
Potent PPARγ Ligands from Swietenia macrophylla Are Capable of Stimulating Glucose Uptake in Muscle Cells


 ,
,
Abstract
:1. Introduction
2. Results
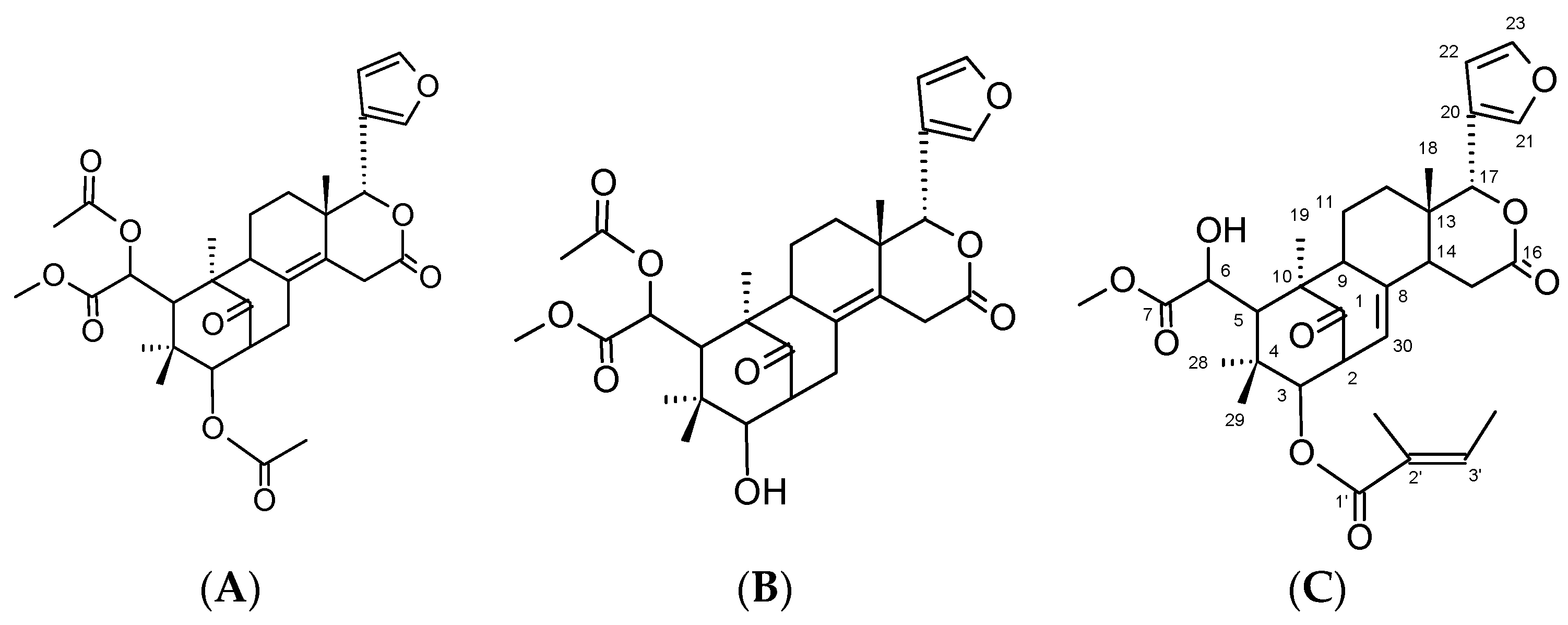
2.1. Identification and Characterization of Compounds

2.2. Binding of 6OA, DS and Sw to PPARγ in an ELISA PPARγ-Binding Assay

2.3. Induction of 3T3-L1 Adipocyte Differentiation in 3T3-L1 Cells by 6OA, DS and Sw


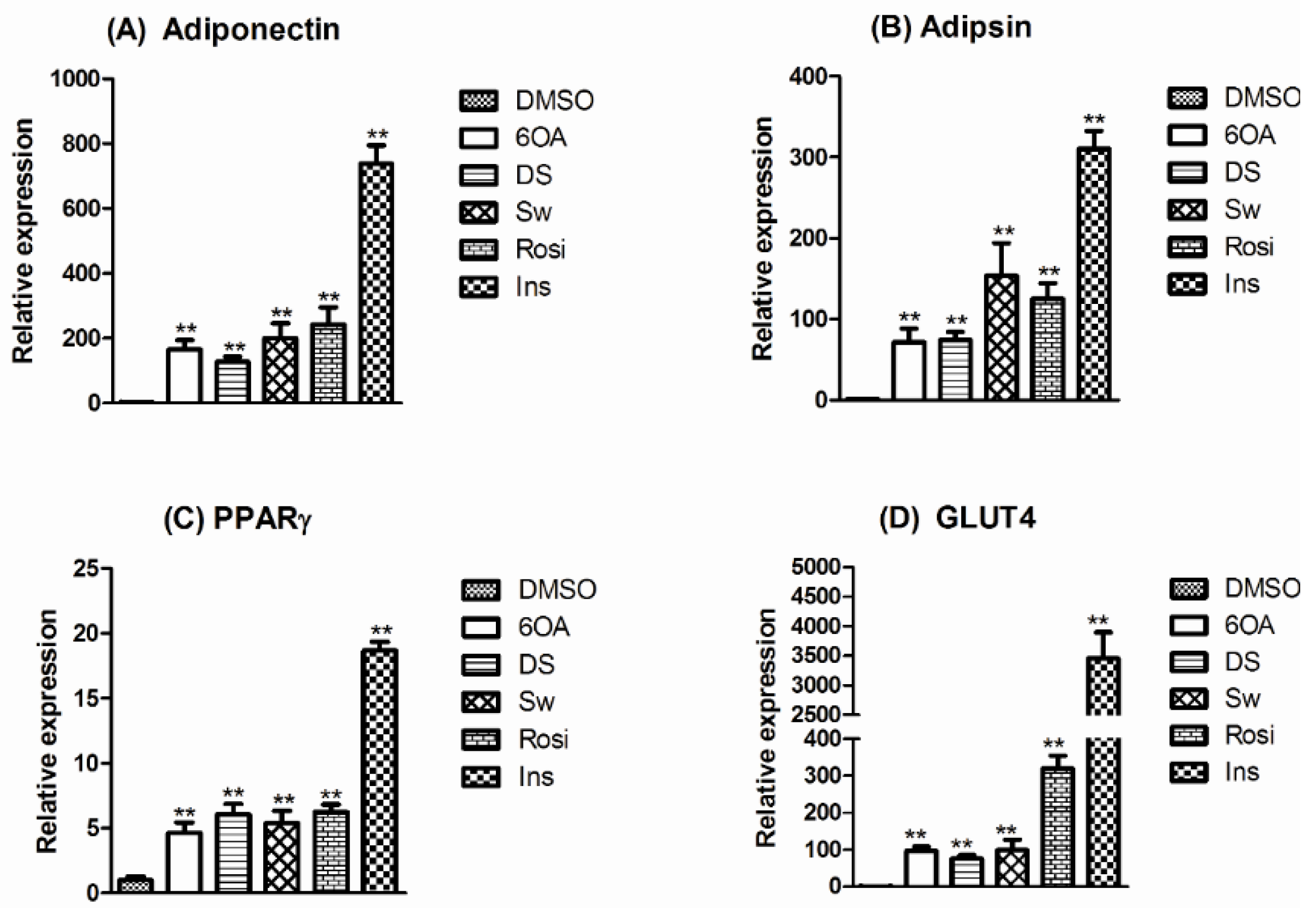
2.4. mRNA Expression of PPARγ and Selected Adipogenic Markers in Mature Differentiated 3T3-L1 Adipocytes

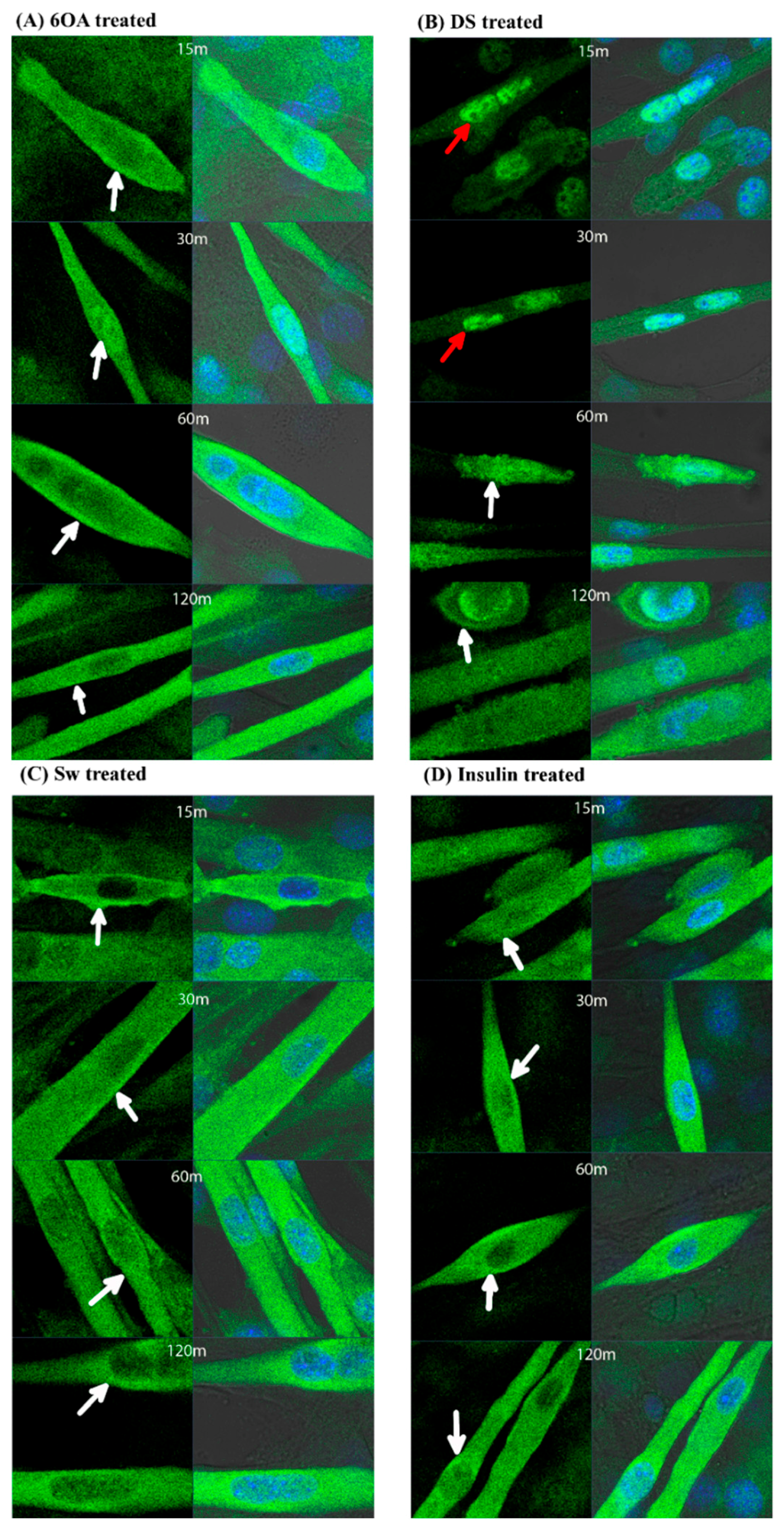
2.5. GLUT4 Translocation in C2C12 Muscle Cells Treated with 6OA, DS and Sw


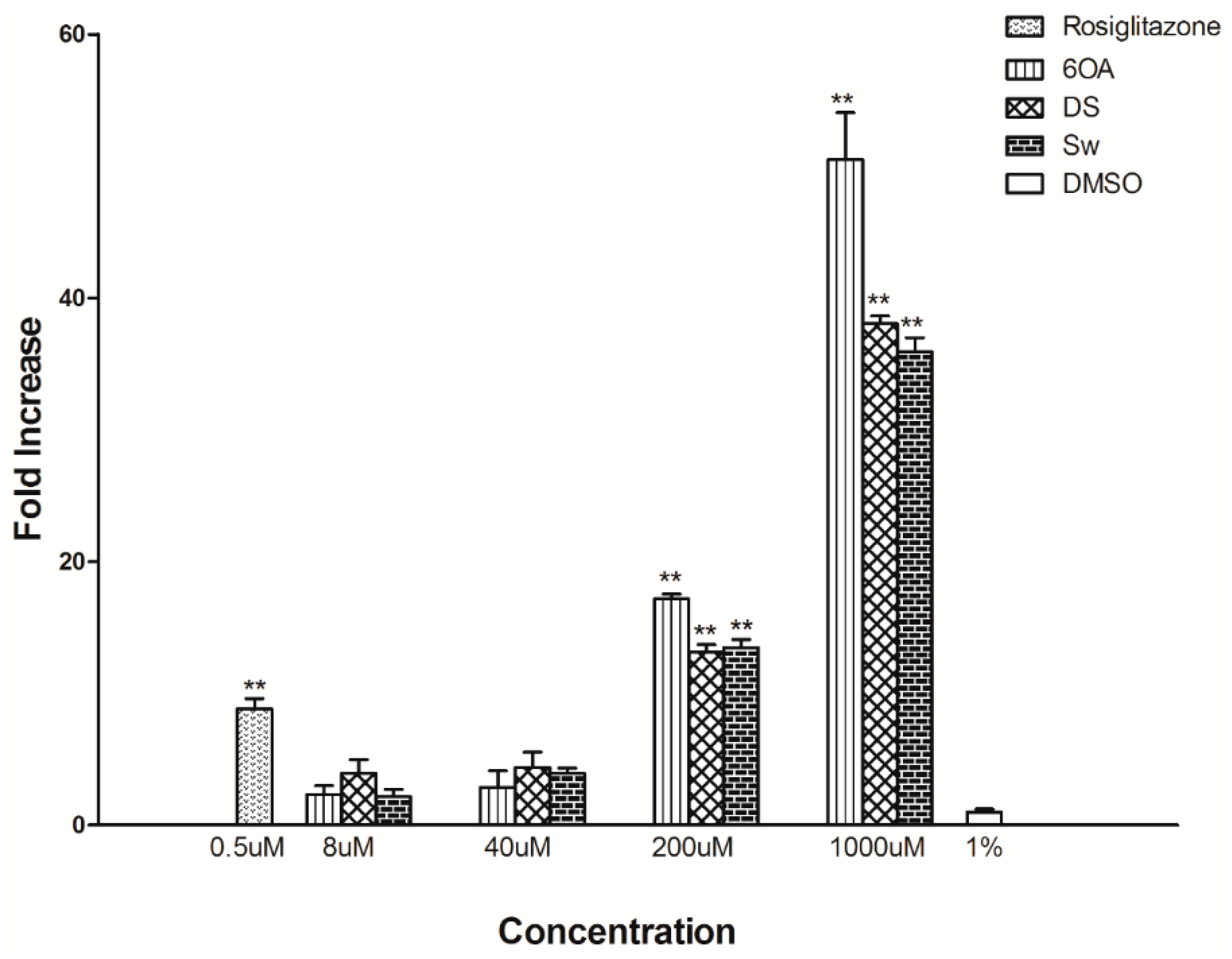
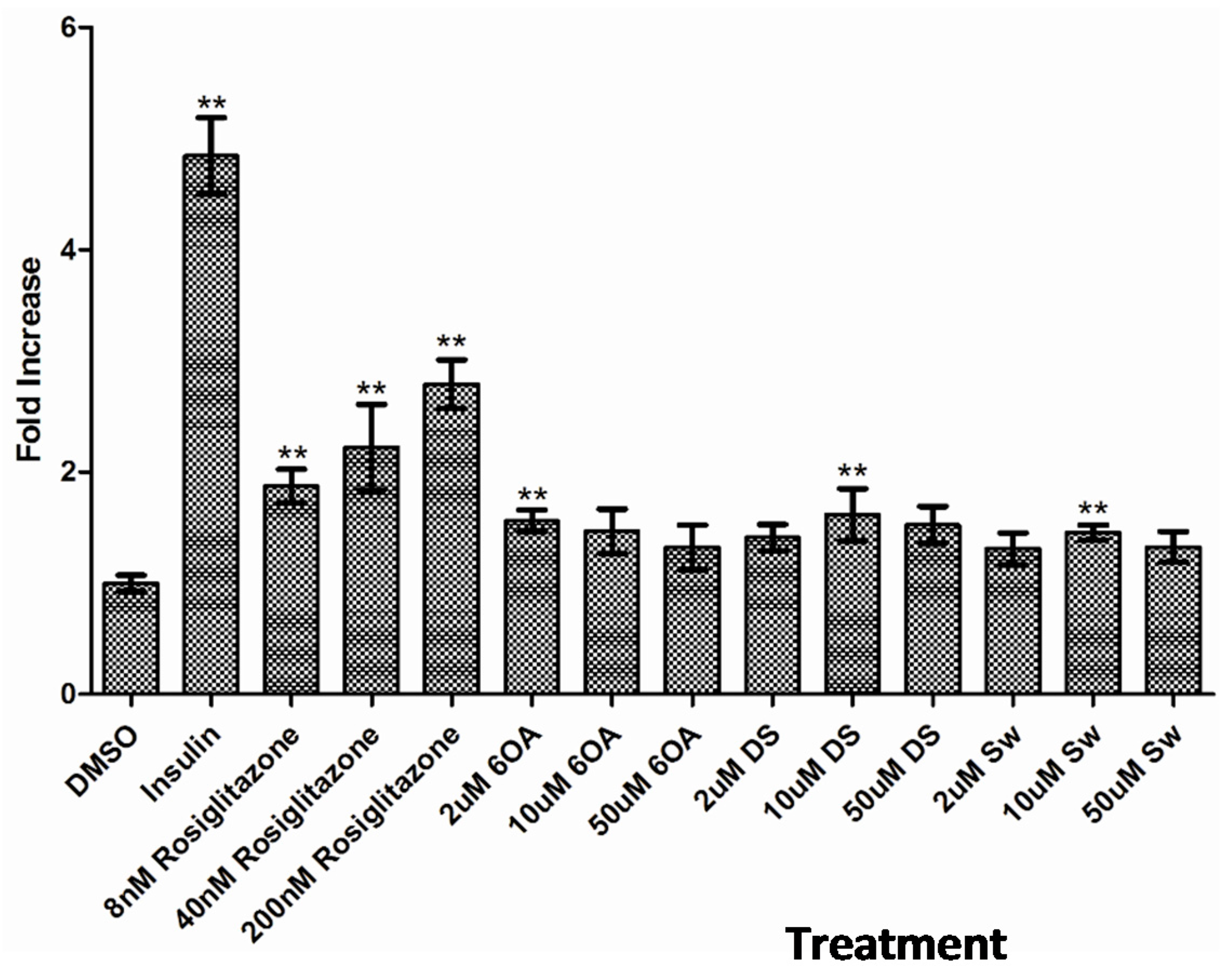
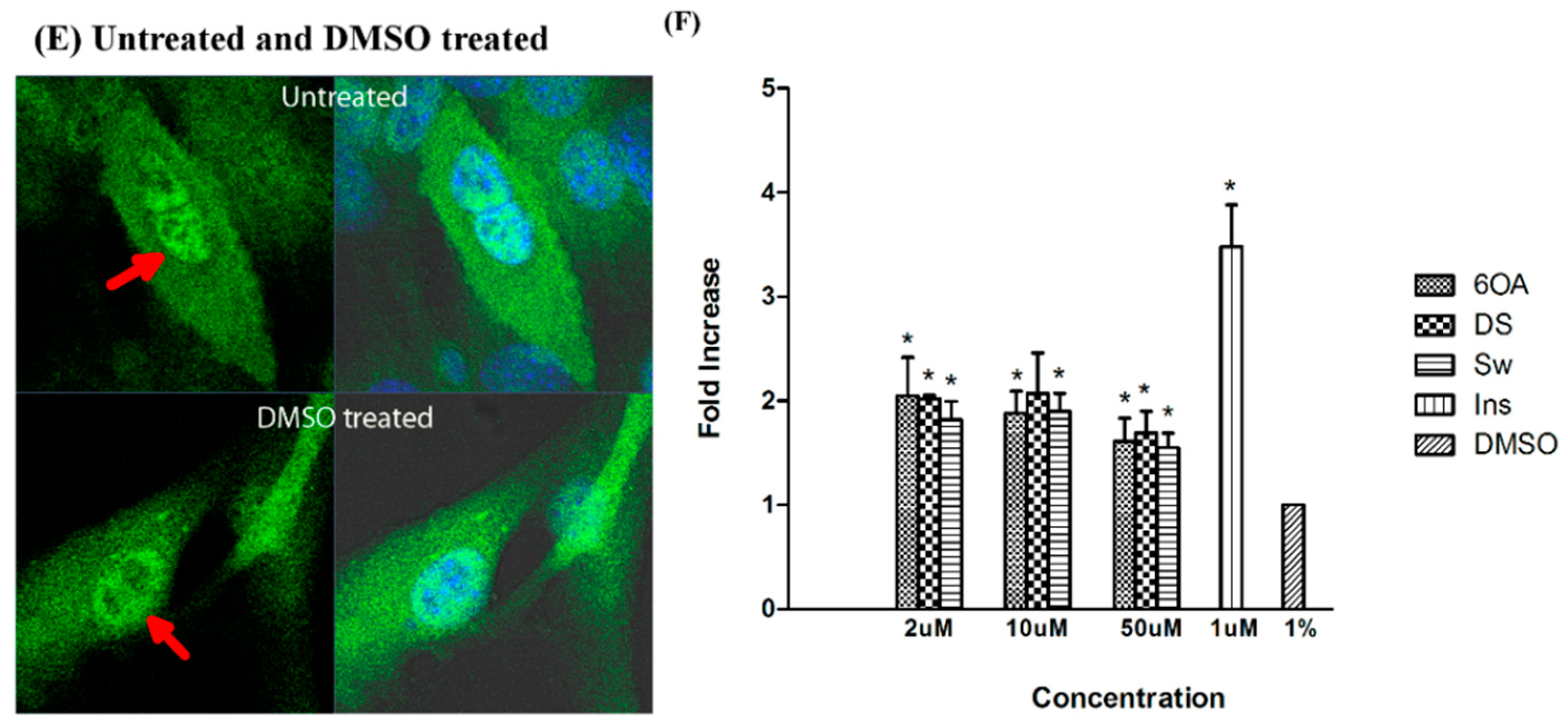
2.6. Glucose Uptake Assay in C2C12 Muscle Cells
2.7. Discussion
3. Experimental Section
3.1. Acquisition of Plant and other Chemical Materials
3.2. Construction of Recombinant Plasmids and Expression of Recombinant Proteins
3.3. Preparation of Extract and Isolation of 6OA, DS and Sw from S. Macrophylla Seeds
3.4. ELISA-Based Binding Assay
3.5. Cell Culture and Differentiation
3.6. Adipocytes Differentiation Assay and Gene Expression

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Sequences (5’-3’) |
|---|---|---|
| β-actin | Forward | TATCGCTGCGCTGGTCGTCG |
| Reverse | ACAGCACAGCCTGGATGGCT | |
| Adiponectin | Forward | TGTTGGAATGACAGGAGCTG |
| Reverse | TGCTGCCGTCATAATGATTC | |
| Adipsin | Forward | ATGACGACTCTGTGCAGGTG |
| Reverse | ATTGCAAGGGTAGGGGTCTC | |
| GLUT4 | Forward | ATCCGGAACCTGGAGGGGCC |
| Reverse | CGGCCAGGCCCAACAGATGG | |
| PPARγ | Forward | GCCTGCGGAAGCCCTTTGGT |
| Reverse | AAGCCTGGGCGGTCTCCACT |
3.7. GLUT4 Translocation Assay
3.8. Glucose Uptake Assay
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dharmalingam, K.; Tan, B.-K.; Mahmud, M.Z.; Sedek, S.A.M.; Majid, M.I.A.; Kuah, M.-K.; Sulaiman, S.F.; Ooi, K.L.; Khan, N.A.K.; Muhammad, T.S.T.; et al. Swietenia macrophylla extract promotes the ability of caenorhabditis elegans to survive pseudomonas aeruginosa infection. J. Ethnopharmacol. 2012, 139, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Goh, B.H.; Chan, C.K.; Kamarudin, M.N.; Kadir, H.A. Swietenia macrophylla king induces mitochondrial-mediated apoptosis through p53 upregulation in hct116 colorectal carcinoma cells. J. Ethnopharmacol. 2014, 153, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Moghadamtousi, S.Z.; Goh, B.H.; Chan, C.K.; Shabab, T.; Kadir, H.A. Biological activities and phytochemicals of swietenia macrophylla king. Molecules 2013, 18, 10465–10483. [Google Scholar] [CrossRef] [PubMed]
- Arumugasamy, K.; Latha, K.V.; Kumar, N.H. Studies on some pharmacognostic profiles of swietenia macophylla. King. Anc. Sci. Life 2004, 24, 97–102. [Google Scholar] [PubMed]
- Dewanjee, S.; Maiti, A.; Das, A.K.; Mandal, S.C.; Dey, S.P. Swietenine: A potential oral hypoglycemic from swietenia macrophylla seed. Fitoterapia 2009, 80, 249–251. [Google Scholar] [CrossRef] [PubMed]
- Kalaivanan, K.; Pugalendi, K.V. Antihyperglycemic effect of the alcoholic seed extract of swietenia macrophylla on streptozotocin-diabetic rats. Pharmacognosy Res. 2011, 3, 67–71. [Google Scholar] [PubMed]
- Dimitriadis, G.; Mitrou, P.; Lambadiari, V.; Maratou, E.; Raptis, S.A. Insulin effects in muscle and adipose tissue. Diabetes Res. Clin. Pract. 2011, 93 (Suppl. 1), S52–S59. [Google Scholar] [CrossRef]
- Armoni, M.; Kritz, N.; Harel, C.; Bar-Yoseph, F.; Chen, H.; Quon, M.J.; Karnieli, E. Peroxisome proliferator-activated receptor-gamma represses GLUT4 promoter activity in primary adipocytes, and rosiglitazone alleviates this effect. J. Biol. Chem. 2003, 278, 30614–30623. [Google Scholar] [CrossRef] [PubMed]
- Willson, T.M.; Lambert, M.H.; Kliewer, S.A. Peroxisome proliferator-activated receptor gamma and metabolic disease. Annu. Rev. Biochem. 2001, 70, 341–367. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.A. Rosiglitazone: A new therapy for type 2 diabetes. Expert Opin. Investig. Drugs 1999, 8, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Boden, G.; Zhang, M. Recent findings concerning thiazolidinediones in the treatment of diabetes. Expert Opin. Investig. Drugs 2006, 15, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Komajda, M.; McMurray, J.J.V.; Beck-Nielsen, H.; Gomis, R.; Hanefeld, M.; Pocock, S.J.; Curtis, P.S.; Jones, N.P.; Home, P.D. Heart failure events with rosiglitazone in type 2 diabetes: Data from the record clinical trial. Eur. Heart J. 2010, 31, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Yokoi, H.; Mizukami, H.; Nagatsu, A.; Ohno, T.; Tanabe, H.; Inoue, M. Peroxisome proliferator-activated receptor gamma ligands isolated from adlay seed (Coix lacryma-jobi L. var. ma-yuen STAPF.). Biol. Pharm. Bull. 2009, 32, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.J.; Day, C. Traditional plant medicines as treatments for diabetes. Diabetes Care 1989, 12, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, J.K.; Cho, M.C.; Shin, S.; Yoon, D.Y.; Heo, Y.S.; Kim, Y. Cytotoxic flavonoids as agonists of peroxisome proliferator-activated receptor gamma on human cervical and prostate cancer cells. J. Nat. Prod. 2010, 73, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Kadota, S.; Marpang, L.; Kikuchi, T.; Ekimoto, H. Constituents of the Seeds of Swietenia mahogani JACQ. I. Isolation, Structures, and 1H- and 13C- Nuclear Magnetic Resonance Signal Assignments of New Tetranortriterpenoids Related to Swietenine and Swietenolide. Chem. Pharm. Bull. 1990, 38, 639–651. [Google Scholar] [CrossRef]
- Goh, B.H.; Abdul Kadir, H.; Abdul Malek, S.N.; Ng, S.W. Swietenolide diacetate from the seeds of swietenia macrophylla. Acta Crystallogr. Sect. E Struct. Rep. Online 2010, 66, o1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, B.H.; Abdul Kadir, H.; Abdul Malek, S.N.; Ng, S.W. (αR,4R,4aR,6aS,7R,8S,10R,11S)-Methyl-α-acet-oxy-4-(3-furan-yl)-10-hy-droxy-4a,7,9,9-tetra-methyl-2,13-dioxo-1,4,4a,5,6,6a,7,8,9,10,11,12-dodeca-hydro-7,11-methano-2H-cyclo-octa-[f][2]benzopyran-8-acetate (6-O-acetyl-swietenolide) from the seeds of Swietenia macrophylla. Acta Crystallogr. Sect. E Struct. Rep. Online 2010, 66, o2802–o2803. [Google Scholar] [PubMed]
- Eriksson, J.; Franssila-Kallunki, A.; Ekstrand, A.; Saloranta, C.; Widen, E.; Schalin, C.; Groop, L. Early metabolic defects in persons at increased risk for non-insulin-dependent diabetes mellitus. N. Engl. J. Med. 1989, 321, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.I. Cellular mechanisms of insulin resistance in humans. Am. J. Cardiol 1999, 84, 3J–10J. [Google Scholar] [CrossRef]
- Spiegelman, B.M. Peroxisome proliferator-activated receptor gamma: A key regulator of adipogenesis and systemic insulin sensitivity. Eur. J. Med. Res. 1997, 2, 457–464. [Google Scholar] [PubMed]
- Ciudin, A.; Hernandez, C.; Simo, R. Update on cardiovascular safety of ppargamma agonists and relevance to medicinal chemistry and clinical pharmacology. Curr. Top. Med. Chem. 2012, 12, 585–604. [Google Scholar] [CrossRef] [PubMed]
- Albrektsen, T.; Frederiksen, K.S.; Holmes, W.E.; Boel, E.; Taylor, K.; Fleckner, J. Novel genes regulated by the insulin sensitizer rosiglitazone during adipocyte differentiation. Diabetes 2002, 51, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Soukas, A.; Socci, N.D.; Saatkamp, B.D.; Novelli, S.; Friedman, J.M. Distinct transcriptional profiles of adipogenesis in vivo and in vitro. J. Biol. Chem. 2001, 276, 34167–34174. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Takahashi, M.; Funahashi, T.; Kihara, S.; Nishizawa, H.; Kishida, K.; Nagaretani, H.; Matsuda, M.; Komuro, R.; Ouchi, N.; et al. Ppargamma ligands increase expression and plasma concentrations of adiponectin, an adipose-derived protein. Diabetes 2001, 50, 2094–2099. [Google Scholar] [CrossRef] [PubMed]
- Moustaid, N.; Lasnier, F.; Hainque, B.; Quignard-Boulange, A.; Pairault, J. Analysis of gene expression during adipogenesis in 3T3-F442A preadipocytes: Insulin and dexamethasone control. J. Cell Biochem. 1990, 42, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Pessin, J.E.; Bell, G.I. Mammalian facilitative glucose transporter family: Structure and molecular regulation. Annu. Rev. Physiol. 1992, 54, 911–930. [Google Scholar] [CrossRef] [PubMed]
- Smith, U. Impaired (“diabetic”) insulin signaling and action occur in fat cells long before glucose intolerance—Is insulin resistance initiated in the adipose tissue? Int. J. Obes. Relat. Metab. Disord. 2002, 26, 897–904. [Google Scholar] [PubMed]
- Ren, J.M.; Marshall, B.A.; Mueckler, M.M.; McCaleb, M.; Amatruda, J.M.; Shulman, G.I. Overexpression of GLUT4 protein in muscle increases basal and insulin-stimulated whole body glucose disposal in conscious mice. J. Clin. Investig. 1995, 95, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Funaki, M.; Randhawa, P.; Janmey, P.A. Separation of insulin signaling into distinct GLUT4 translocation and activation steps. Mol. Cell Biol. 2004, 24, 7567–7577. [Google Scholar] [CrossRef] [PubMed]
- Han, J.H.; Zhou, W.; Li, W.; Tuan, P.Q.; Khoi, N.M.; Thuong, P.T.; Na, M.; Myung, C.S. Pentacyclic triterpenoids from Astilbe rivularis that enhance glucose uptake via the activation of Akt and Erk1/2 in C2C12 myotubes. J. Nat. Prod. 2015, 78, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Han, J.H.; Oh, J.; Kulkarni, R.; Zhou, W.; Ferreira, D.; Jang, T.S.; Myung, C.S.; Na, M. Steroidal alkaloids from veratrum nigrum enhance glucose uptake in skeletal muscle cells. J. Nat. Prod. 2015, 78, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xie, Y.; Morrison, R.F.; Bucher, N.L.; Farmer, S.R. PPARγ induces the insulin-dependent glucose transporter GLUT4 in the absence of C/EBPα during the conversion of 3T3 fibroblasts into adipocytes. J. Clin. Investig. 1998, 101, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Leney, S.E.; Tavare, J.M. The molecular basis of insulin-stimulated glucose uptake: Signalling, trafficking and potential drug targets. J. Endocrinol. 2009, 203, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M.; Flier, J.S. Adipogenesis and obesity: Rounding out the big picture. Cell 1996, 87, 377–389. [Google Scholar] [CrossRef]
- Solomon, K.A.; Malathi, R.; Rajan, S.S.; Narasimhan, S.; Nethaji, M. Swietenine. Acta Crystallogr. 2003, E59, o1519–o1521. [Google Scholar] [CrossRef]
- Johnson, B.H.; Hecht, M.H. Recombinant proteins can be isolated from E. coli cells by repeated cycles of freezing and thawing. Biotechnology 1994, 12, 1357–1360. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lau, W.K.; Goh, B.H.; Kadir, H.A.; Shu-Chien, A.C.; Tengku Muhammad, T.S. Potent PPARγ Ligands from Swietenia macrophylla Are Capable of Stimulating Glucose Uptake in Muscle Cells. Molecules 2015, 20, 22301-22314. https://doi.org/10.3390/molecules201219847
Lau WK, Goh BH, Kadir HA, Shu-Chien AC, Tengku Muhammad TS. Potent PPARγ Ligands from Swietenia macrophylla Are Capable of Stimulating Glucose Uptake in Muscle Cells. Molecules. 2015; 20(12):22301-22314. https://doi.org/10.3390/molecules201219847
Chicago/Turabian StyleLau, Wai Kwan, Bey Hing Goh, Habsah Abdul Kadir, Alexander Chong Shu-Chien, and Tengku Sifzizul Tengku Muhammad. 2015. "Potent PPARγ Ligands from Swietenia macrophylla Are Capable of Stimulating Glucose Uptake in Muscle Cells" Molecules 20, no. 12: 22301-22314. https://doi.org/10.3390/molecules201219847