Effects of Yerba maté, a Plant Extract Formulation (“YGD”) and Resveratrol in 3T3-L1 Adipogenesis


Abstract
:1. Introduction
2. Results and Discussion




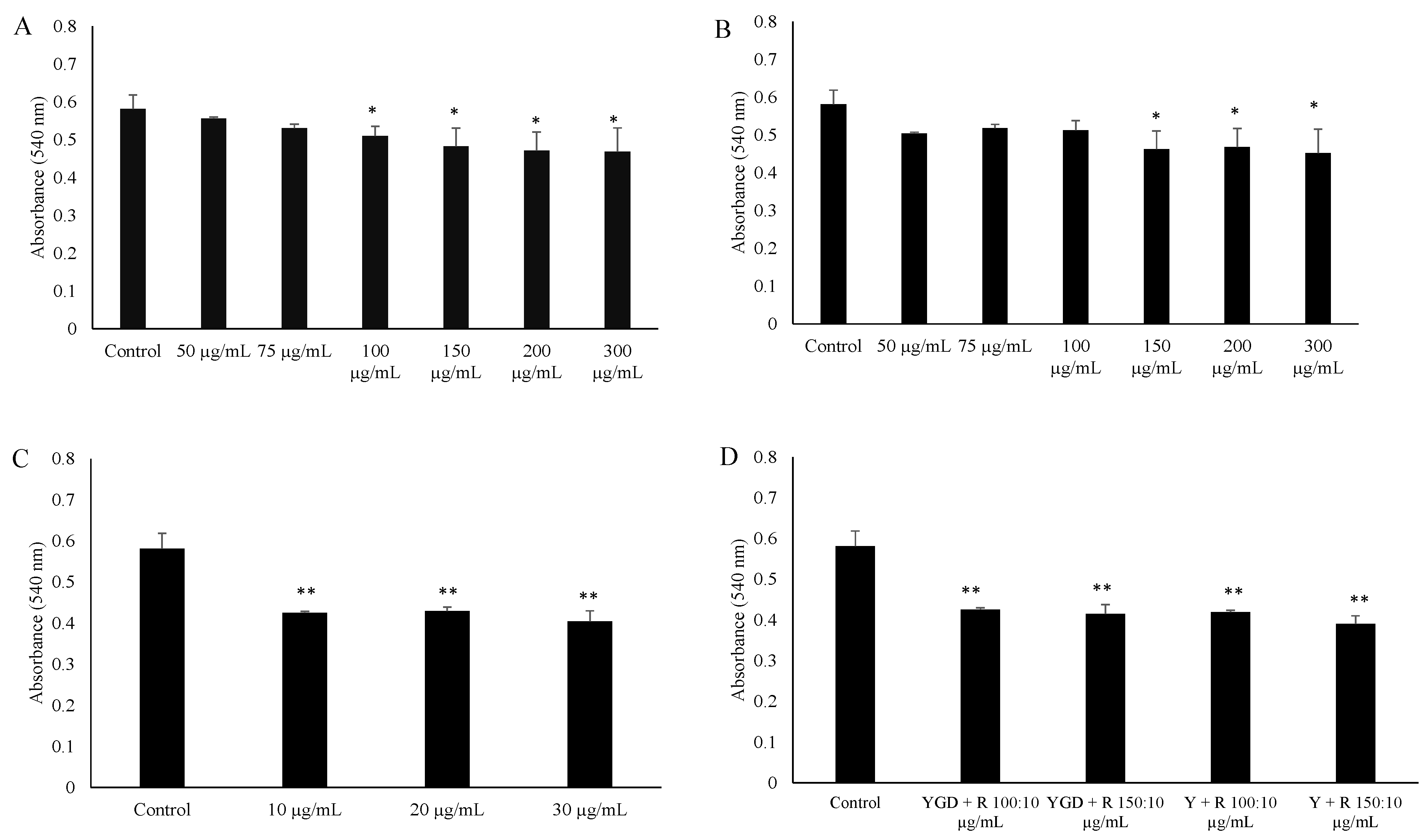{kind=link}
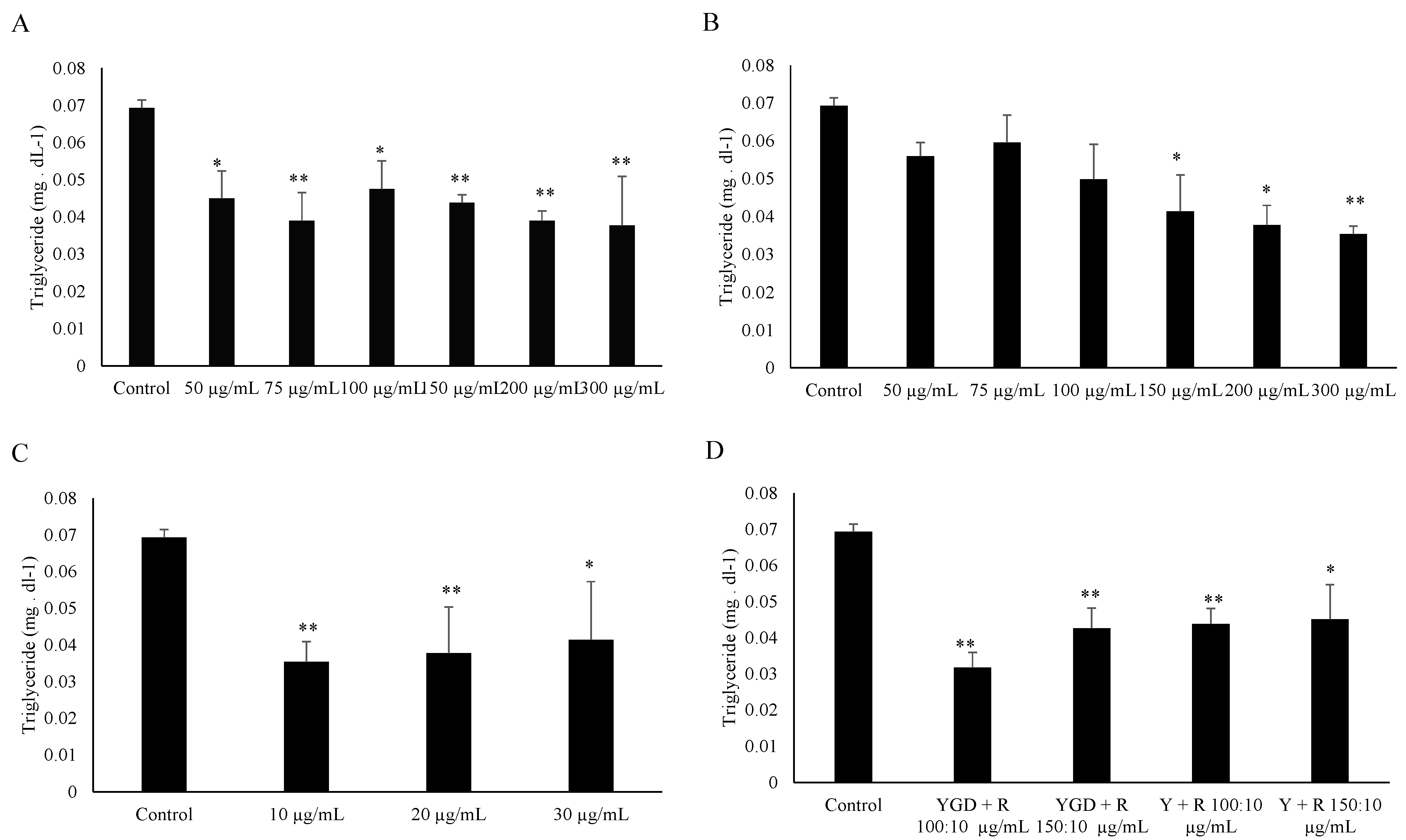{kind=link}
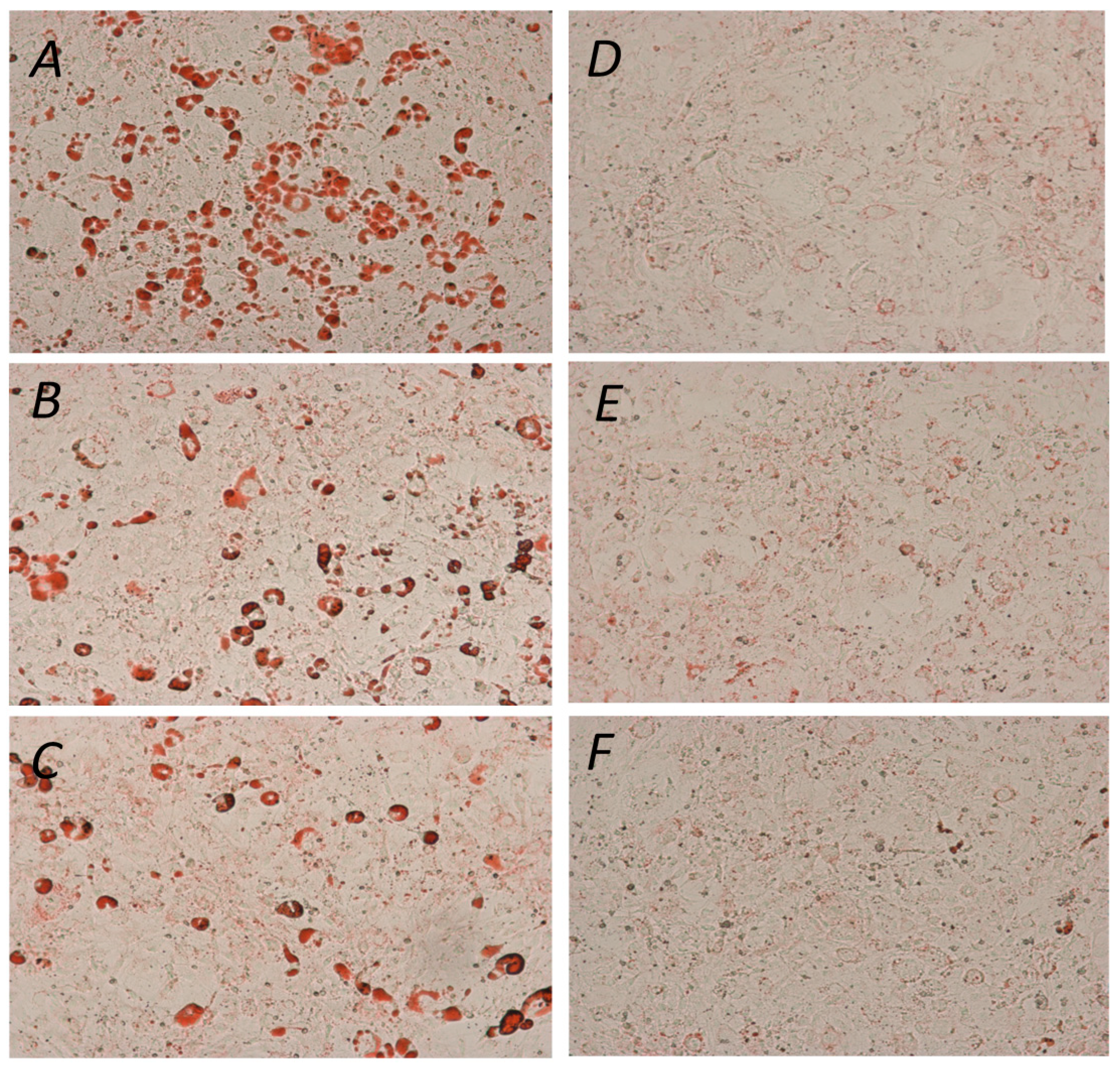{kind=link}
| Functional Gene Group | Gene | Yerba maté 150 μg/mL | YGD 100 μg/mL | Resveratrol 10 μg/mL | YGD 100 + R 10 μg/mL | Y 150 + R 10 μg/mL |
|---|---|---|---|---|---|---|
| Pro-adipogenic | Adig | −2.08 ± 0.31 * | −2.46 ± 0.93 * | −2.81 ± 0.32 * | −2.04 ± 0.47 * | −2.43 ± 0.59 * |
| Axin | −2.20 ± 0.83 * | −2.80 ± 0.86 * | −1.58 ± 0.11 | −2.99 ± 0.68 * | −2.60 ± 0.51 * | |
| Bmp2 | −1.95 ± 0.54 | −4.20 ± 1.58 * | −2.00 ± 0.11 * | −3.65 ± 0.94 * | −2.01 ± 0.93 * | |
| Cebpa | −4.13 ± 0.63 * | −7.20 ± 2.11 * | −2.29 ± 0.31 * | −5.58 ± 1.58 * | −4.41 ± 1.48 * | |
| Cebpb | −1.04 ± 0.14 | −1.24 ± 0.+23 | −2.28 ±0.88 * | −2.69 ± 0.69 * | −2.61 ± 0.62 * | |
| Ccnd1 | −1.92 ± 0.58 | −2.17 ± 0.83 * | −2.65 ± 0.65 * | −-2.00 ± 0.65 * | −2.06 ± 0.65 * | |
| Cdk4 | 1.12 ± 0.44 | 1.81 ± 0.28 | −2.39 ± 0.65 * | −2.88 ± 0.69 * | −3.74 ± 0.77 * | |
| Fasn | −1.97 ± 0.18 | −2.53 ± 0.64 * | −2.31 ±0.48 * | −2.52 ± 0.82 * | −2.22 ± 0.12 * | |
| Fgf2 | −1.41 ± 0.09 | −1.34 ± 0.49 | −2.36 ±0.81 * | −2.45 ± 0.51 * | −3.25 ± 0.31 * | |
| Fgf10 | −3.19 ± 0.69 * | −3.59 ± 1.64 * | −4.89 ± 1.94 * | −2.74 ± 0.34 * | −3.55 ± 0.84 * | |
| Klf15 | −1.25 ± 0.21 | −1.88 ± 0.22 | −3.83 ± 0.97 * | −4.55 ± 0.94 * | −4.77 ± 0.84 * | |
| Lep | −3.32 ± 0.45 * | −4.52 ± 1.21 * | −2.07 ± 0.41 * | −2.21 ± 0.61 * | −2.38 ± 0.96 * | |
| Lpl | −2.94 ± 0.69 * | −6.79 ± 1.65 * | −7.03 ± 0.91 * | −6.21 ± 2.01 * | −4.09 ± 1.19 * | |
| Pparg | −2.02 ± 0.22 * | −3.08 ±1.05 * | −2.41 ± 0.89 * | −2.69 ± 0.88 * | −2.24 ± 0.51 * | |
| Srebf1 | −1.87 ± 0.35 | −2.97 ± 1.18 * | -1.06 ± 0.58 | −2.32 ± 0.76 * | −2.88 ± 0.83* | |
| Anti-adipogenic | Ddit3 | 1.00 ± 0.14 | 1.47 ± 0.72 | 2.47 ± 0.57 * | 2.51 ± 0.40 * | 2.73 ± 0.52 * |
| Dlk1 | 4.82 ± 1.42 * | 1.01 ± 0.23 | 1.44 ± 0.81 | 2.98 ± 0.88 * | 3.61 ± 0.89 * | |
| Foxo1 | 1.87 ± 0.50 | 1.78 ± 0.29 | 2.30 ± 0.88 * | 3.54 ± 0.92 * | 3.65 ± 0.91 * | |
| Klf2 | 3.91 ±1.06 * | 4.82 ± 1.14 * | 5.57 ± 1.28 * | 3.65 ± 0.55 * | 4.63 ± 0.73 * | |
| Ucp1 | 3.47 ± 1.59 * | 4.15 ± 1.67 * | 6.29 ± 2.16 * | 6.02 ± 1.99 * | 6.80 ± 2.73 * | |
| Sirt1 | 1.23 ± 0.13 | 1.30 ± 0.50 | 2.94 ± 0.88 * | 3.20 ± 1.02 * | 3.08 ± 0.99 * | |
| Sirt2 | 1.79 ± 0.48 | 1.52 ± 0.22 | 2.08 ± 0.27 * | 2.88 ± 0.92 * | 2.76 ± 0.89 * |
3 Experimental Section
3.1. Materials
3.2. In Vitro Evaluation of Adipogenesis
3.2.1. 3T3-L1 Cell Culture
3.2.2. Cytotoxicity Determination—MTT Assay
3.2.3. 3T3-L1 Differentiation Assay—Oil Red O
3.2.4. Quantification of the Triglyceride Content
3.2.5. RNA Extraction and cDNA Synthesis
3.3. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berg, A.H.; Scherer, P.E. Adipose tissue, inflammation, and cardiovascular disease. Circ. Res. 2005, 96, 939–949. [Google Scholar]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar]
- Waxman, A. Why a global strategy on diet, physical activity and health? World Rev. Nutr. Diet. 2005, 95, 162–166. [Google Scholar]
- Santos, A.P.; Rogero, M.M.; Bastos, D.H. Edible plants, their secondary metabolites and antiobesogenic potential. Recent Pat. Food Nutr. Agric. 2010, 2, 195–212. [Google Scholar]
- Rayalam, S.; Della-Fera, M.A.; Baile, C.A. Phytochemicals and regulation of the adipocyte life cycle. J. Nutr. Biochem. 2008, 19, 717–726. [Google Scholar]
- Pang, J.; Choi, Y.; Park, T. Ilex paraguariensis extract ameliorates obesity induced by high-fat diet: Potential role of ampk in the visceral adipose tissue. Arch. Biochem. Biophys. 2008, 476, 178–185. [Google Scholar]
- Arcari, D.P.; Bartchewsky, W.; dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzoli, J.; de Souza, M.F.; Saad, M.J.; Bastos, D.H.; Gambero, A.; et al. Antiobesity effects of yerba mate extract (Ilex paraguariensis) in high-fat diet-induced obese mice. Obesity 2009, 17, 2127–2133. [Google Scholar]
- Arcari, D.P.; Bartchewsky, W., Jr.; dos Santos, T.W.; Oliveira, K.A.; DeOliveira, C.C.; Gotardo, E.M.; Pedrazzoli, J., Jr.; Gambero, A.; Ferraz, L.F.; Carvalho Pde, O.; et al. Anti-inflammatory effects of yerba mate extract (Ilex paraguariensis) ameliorate insulin resistance in mice with high fat diet-induced obesity. Mol. Cell Endocrinol. 2011, 335, 110–115. [Google Scholar]
- Hussein, G.M.; Matsuda, H.; Nakamura, S.; Hamao, M.; Akiyama, T.; Tamura, K.; Yoshikawa, M. Mate tea (Ilex paraguariensis) promotes satiety and body weight lowering in mice: Involvement of glucagon-like peptide-1. Biol. Pharm. Bull. 2011, 34, 1849–1855. [Google Scholar]
- Kang, Y.R.; Lee, H.Y.; Kim, J.H.; Moon, D.I.; Seo, M.Y.; Park, S.H.; Choi, K.H.; Kim, C.R.; Kim, S.H.; Oh, J.H.; et al. Anti-obesity and anti-diabetic effects of yerba mate (Ilex paraguariensis) in c57bl/6j mice fed a high-fat diet. Lab. Anim. Res. 2012, 28, 23–29. [Google Scholar]
- Pimentel, G.D.; Lira, F.S.; Rosa, J.C.; Caris, A.V.; Pinheiro, F.; Ribeiro, E.B.; Oller do Nascimento, C.M.; Oyama, L.M. Yerba mate extract (ilex paraguariensis) attenuates both central and peripheral inflammatory effects of diet-induced obesity in rats. J. Nutr. Biochem. 2013, 24, 809–818. [Google Scholar]
- Gosmann, G.; Barlette, A.G.; Dhamer, T.; Arcari, D.P.; Santos, J.C.; de Camargo, E.R.; Acedo, S.; Gambero, A.; Gnoatto, S.C.; Ribeiro, M.L. Phenolic compounds from mate (Ilex paraguariensis) inhibit adipogenesis in 3t3-l1 preadipocytes. Plant Foods Hum. Nutr. 2012, 67, 156–161. [Google Scholar]
- Arcari, D.P.; Santos, J.C.; Gambero, A.; Ferraz, L.F.; Ribeiro, M.L. Modulatory effects of yerba mate (Ilex paraguariensis) on the pi3k-akt signaling pathway. Mol. Nutr. Food Res. 2013, 57, 1882–1885. [Google Scholar]
- Arcari, D.P.; Santos, J.C.; Gambero, A.; Ribeiro, M.L. The in vitro and in vivo effects of yerba mate (Ilex paraguariensis) extract on adipogenesis. Food Chem. 2013, 141, 809–815. [Google Scholar]
- Carmo, L.S.; Rogero, M.M.; Cortez, M.; Yamada, M.; Jacob, P.S.; Bastos, D.H.; Borelli, P.; Ambrosio Fock, R. The effects of yerba mate (Ilex paraguariensis) consumption on il-1, il-6, tnf-α and il-10 production by bone marrow cells in wistar rats fed a high-fat die. Int. J. Vitam. Nutr. Res. 2013, 83, 26–35. [Google Scholar]
- Lima Nda, S.; Franco, J.G.; Peixoto-Silva, N.; Maia, L.A.; Kaezer, A.; Felzenszwalb, I.; de Oliveira, E.; de Moura, E.G.; Lisboa, P.C. Ilex paraguariensis (yerba mate) improves endocrine and metabolic disorders in obese rats primed by early weaning. Eur. J. Nutr. 2014, 53, 73–82. [Google Scholar]
- Borges, M.C.; Vinolo, M.A.; Nakajima, K.; de Castro, I.A.; Bastos, D.H.; Borelli, P.; Fock, R.A.; Tirapegui, J.; Curi, R.; Rogero, M.M. The effect of mate tea (Ilex paraguariensis) on metabolic and inflammatory parameters in high-fat diet-fed wistar rats. Int. J. Food Sci. Nutr. 2013, 64, 561–569. [Google Scholar]
- Martins, F.; Suzan, A.J.; Cerutti, S.M.; Arcari, D.P.; Ribeiro, M.L.; Bastos, D.H.; Carvalho Pde, O. Consumption of mate tea (Ilex paraguariensis) decreases the oxidation of unsaturated fatty acids in mouse liver. Br. J. Nutr. 2009, 101, 527–532. [Google Scholar]
- Paganini Stein, F.L.; Schmidt, B.; Furlong, E.B.; Souza-Soares, L.A.; Soares, M.C.; Vaz, M.R.; Muccillo Baisch, A.L. Vascular responses to extractable fractions of Ilex paraguariensis in rats fed standard and high-cholesterol diets. Biol. Res. Nurs. 2005, 7, 146–156. [Google Scholar]
- Pomilio, A.B.; Trajtemberg, S.; Vitale, A.A. High-performance capillary electrophoresis analysis of mate infusions prepared from stems and leaves of Ilex paraguariensis using automated micellar electrokinetic capillary chromatography. Phytochem. Anal. 2002, 13, 235–241. [Google Scholar]
- Martinet, A.; Hostettmann, K.; Schutz, Y. Thermogenic effects of commercially available plant preparations aimed at treating human obesity. Phytomedicine 1999, 6, 231–238. [Google Scholar]
- Resende, P.E.; Verza, S.G.; Kaiser, S.; Gomes, L.F.; Kucharski, L.C.; Ortega, G.G. The activity of mate saponins (Ilex paraguariensis) in intra-abdominal and epididymal fat, and glucose oxidation in male wistar rats. J. Ethnopharmacol. 2012, 144, 735–740. [Google Scholar]
- Andersen, T.; Fogh, J. Weight loss and delayed gastric emptying following a south american herbal preparation in overweight patients. J. Hum. Nutr. Diet. 2001, 14, 243–250. [Google Scholar]
- Harrold, J.A.; Hughes, G.M.; O’Shiel, K.; Quinn, E.; Boyland, E.J.; Williams, N.J.; Halford, J.C. Acute effects of a herb extract formulation and inulin fiber on appetite, energy intake and food choice. Appetite 2013, 62, 84–90. [Google Scholar]
- Szkudelska, K.; Szkudelski, T. Resveratrol, obesity and diabetes. Eur. J. Pharmacol. 2010, 635, 1–8. [Google Scholar]
- Lippi, G.; Franchini, M.; Favaloro, E.J.; Targher, G. Moderate red wine consumption and cardiovascular disease risk: Beyond the “french paradox”. Semin. Thromb. Hemost. 2010, 36, 59–70. [Google Scholar]
- Bradamante, S.; Barenghi, L.; Villa, A. Cardiovascular protective effects of resveratrol. Cardiovasc. Drug Rev. 2004, 22, 169–188. [Google Scholar]
- Athar, M.; Back, J.H.; Tang, X.; Kim, K.H.; Kopelovich, L.; Bickers, D.R.; Kim, A.L. Resveratrol: A review of preclinical studies for human cancer prevention. Toxicol. Appl. Pharmacol. 2007, 224, 274–283. [Google Scholar]
- De la Lastra, C.A.; Villegas, I. Resveratrol as an antioxidant and pro-oxidant agent: Mechanisms and clinical implications. Biochem. Soc. Trans. 2007, 35, 1156–1160. [Google Scholar]
- Zhang, X.H.; Huang, B.; Choi, S.K.; Seo, J.S. Anti-obesity effect of resveratrol-amplified grape skin extracts on 3t3-l1 adipocytes differentiation. Nutr. Res. Pract. 2012, 6, 286–293. [Google Scholar]
- Ahn, J.; Lee, H.; Kim, S.; Ha, T. Resveratrol inhibits tnf-α-induced changes of adipokines in 3t3-l1 adipocytes. Biochem. Biophys. Res. Commun. 2007, 364, 972–977. [Google Scholar]
- Chen, S.; Li, Z.; Li, W.; Shan, Z.; Zhu, W. Resveratrol inhibits cell differentiation in 3t3-l1 adipocytes via activation of ampk. Can. J. Physiol. Pharmacol. 2011, 89, 793–799. [Google Scholar]
- Chen, S.; Xiao, X.; Feng, X.; Li, W.; Zhou, N.; Zheng, L.; Sun, Y.; Zhang, Z.; Zhu, W. Resveratrol induces sirt1-dependent apoptosis in 3t3-l1 preadipocytes by activating ampk and suppressing akt activity and survivin expression. J. Nutr. Biochem. 2012, 23, 1100–1112. [Google Scholar]
- Costa Cdos, S.; Rohden, F.; Hammes, T.O.; Margis, R.; Bortolotto, J.W.; Padoin, A.V.; Mottin, C.C.; Guaragna, R.M. Resveratrol upregulated sirt1, foxo1, and adiponectin and downregulated pparγ1-3 mrna expression in human visceral adipocytes. Obes. Surg. 2011, 21, 356–361. [Google Scholar]
- Kang, L.; Heng, W.; Yuan, A.; Baolin, L.; Fang, H. Resveratrol modulates adipokine expression and improves insulin sensitivity in adipocytes: Relative to inhibition of inflammatory responses. Biochimie 2010, 92, 789–796. [Google Scholar]
- Kang, N.E.; Ha, A.W.; Kim, J.Y.; Kim, W.K. Resveratrol inhibits the protein expression of transcription factors related adipocyte differentiation and the activity of matrix metalloproteinase in mouse fibroblast 3t3-l1 preadipocytes. Nutr. Res. Pract. 2012, 6, 499–504. [Google Scholar]
- Kwon, J.Y.; Seo, S.G.; Yue, S.; Cheng, J.X.; Lee, K.W.; Kim, K.H. An inhibitory effect of resveratrol in the mitotic clonal expansion and insulin signaling pathway in the early phase of adipogenesis. Nutr. Res. 2012, 32, 607–616. [Google Scholar]
- Lasa, A.; Schweiger, M.; Kotzbeck, P.; Churruca, I.; Simon, E.; Zechner, R.; Portillo, M.P. Resveratrol regulates lipolysis via adipose triglyceride lipase. J. Nutr. Biochem. 2012, 23, 379–384. [Google Scholar]
- Rayalam, S.; Yang, J.Y.; Ambati, S.; Della-Fera, M.A.; Baile, C.A. Resveratrol induces apoptosis and inhibits adipogenesis in 3t3-l1 adipocytes. Phytother. Res. 2008, 22, 1367–1371. [Google Scholar]
- Wang, A.; Liu, M.; Liu, X.; Dong, L.Q.; Glickman, R.D.; Slaga, T.J.; Zhou, Z.; Liu, F. Up-regulation of adiponectin by resveratrol: The essential roles of the akt/foxo1 and amp-activated protein kinase signaling pathways and dsba-l. J. Biol. Chem. 2011, 286, 60–66. [Google Scholar]
- Zhang, H.Y.; Du, Z.X.; Meng, X. Resveratrol prevents tnfα-induced suppression of adiponectin expression via pparγ activation in 3t3-l1 adipocytes. Clin. Exp. Med. 2013, 13, 193–199. [Google Scholar]
- Zhu, J.; Yong, W.; Wu, X.; Yu, Y.; Lv, J.; Liu, C.; Mao, X.; Zhu, Y.; Xu, K.; Han, X.; et al. Anti-inflammatory effect of resveratrol on tnf-α-induced mcp-1 expression in adipocytes. Biochem. Biophys. Res. Commun. 2008, 369, 471–477. [Google Scholar]
- Kim, S.; Jin, Y.; Choi, Y.; Park, T. Resveratrol exerts anti-obesity effects via mechanisms involving down-regulation of adipogenic and inflammatory processes in mice. Biochem. Pharmacol. 2011, 81, 1343–1351. [Google Scholar]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van de Weijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar]
- Tome-Carneiro, J.; Gonzalvez, M.; Larrosa, M.; Garcia-Almagro, F.J.; Aviles-Plaza, F.; Parra, S.; Yanez-Gascon, M.J.; Ruiz-Ros, J.A.; Garcia-Conesa, M.T.; Tomas-Barberan, F.A.; et al. Consumption of a grape extract supplement containing resveratrol decreases oxidized ldl and apob in patients undergoing primary prevention of cardiovascular disease: A triple-blind, 6-month follow-up, placebo-controlled, randomized trial. Mol. Nutr. Food Res. 2012, 56, 810–821. [Google Scholar]
- Tome-Carneiro, J.; Gonzalvez, M.; Larrosa, M.; Yanez-Gascon, M.J.; Garcia-Almagro, F.J.; Ruiz-Ros, J.A.; Tomas-Barberan, F.A.; Garcia-Conesa, M.T.; Espin, J.C. Grape resveratrol increases serum adiponectin and downregulates inflammatory genes in peripheral blood mononuclear cells: A triple-blind, placebo-controlled, one-year clinical trial in patients with stable coronary artery disease. Cardiovasc. Drugs Ther. 2013, 27, 37–48. [Google Scholar]
- Rayalam, S.; Della-Fera, M.A.; Yang, J.Y.; Park, H.J.; Ambati, S.; Baile, C.A. Resveratrol potentiates genistein’s antiadipogenic and proapoptotic effects in 3t3-l1 adipocytes. J. Nutr. 2007, 137, 2668–2673. [Google Scholar]
- Brown, M.S.; Goldstein, J.L. The srebp pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell 1997, 89, 331–340. [Google Scholar]
- Kim, J.B.; Sarraf, P.; Wright, M.; Yao, K.M.; Mueller, E.; Solanes, G.; Lowell, B.B.; Spiegelman, B.M. Nutritional and insulin regulation of fatty acid synthetase and leptin gene expression through add1/srebp1. J. Clin. Investig. 1998, 101, 1–9. [Google Scholar]
- Lasa, A.; Churruca, I.; Eseberri, I.; Andres-Lacueva, C.; Portillo, M.P. Delipidating effect of resveratrol metabolites in 3t3-l1 adipocytes. Mol. Nutr. Food Res. 2012, 56, 1559–1568. [Google Scholar]
- Banerjee, S.S.; Feinberg, M.W.; Watanabe, M.; Gray, S.; Haspel, R.L.; Denkinger, D.J.; Kawahara, R.; Hauner, H.; Jain, M.K. The kruppel-like factor klf2 inhibits peroxisome proliferator-activated receptor-γ expression and adipogenesis. J. Biol. Chem. 2003, 278, 2581–2584. [Google Scholar]
- Mori, T.; Sakaue, H.; Iguchi, H.; Gomi, H.; Okada, Y.; Takashima, Y.; Nakamura, K.; Nakamura, T.; Yamauchi, T.; Kubota, N.; et al. Role of kruppel-like factor 15 (klf15) in transcriptional regulation of adipogenesis. J. Biol. Chem. 2005, 280, 12867–12875. [Google Scholar]
- Wang, Y.; Kim, K.A.; Kim, J.H.; Sul, H.S. Pref-1, a preadipocyte secreted factor that inhibits adipogenesis. J. Nutr. 2006, 136, 2953–2956. [Google Scholar]
- Kopan, R.; Ilagan, M.X. The canonical notch signaling pathway: Unfolding the activation mechanism. Cell 2009, 137, 216–233. [Google Scholar]
- Chang, H.; Brown, C.W.; Matzuk, M.M. Genetic analysis of the mammalian transforming growth factor-beta superfamily. Endocr. Rev. 2002, 23, 787–823. [Google Scholar]
- Sottile, V.; Seuwen, K. Bone morphogenetic protein-2 stimulates adipogenic differentiation of mesenchymal precursor cells in synergy with brl 49653 (rosiglitazone). FEBS Lett. 2000, 475, 201–204. [Google Scholar]
- Dowell, P.; Otto, T.C.; Adi, S.; Lane, M.D. Convergence of peroxisome proliferator-activated receptor γ and foxo1 signaling pathways. J. Biol. Chem. 2003, 278, 45485–45491. [Google Scholar]
- Armoni, M.; Harel, C.; Karni, S.; Chen, H.; Bar-Yoseph, F.; Ver, M.R.; Quon, M.J.; Karnieli, E. Foxo1 represses peroxisome proliferator-activated receptor-γ1 and -γ2 gene promoters in primary adipocytes. A novel paradigm to increase insulin sensitivity. J. Biol. Chem. 2006, 281, 19881–19891. [Google Scholar]
- Jing, E.; Gesta, S.; Kahn, C.R. Sirt2 regulates adipocyte differentiation through foxo1 acetylation/deacetylation. Cell Metab. 2007, 6, 105–114. [Google Scholar]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; Machado de Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing ppar-gamma. Nature 2004, 429, 771–776. [Google Scholar]
- Wang, F.; Tong, Q. Sirt2 suppresses adipocyte differentiation by deacetylating foxo1 and enhancing foxo1's repressive interaction with ppargamma. Mol. Biol. Cell 2009, 20, 801–808. [Google Scholar]
- Pacholec, M.; Bleasdale, J.E.; Chrunyk, B.; Cunningham, D.; Flynn, D.; Garofalo, R.S.; Griffith, D.; Griffor, M.; Loulakis, P.; Pabst, B.; et al. Srt1720, srt2183, srt1460, and resveratrol are not direct activators of sirtl. J. Biol. Chem. 2010, 285, 8340–8351. [Google Scholar]
- Tang, Q.Q.; Lane, M.D. Role of c/ebp homologous protein (chop-10) in the programmed activation of ccaat/enhancer-binding protein-beta during adipogenesis. Proc. Natl. Acad. Sci. USA 2000, 97, 12446–12450. [Google Scholar]
- Huang, H.; Lane, M.D.; Tang, Q.Q. Effect of serum on the down-regulation of chop-10 during differentiation of 3t3-l1 preadipocytes. Biochem. Biophys. Res. Commun. 2005, 338, 1185–1188. [Google Scholar]
- Sakaue, H.; Konishi, M.; Ogawa, W.; Asaki, T.; Mori, T.; Yamasaki, M.; Takata, M.; Ueno, H.; Kato, S.; Kasuga, M.; et al. Requirement of fibroblast growth factor 10 in development of white adipose tissue. Genes Dev. 2002, 16, 908–912. [Google Scholar]
- Abella, A.; Dubus, P.; Malumbres, M.; Rane, S.G.; Kiyokawa, H.; Sicard, A.; Vignon, F.; Langin, D.; Barbacid, M.; Fajas, L. Cdk4 promotes adipogenesis through ppargamma activation. Cell Metab. 2005, 2, 239–249. [Google Scholar]
- Bennett, C.N.; Ross, S.E.; Longo, K.A.; Bajnok, L.; Hemati, N.; Johnson, K.W.; Harrison, S.D.; MacDougald, O.A. Regulation of wnt signaling during adipogenesis. J. Biol. Chem. 2002, 277, 30998–31004. [Google Scholar]
- Peifer, M.; Polakis, P. Wnt signaling in oncogenesis and embryogenesis—A look outside the nucleus. Science 2000, 287, 1606–1609. [Google Scholar]
- Fu, M.; Rao, M.; Bouras, T.; Wang, C.; Wu, K.; Zhang, X.; Li, Z.; Yao, T.P.; Pestell, R.G. Cyclin d1 inhibits peroxisome proliferator-activated receptor γ-mediated adipogenesis through histone deacetylase recruitment. J. Biol. Chem. 2005, 280, 16934–16941. [Google Scholar]
- Mitterberger, M.C.; Zwerschke, W. Mechanisms of resveratrol-induced inhibition of clonal expansion and terminal adipogenic differentiation in 3t3-l1 preadipocytes. J. Gerontol. Ser. A 2013, 68, 1356–1376. [Google Scholar]
- Andrade, J.M.; Frade, A.C.; Guimaraes, J.B.; Freitas, K.M.; Lopes, M.T.; Guimaraes, A.L.; de Paula, A.M.; Coimbra, C.C.; Santos, S.H. Resveratrol increases brown adipose tissue thermogenesis markers by increasing sirt1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur. J. Nutr. 2014, 53, 1503–1510. [Google Scholar]
- Moon, H.S.; Chung, C.S.; Lee, H.G.; Kim, T.G.; Choi, Y.J.; Cho, C.S. Inhibitory effect of (−)-epigallocatechin-3-gallate on lipid accumulation of 3t3-l1 cells. Obesity 2007, 15, 2571–2582. [Google Scholar]
- Sample Availability: Samples of all the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, J.C.; Gotardo, É.M.F.; Brianti, M.T.; Piraee, M.; Gambero, A.; Ribeiro, M.L. Effects of Yerba maté, a Plant Extract Formulation (“YGD”) and Resveratrol in 3T3-L1 Adipogenesis. Molecules 2014, 19, 16909-16924. https://doi.org/10.3390/molecules191016909
Santos JC, Gotardo ÉMF, Brianti MT, Piraee M, Gambero A, Ribeiro ML. Effects of Yerba maté, a Plant Extract Formulation (“YGD”) and Resveratrol in 3T3-L1 Adipogenesis. Molecules. 2014; 19(10):16909-16924. https://doi.org/10.3390/molecules191016909
Chicago/Turabian StyleSantos, Juliana C., Érica M. F. Gotardo, Mitsue T. Brianti, Mahmood Piraee, Alessandra Gambero, and Marcelo L. Ribeiro. 2014. "Effects of Yerba maté, a Plant Extract Formulation (“YGD”) and Resveratrol in 3T3-L1 Adipogenesis" Molecules 19, no. 10: 16909-16924. https://doi.org/10.3390/molecules191016909