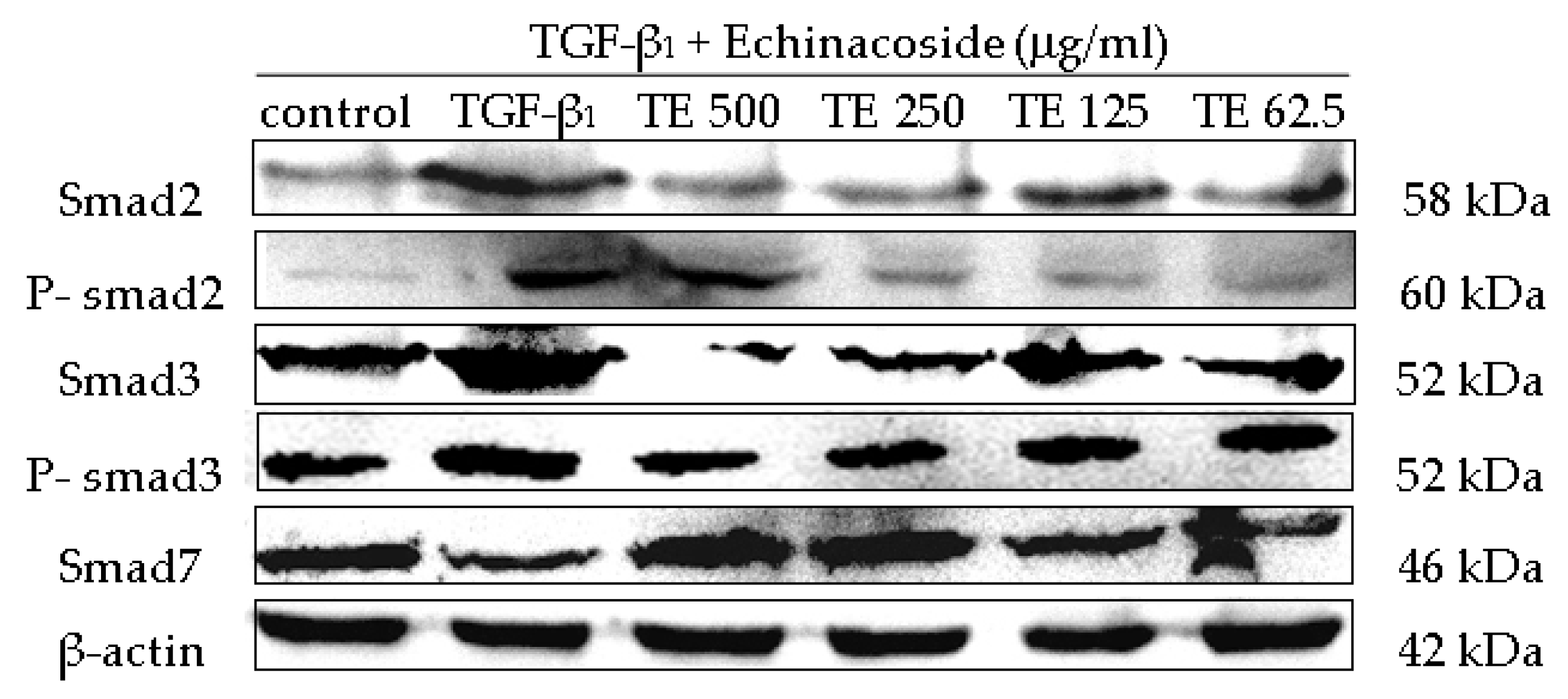
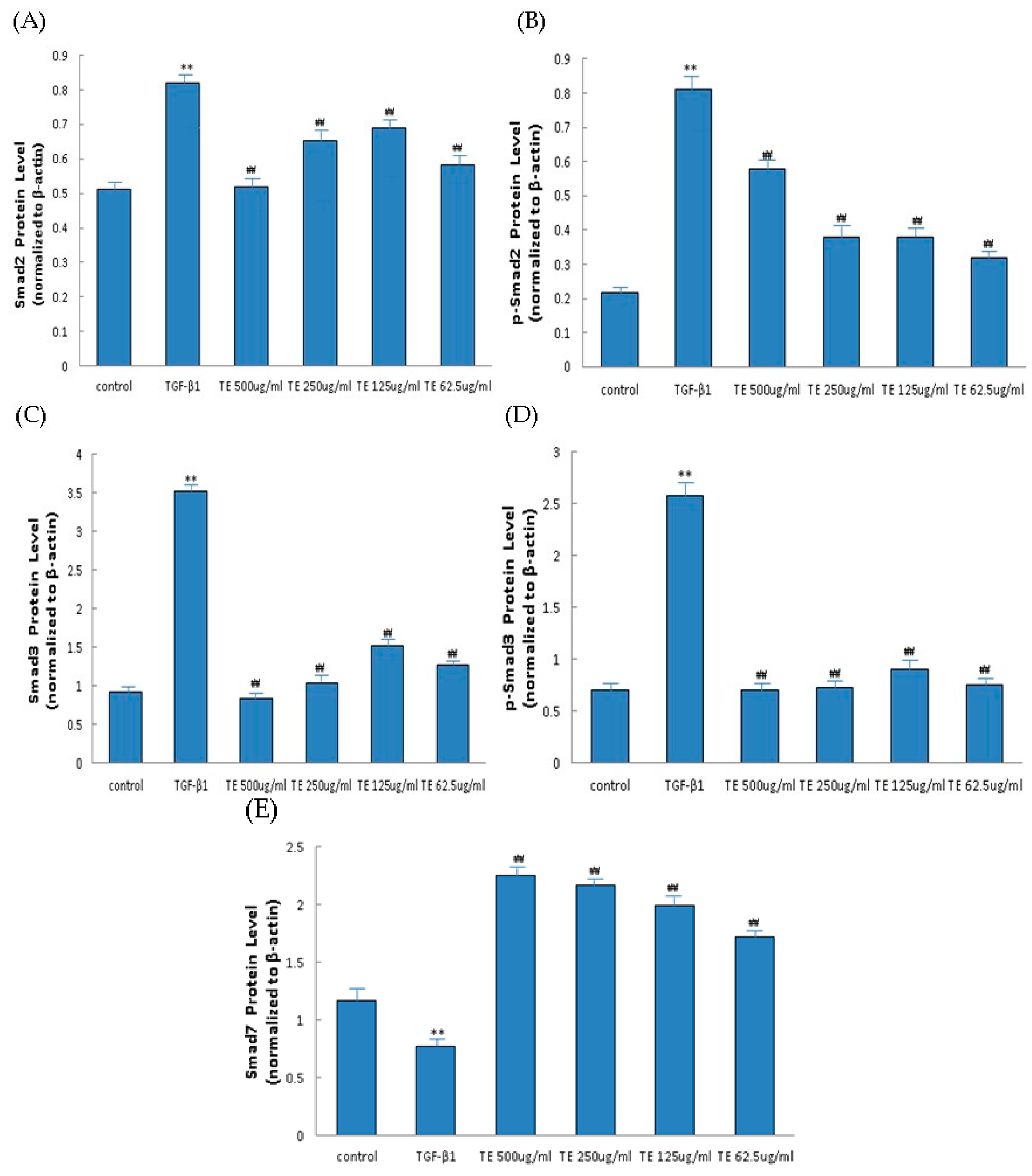
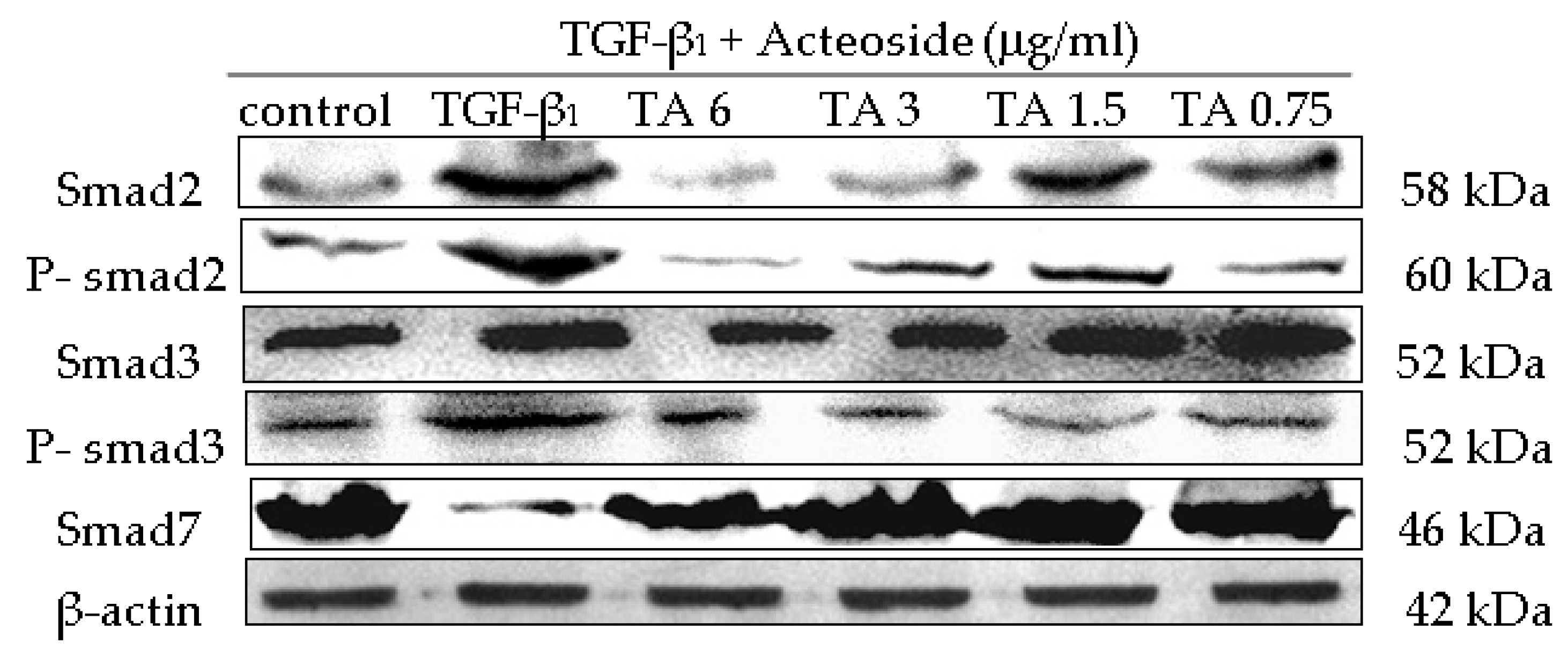
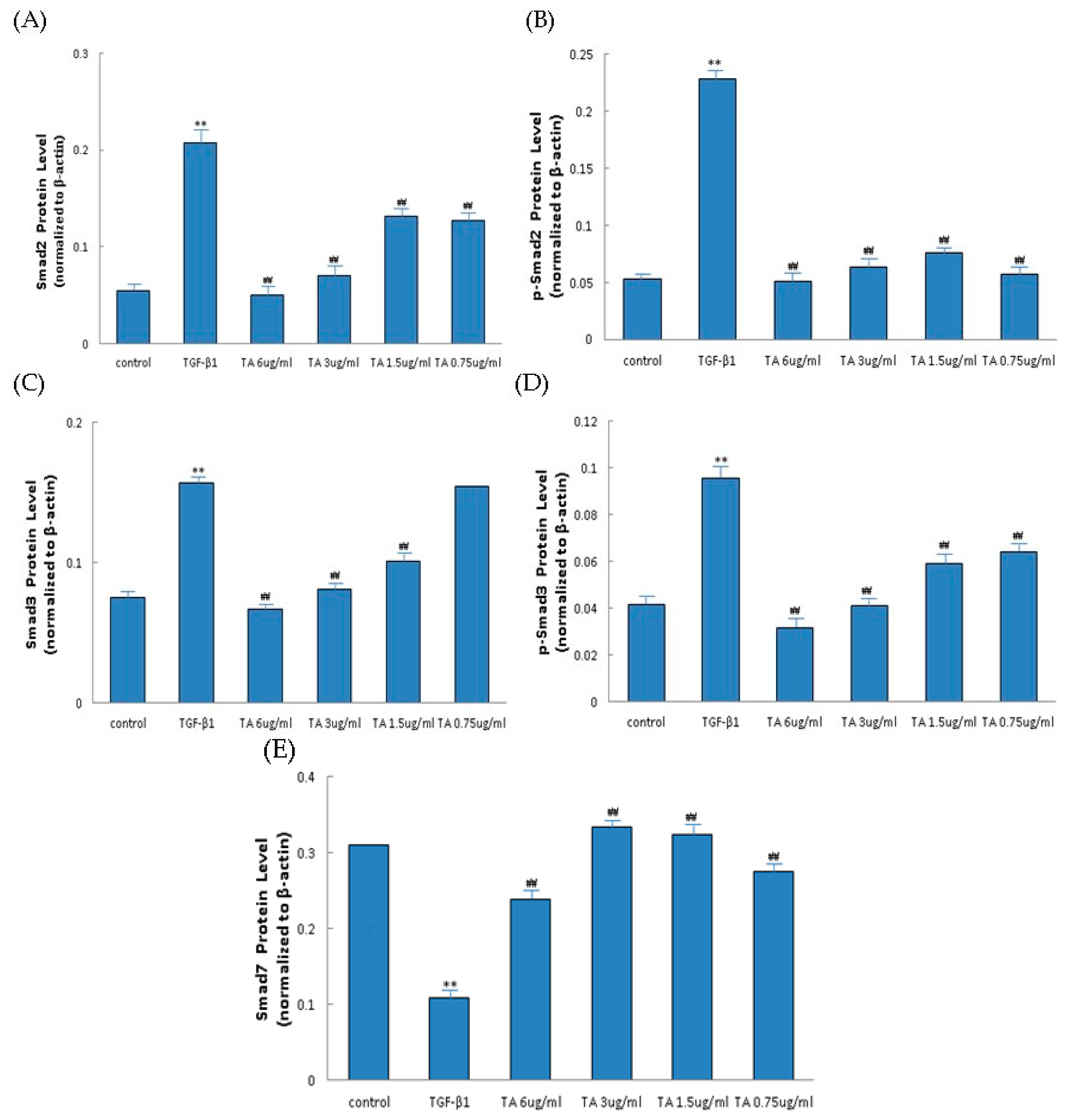
2.6. Influence of CPhGs, Echinacoside and Acteoside on TGF-β1/smad Signaling Pathway in HSC





3. Discussion
4. Experimental Section
4.1. Chemicals and Reagents
4.2. Plant Material
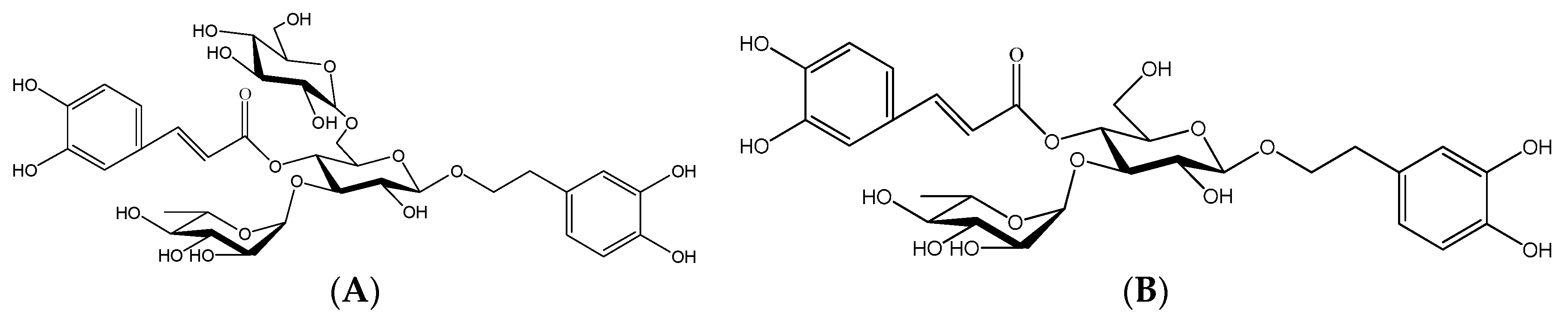
4.3. Isolation and Purification of CPhGs and Its Compounds

4.4. Quantitative Determination of CPhGs
4.5. Cell Culture
4.6. Determination of IC50 Values
4.7. The Cell Proliferative Inhibition Effects and Cell Viability
4.8. Anti-Proliferative Activities by TGF-β1
4.9. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
| Target Gene Primer Sequence (Forward/Reverse) (Rat) | GenBank | ||
|---|---|---|---|
| Smad2 | F: ACGGCTTTACAGATCCATCG | R: GCCAGAAGAGCAGCAAATTC | NM_0012774501 |
| Smad3 | F: GGCCATGTTGGTTTATGGAG | R: CCAGGGTGAAGATGACAGGT | NM_013095.3 |
| Smad7 | F: GTGGCATACTGGGAGGAGAA | R: TGGAGAAACCAGGGAACACT | NM_030858.1 |
| β-actin | F: TAAGGCCAACCGTGAAAAGATG | R: AGAGGCATACAGGGACAACACA | NM_031144.3 |
4.10. Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CPhGs | Phenylethanol glycosides from cistanche |
| BSA | Bovine serum albumin |
| HSC | Hepatic stellate cells |
| mAb | Monoclonal antibody |
| TGF-β1 | Transforming growth factor-β1 |
| IC50 | Inhibition of 50% of cell growth activity |
| ECM | Extracellular matrix |
| TCHM | Chinese and herb medicine |
| TC | TGF-β1 + CPhGs |
| TE | TGF-β1 + echinacoside |
| TA | TGF-β1 + acteoside |
| p-smad2 | phosphorylation of smad2 |
| p-smad3 | phosphorylation of smad3 |
| LDH | Lactate dehydrogenase |
| HPLC | Liquid chromatography |
| RIPA | Radioimmunoprecipitation assay |
| RT-PCR | Reverse transcriptase polymerase chain reaction |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
References
- Subramoniam, A.; Pushpangadan, P. Development of phytomedicine for liver diseases. Ind. J. Pharmacol. 1999, 31, 166–175. [Google Scholar]
- Friedman, S.L. Mechanisms of disease: Mechanisms of hepatic fibrosis and therapeutic implications. Nat. Clin. Pract. Gastroenterol Hepatol. 2004, 1, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Roll, F.J.; Boyles, J.; Bissell, D.M. Hepatic lipocytes: The principal collagen-producing cells of normal rat liver. Proc. Natl. Acad. Sci. USA 1985, 82, 8681–8685. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Bansal, M.B. Reversal of hepatic fibrosis—Fact or fantasy? Hepatology 2006, 43, S82–S88. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Ahmad, R. Understanding the mechanism of hepatic fibrosis and potential therapeutic approaches. Saudi J. Gastroenterol. 2012, 18, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Peter, H.; Raven; Zhang, L.B. Orobanchaceae Ventenat. In Flora of China, 23rd ed.; Editorial Committee of Flora of China: Chinese Academy of Sciences, Beijing, China, 2013; pp. 1–16. [Google Scholar]
- Chinese Pharmacopoeia Commission of Sanitary Ministry of People’s Republic of China. Chinese Pharmacopoeia; Part 1; Chemical Industry Publishing House: Beijing, China, 2010; p. 126. [Google Scholar]
- Li, J.; Huang, D.; He, L. Effect of roucongrong (Herba Cistanches Deserticolae) on reproductive toxicity in mice induced by glycoside of Leigongteng (Radix et Rhizoma Tripterygii). J. Tradit. Chin. Med. 2014, 34, 324–332. [Google Scholar] [CrossRef]
- Xing, Y.; Liao, J.; Tang, Y.; Zhang, P.; Tan, C.; Ni, H.; Wu, X.; Li, N.; Jia, X. ACE and platelet aggregation inhibitors from Tamarix hohenackeri Bunge (host plant of Herba Cistanches) growing in Xinjiang. Pharmacogn. Mag. 2014, 10, 111–118. [Google Scholar] [PubMed]
- Jia, Y.; Guan, Q.; Jiang, Y.; Salh, B.; Guo, Y.; Tu, P.; Du, C. Amelioration of dextran sulphate sodium-induced colitis in mice by echinacoside-enriched extract of Cistanche tubulosa. Phytother. Res. 2014, 28, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.S.; Ko, K.M. Herba Cistanches stimulates cellular glutathione redox cycling by reactive oxygen species generated from mitochondrial respiration in H9c2 cardiomyocytes. Pharm. Biol. 2013, 51, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.; Clynes, M. Acid phosphatase: Endpoint for in vitro toxicity tests. Vitro Cell. Dev. Biol. 1991, 27, 183–184. [Google Scholar] [CrossRef]
- Lechuga, C.G.; Hernandez-Nazara, Z.H.; Dominguez Rosales, J.A.; Morris, E.R.; Rincon, A.R.; Rivas-Estilla, A.M.; Esteban-Gamboa, A.; Rojkind, M. TGF-β1 modulates matrix metalloproteinase-13 expression in hepatic stellate cells by complex mechanisms involving p38MAPK, PI3-kinase, AKT, and p70S6k. Am. J. Physiol. Gastrointest Liver Physiol. 2004, 287, G974–G987. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, J.; Tang, S.Y.; Nguyen, D.; Alliston, T. Load regulates bone formation and Sclerostin expression through a TGFβ-dependent mechanism. PLoS ONE 2013, 8, e53813. [Google Scholar] [CrossRef] [PubMed]
- Pessah, M.; Marais, J.; Prunier, C.; Ferrand, N.; Lallemand, F.; Mauviel, A.; Atfi, A. C-Jun associates with the oncoprotein Ski and suppresses Smad2 transcriptional activity. J. Biol. Chem. 2002, 277, 29094–29100. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Yu, R.T.; Subramaniam, N.; Sherman, M.H.; Wilson, C.; Rao, R.; Leblanc, M.; Coulter, S.; He, M.; Scott, C.; et al. A Vitamin D receptor/SMAD genomic circuit gates hepatic fibrotic response. Cell 2013, 153, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.R.; Fang, B.W.; Lou, J.S. Effects of Fufang Biejia Ruangan Pills on hepatic fibrosis in vivo and in vitro. World J. Gastroenterol. 2013, 19, 5326–5333. [Google Scholar] [CrossRef] [PubMed]
- Sakaida, I.; Hironaka, K.; Kimura, T.; Terai, S.; Yamasaki, T.; Okita, K. Herbal medicine Sho-saiko-to (TJ-9) increases expression matrix metalloproteinases (MMPs) with reduced expression of tissue inhibitor of metalloproteinases (TIMPs) in rat stellate cell. Life Sci. 2004, 74, 2251–2263. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, J.; Tsai, C.; Wang, W.; Chang, D.; Tu, D.G.; Hsieh, H.Y. The role of TGF-β1 and cytokines in the modulation of liver fibrosis by Sho-saiko-to in rat’s bile duct ligated model. J. Ethnopharmacol. 2005, 97, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Bolarin, D.M.; Azinge, E.C. Biochemical markers, extracellular components in liver fibrosis and cirrhosis. Nig. Q. J. Hosp. Med. 2007, 17, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Zheng, S.; Lu, Y. Source and role of myofibroblasts in liver fibrosis. Chin. Pharmacol. Bull. 2011, 27, 297–300. [Google Scholar]
- De Lavor, M.S.L.; Binda, N.S.; Fukushima, F.B.; Caldeira, F.M.C.; Da Silva, J.F.; Silva, C.M.O.; de Oliveira, K.M.; Martins Bde, C.; Torres, B.B.; Rosado, I.R.; et al. Ischemia-reperfusion model in rat spinal cord: Cell viability and apoptosis signaling study. Int. J. Clin. Exp. Pathol. 2015, 8, 9941–9949. [Google Scholar] [PubMed]
- Bahadar, H.; Maqbool, F.; Niaz, K.; Abdollahi, M. Toxicity of nanoparticles and an overview of current experimental models. Iran Biomed. J. 2016, 20, 1–11. [Google Scholar] [PubMed]
- Jing, L.; Ma, H.; Fan, P.; Gao, R.; Jia, Z. Antioxidant potential, total phenolic and total flavonoid contents of Rhododendron anthopogonoides and its protective effect on hypoxia-induced injury in PC12 cells. BMC Complement. Altern. Med. 2015, 15, 287. [Google Scholar] [CrossRef] [PubMed]
- Tipoe, G.L.; Leung, T.M.; Liong, E.C.; Lau, T.Y.; Fung, M.L.; Nanji, A.A. Epigallocatechin-3-gallate (EGCG) reduces liver inflammation, oxidative stress and fibrosis in carbon tetrachloride (CCl4)-induced liver injury in mice. Toxicology 2010, 273, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.F.; Huang, R.B.; Zhang, S.J.; Lin, J.; Wei, L.; He, M.; Zhuo, L.; Lin, X. Protective effect of genistein isolated from Hydrocotyle sibthorpioides on hepatic injury and fibrosis induced by chronic alcohol in rats. Toxicol. Lett. 2013, 217, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.X.; Lu, C.H.; Xie, W.F.; Zhang, X.R.; Zhang, Z.B.; Wei, L.X.; Jin, Y.X.; Guo, Y.J. Effects of ribozyme targeting platelet-derived growth factor receptor β subunit gene on the proliferation and apoptosis of hepatic stellate cells in vitro. Chin. Med. J. (Engl.) 2005, 118, 982–988. [Google Scholar] [PubMed]
- Jiang, Y.; Wang, C.; Li, Y.Y.; Wang, X.C.; An, J.D.; Wang, Y.J.; Wang, X.J. Mistletoe alkaloid fractions alleviates carbon tetrachloride-induced liver fibrosis through inhibition of hepatic stellate cell activation via TGF-β/Smad interference. J. Ethnopharmacol. 2014, 158, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, M.; Nishitoh, H.; Amagasa, T.; Miyazono, K.; Takagi, M.; Ichijo, H. Identification of important regions in the cytoplasmic juxtamembrane domain of type I receptor that separate signaling pathways of transforming growth factor-beta. J. Biol. Chem. 1996, 271, 2769–2775. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Smad phosphoisoform signals in acute and chronic liver injury: Similarities and differences between epithelial and mesenchymal cells. Cell Tissue Res. 2012, 347, 225–243. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Wrana, J.L. TGF-β and the Smad signal transduction pathway. Biochem. Cell Biol. 2002, 80, 605–622. [Google Scholar] [CrossRef] [PubMed]
- Briones-Orta, M.A.; Tecalco-Cruz, A.C.; Sosa-Garrocho, M.; Caligaris, C.; Macías-Silva, M. Inhibitory Smad7: Emerging roles in health and disease. Curr. Mol. Pharmacol. 2011, 4, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Dooley, S.; Hamzavi, J.; Breitkopf, K.; Wiercinska, E.; Said, H.M.; Lorenzen, J.; Ten Dijke, P.; Gressner, A.M. Smad7 prevents activation of hepatic stellate cells and liver fibrosis in rats. Gastroenterology 2003, 125, 178–191. [Google Scholar] [CrossRef]
- Ka, S.M.; Yeh, Y.C.; Huang, X.R.; Chao, T.K.; Hung, Y.J.; Yu, C.P.; Lin, T.J.; Wu, C.C.; Lan, H.Y.; Chen, A. Kidney-targeting Smad7 gene transfer inhibits renal TGF-β/MAD homologue (SMAD) and nuclear factor κB (NF-κB) signalling pathways, and improves diabetic nephropathy in mice. Diabetologia 2012, 55, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, T.; Pan, Y.; Ninomiya, K.; Imura, K.; Matsuda, H.; Yoshikawa, M.; Yuan, D.; Muraoka, O. Acylated phenylethanoid oligoglycosides with hepatoprotective activity from the desert plant Cistanche tubulosa. Bioorg. Med. Chem. 2010, 18, 1882–1890. [Google Scholar] [CrossRef] [PubMed]
- Gélinas, S.; Martinoli, M.G. Neuroprotective effect of estradiol and phytoestrogens on MPP+-induced cytotoxicity in neuronal PC12 cells. J. Neurosci. Res. 2002, 70, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bartalis, J.; Halaweish, F.T. In vitro and QSAR studies of cucurbitacins on HepG2 and HSC-T6 liver cell lines. Bioorg. Med. Chem. Lett. 2011, 19, 2757–2766. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Wang, Z.; Kwong, S.Q.; Lui, E.L.H.; Friedman, S.L.; Li, F.R.; Lam, R.W.C.; Zhang, G.C.; Zhang, H.; Ye, T. Inhibition of PDGF, TGF-β, and Abl signaling and reduction of liver fibrosis by the small molecule Bcr-Abl tyrosine kinase antagonist Nilotinib. J. Hepatol. 2011, 55, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, S.-P.; Ma, L.; Zhao, J.; Zhang, S.-L.; Liu, T. Phenylethanol Glycosides from Cistanche tubulosa Suppress Hepatic Stellate Cell Activation and Block the Conduction of Signaling Pathways in TGF-β1/smad as Potential Anti-Hepatic Fibrosis Agents. Molecules 2016, 21, 102. https://doi.org/10.3390/molecules21010102
You S-P, Ma L, Zhao J, Zhang S-L, Liu T. Phenylethanol Glycosides from Cistanche tubulosa Suppress Hepatic Stellate Cell Activation and Block the Conduction of Signaling Pathways in TGF-β1/smad as Potential Anti-Hepatic Fibrosis Agents. Molecules. 2016; 21(1):102. https://doi.org/10.3390/molecules21010102
Chicago/Turabian StyleYou, Shu-Ping, Long Ma, Jun Zhao, Shi-Lei Zhang, and Tao Liu. 2016. "Phenylethanol Glycosides from Cistanche tubulosa Suppress Hepatic Stellate Cell Activation and Block the Conduction of Signaling Pathways in TGF-β1/smad as Potential Anti-Hepatic Fibrosis Agents" Molecules 21, no. 1: 102. https://doi.org/10.3390/molecules21010102