Cordycepin Induces S Phase Arrest and Apoptosis in Human Gallbladder Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
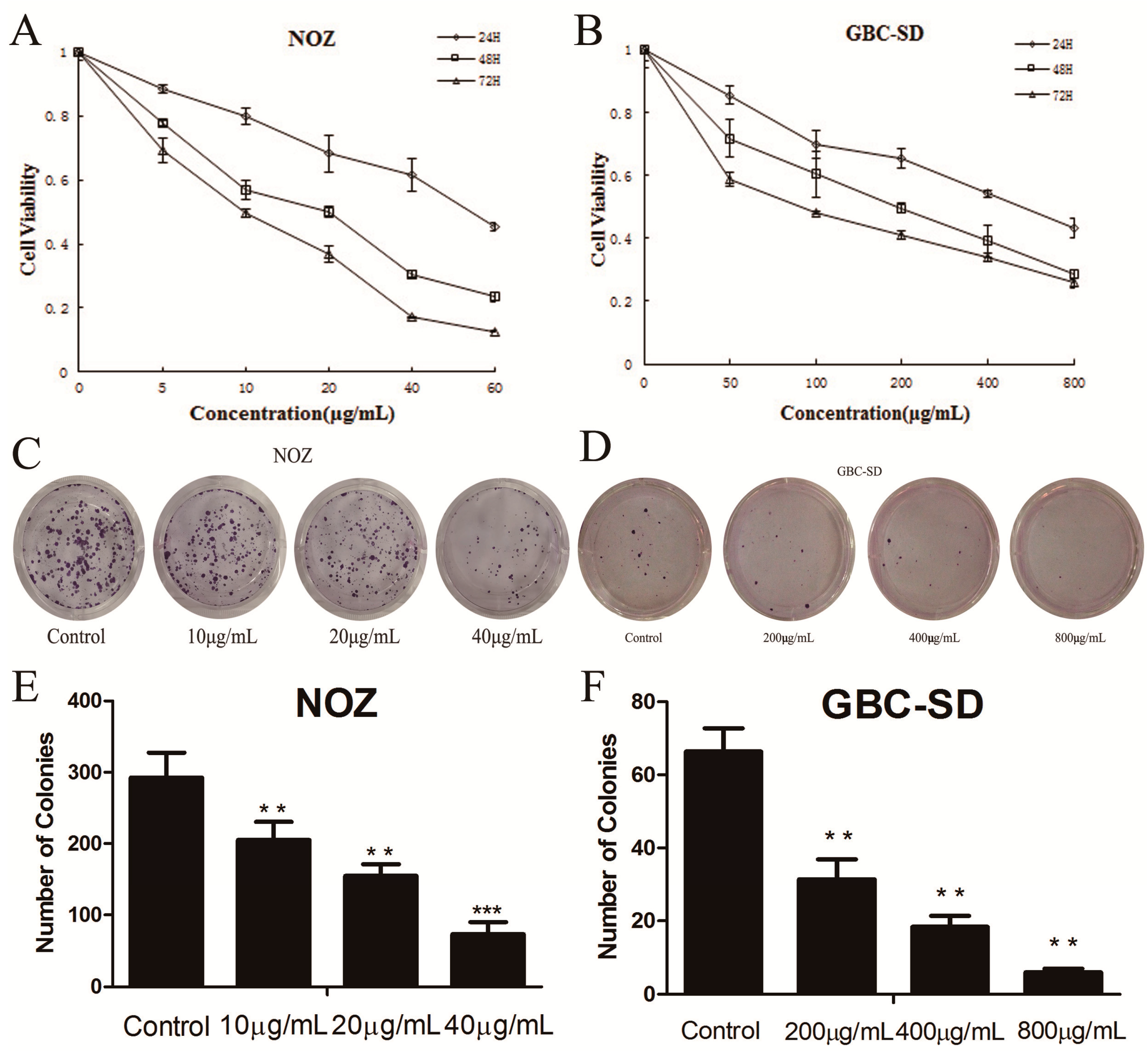
2.1. Cordycepin Inhibits Proliferation and Colony Formation of Gallbladder Cancer Cells

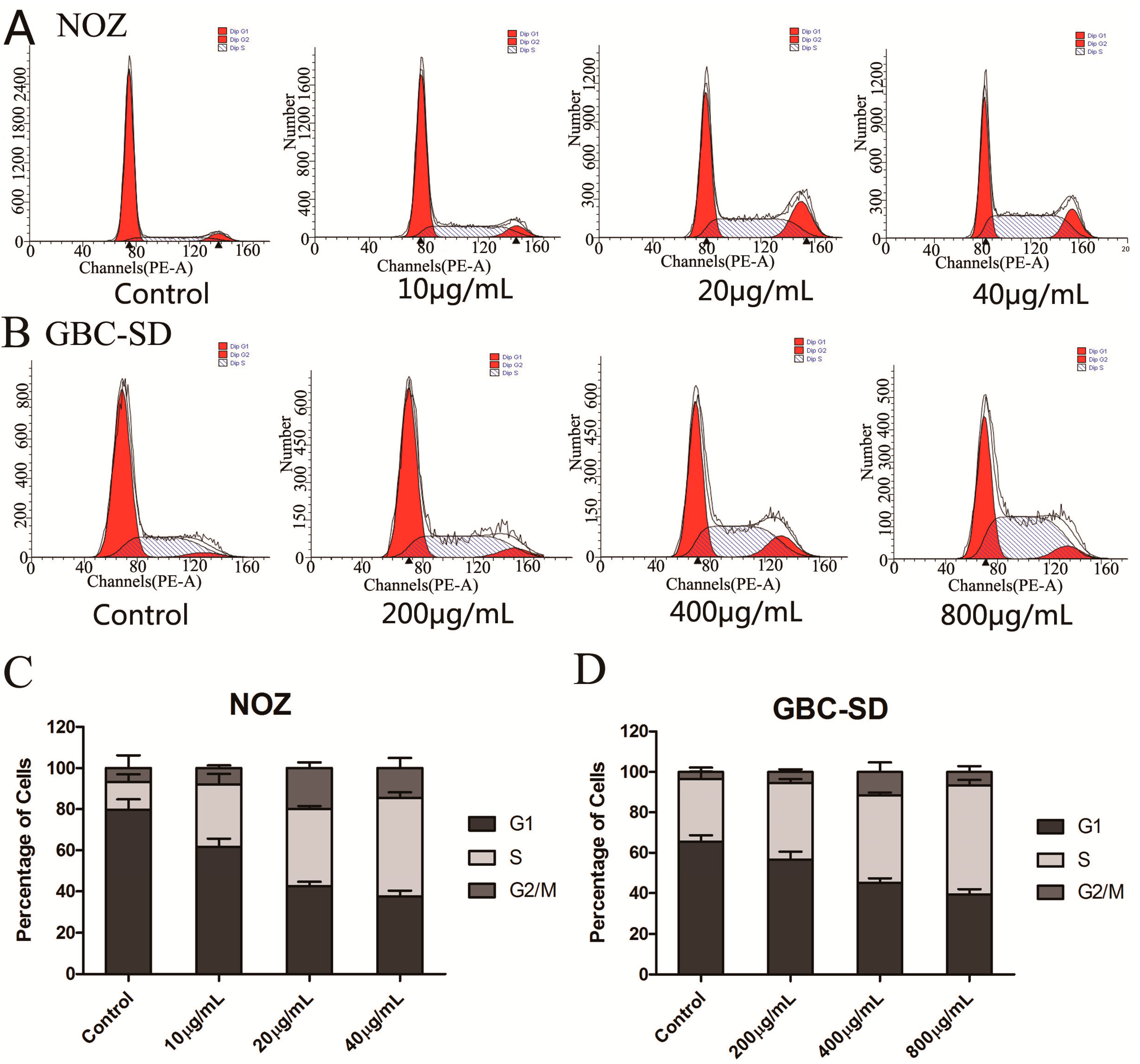
2.2. Cordycepin Induces S Phase Arrest and Regulates the Expression of Cell Cycle-Related Proteins in Gallbladder Cancer Cells

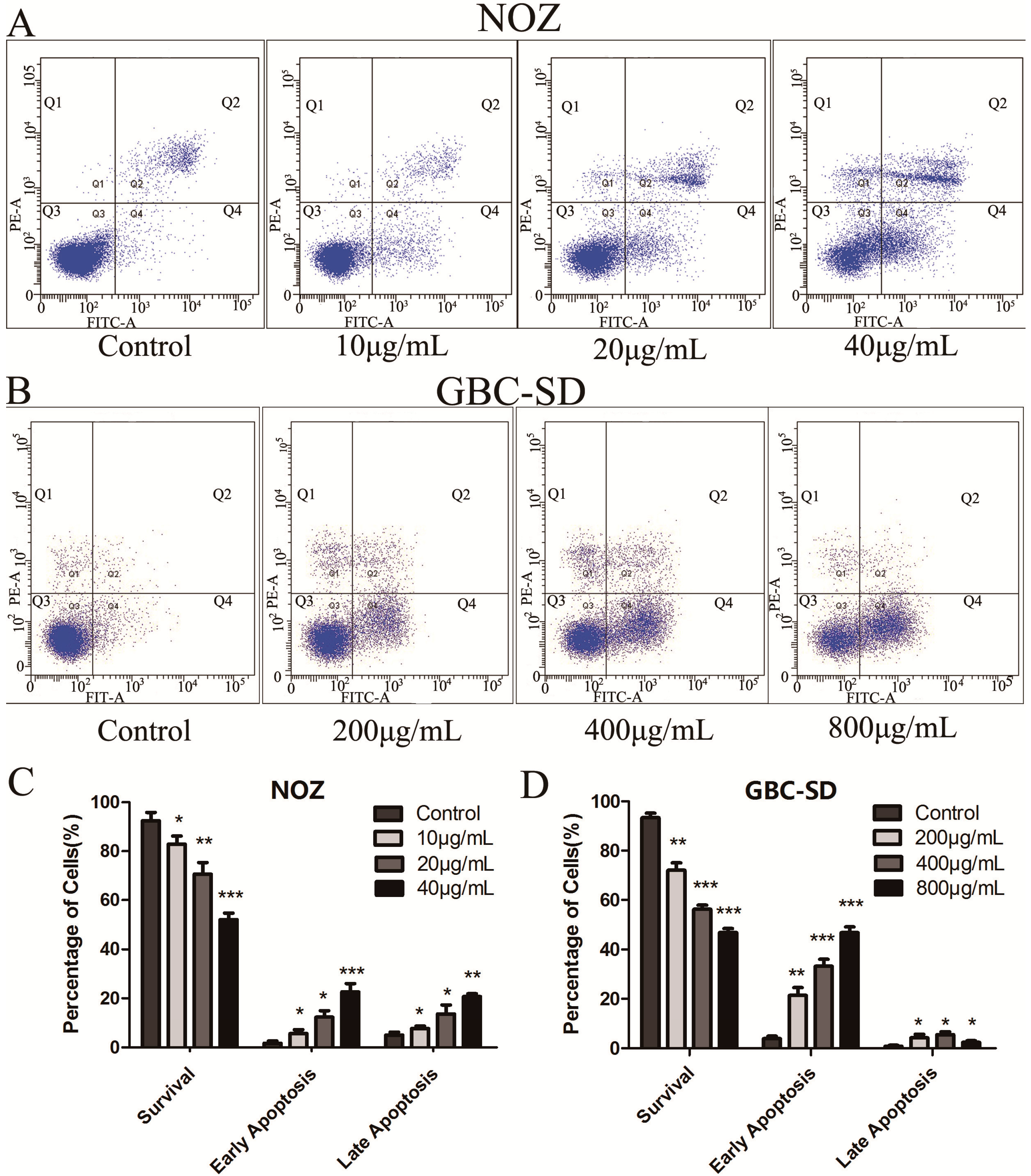
2.3. Cordycepin Induces Apoptosis in Human Gallbladder Cancer Cells

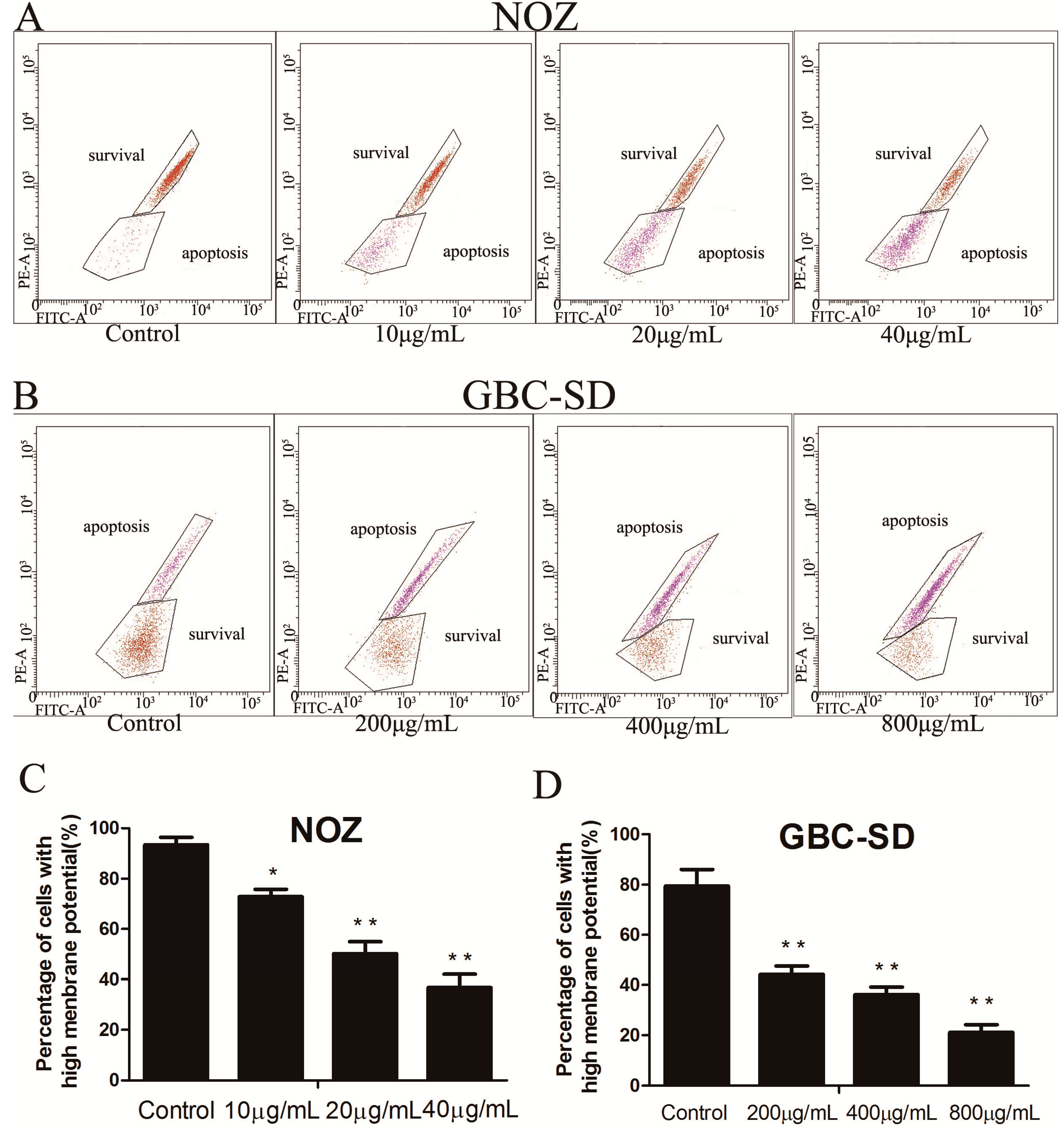
2.4. Cordycepin Reduces ΔΨm in Human Gallbladder Cancer Cells
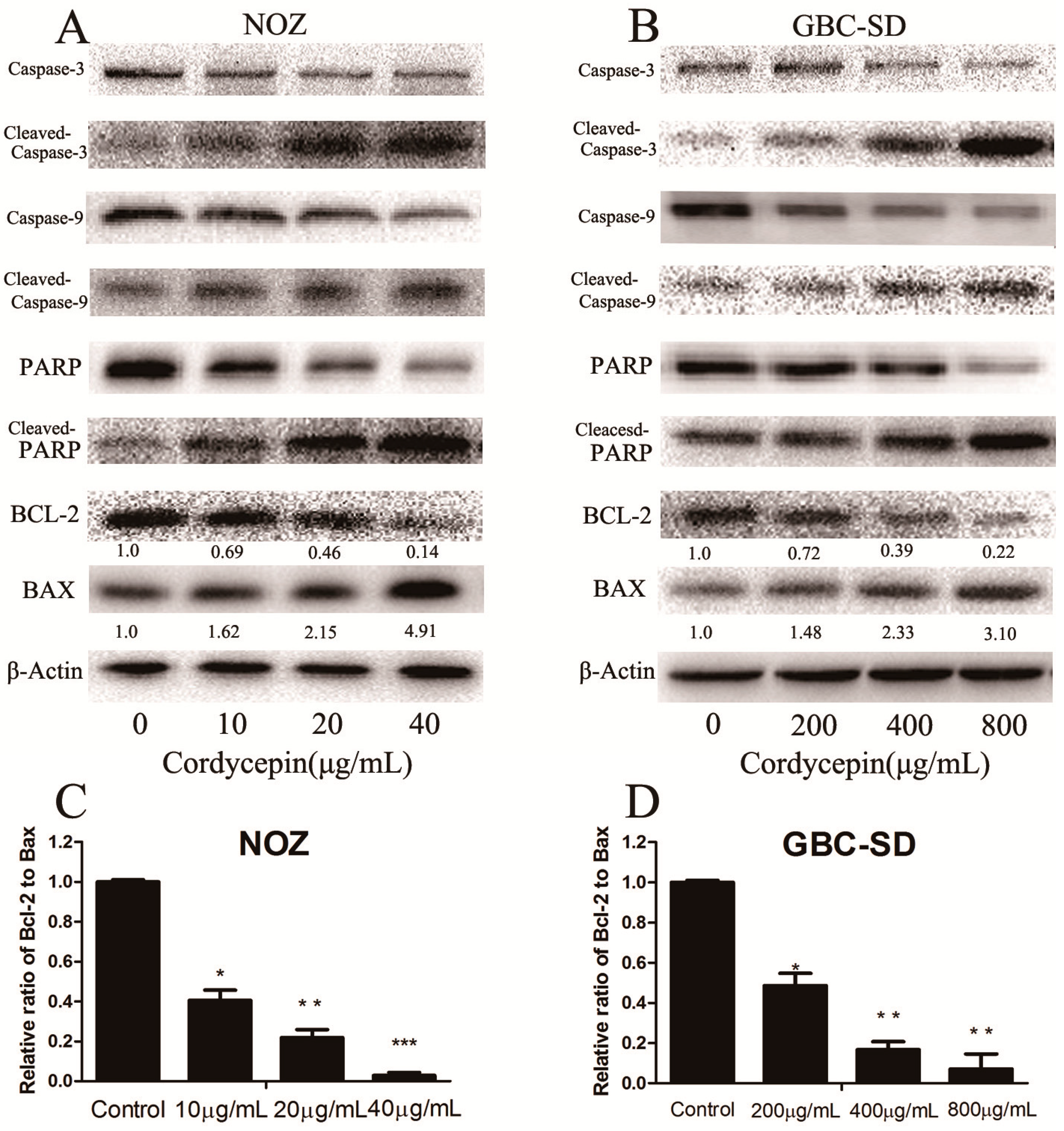
2.5. Cordycepin Induces Apoptosis by Regulating Bcl-2 Family Members and Caspase-3 in Human Gallbladder Cancer Cells


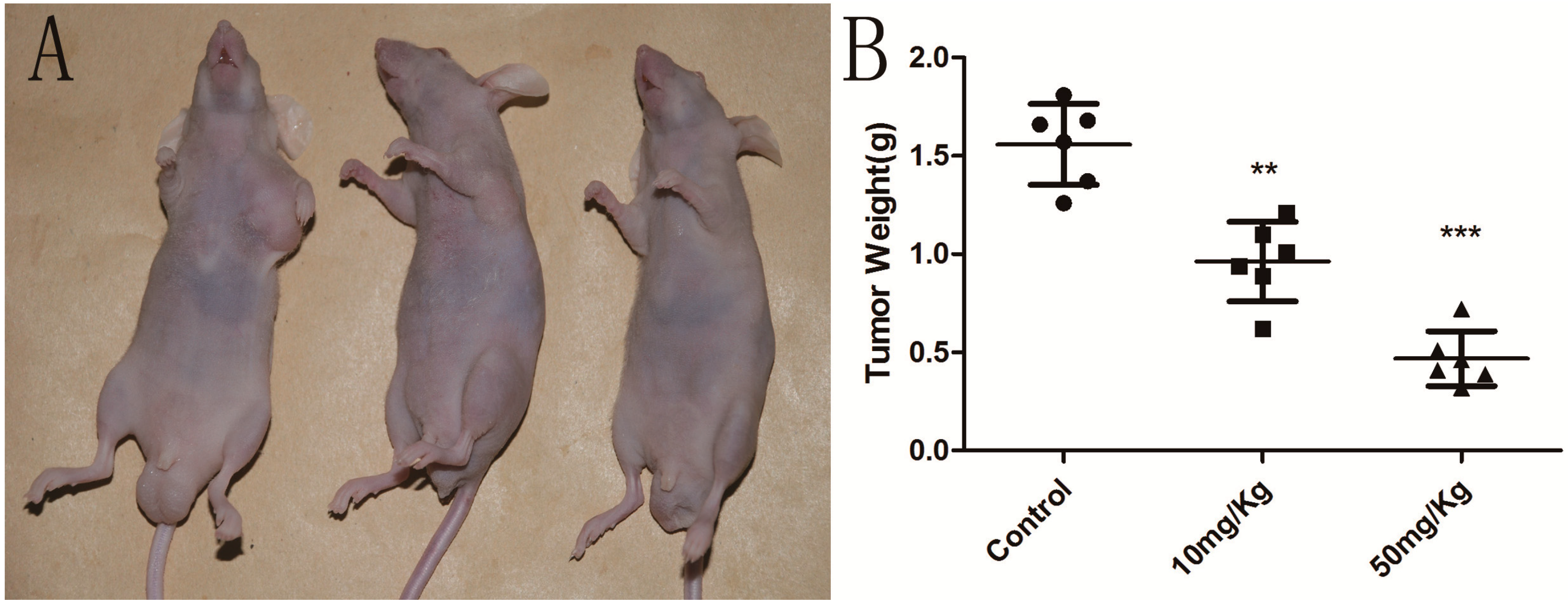
2.6. Cordycepin Suppresses Tumor Growth in Vivo

3. Experimental Section
3.1. Drugs and Antibodies
3.2. Cell Lines and Culture
3.3. Cell Proliferation Assay
3.4. Colony Formation Assay
3.5. Cell Cycle Analysis
3.6. Cell Apoptosis Assay
3.7. Mitochondrial Membrane Potential (ΔΨm) Assay
3.8. Western Blot Analysis
3.9. In Vivo Efficacy of Cordycepin
3.10. Statistical Analysis
4. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hueman, M.T.; Vollmer, C.M., Jr.; Pawlik, T.M. Evolving treatment strategies for gallbladder cancer. Ann. Surg. Oncol. 2009, 16, 2101–2115. [Google Scholar] [CrossRef]
- Dong, P.; He, X.W.; Gu, J.; Wu, W.G.; Li, M.L.; Yang, J.H.; Zhang, L.; Ding, Q.C.; Lu, J.H.; Mu, J.S.; et al. Vimentin significantly promoted gallbladder carcinoma metastasis. Chin. Med. J. 2011, 124, 4236–4244. [Google Scholar]
- Wang, J.W.; Peng, S.Y.; Li, J.T.; Wang, Y.; Zhang, Z.P.; Cheng, Y.; Cheng, D.Q.; Weng, W.H.; Wu, X.S.; Fei, X.Z.; et al. Identification of metastasis-associated proteins involved in gallbladder carcinoma metastasis by proteomic analysis and functional exploration of chloride intracellular channel 1. Cancer Lett. 2009, 281, 71–81. [Google Scholar] [CrossRef]
- Liu, T.Y.; Tan, Z.J.; Jiang, L.; Gu, J.F.; Wu, X.S.; Cao, Y.; Li, M.L.; Wu, K.J.; Liu, Y.B. Curcumin induces apoptosis in gallbladder carcinoma cell line GBC-SD cells. Cancer Cell Int. 2013, 13, 64. [Google Scholar] [CrossRef]
- Wu, X.S.; Wang, X.A.; Wu, W.G.; Hu, Y.P.; Li, M.L.; Ding, Q.; Weng, H.; Shu, Y.J.; Liu, T.Y.; Jiang, L.; et al. MALAT1 promotes the proliferation and metastasis of gallbladder cancer cells by activating the ERK/MAPK pathway. Cancer Biol. Ther. 2014, 15, 806–814. [Google Scholar] [CrossRef]
- Wistuba, II.; Gazdar, A.F. Gallbladder cancer: Lessons from a rare tumour. Nat. Rev. Cancer 2004, 4, 695–706. [Google Scholar] [CrossRef]
- Lai, C.H.; Lau, W.Y. Gallbladder cancer—a comprehensive review. The Surgeon 2008, 6, 101–110. [Google Scholar] [CrossRef]
- Boutros, C.; Gary, M.; Baldwin, K.; Somasundar, P. Gallbladder cancer: Past, present and an uncertain future. Surg. Oncol. 2012, 21, e183–e191. [Google Scholar] [CrossRef]
- Sharma, A.; Dwary, A.D.; Mohanti, B.K.; Deo, S.V.; Pal, S.; Sreenivas, V.; Raina, V.; Shukla, N.K.; Thulkar, S.; Garg, P.; et al. Best supportive care compared with chemotherapy for unresectable gall bladder cancer: A randomized controlled study. J. Clin. Oncol. 2010, 28, 4581–4586. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Manson, W.; Spring, F.S.; Hutchinson, S.A. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (Linn.). Nature 1950, 166, 949. [Google Scholar]
- Paterson, R.R. Cordyceps: A traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 2008, 69, 1469–1495. [Google Scholar] [CrossRef] [Green Version]
- Wong, Y.Y.; Moon, A.; Duffin, R.; Barthet-Barateig, A.; Meijer, H.A.; Clemens, M.J.; de Moor, C.H. Cordycepin inhibits protein synthesis and cell adhesion through effects on signal transduction. J. Biol. Chem. 2010, 285, 2610–2621. [Google Scholar]
- Won, S.Y.; Park, E.H. Anti-inflammatory and related pharmacological activities of cultured mycelia and fruiting bodies of Cordyceps militaris. J. Ethnopharmacol. 2005, 96, 555–561. [Google Scholar] [CrossRef]
- Ng, T.B.; Wang, H.X. Pharmacological actions of Cordyceps, a prized folk medicine. J. Pharm. Pharmacol. 2005, 57, 1509–1519. [Google Scholar] [CrossRef]
- Wu, W.C.; Hsiao, J.R.; Lian, Y.Y.; Lin, C.Y.; Huang, B.M. The apoptotic effect of cordycepin on human OEC-M1 oral cancer cell line. Cancer Chemother. Pharmacol. 2007, 60, 103–111. [Google Scholar] [CrossRef]
- Lee, S.Y.; Debnath, T.; Kim, S.K.; Lim, B.O. Anti-cancer effect and apoptosis induction of cordycepin through DR3 pathway in the human colonic cancer cell HT-29. Food Chem. Toxicol. 2013, 60, 439–447. [Google Scholar] [CrossRef]
- Nakamura, K.; Yoshikawa, N.; Yamaguchi, Y.; Kagota, S.; Shinozuka, K.; Kunitomo, M. Antitumor effect of cordycepin (3'-deoxyadenosine) on mouse melanoma and lung carcinoma cells involves adenosine A3 receptor stimulation. Anticancer Res. 2006, 26, 43–47. [Google Scholar]
- Lee, H.H.; Park, C.; Jeong, J.W.; Kim, M.J.; Seo, M.J.; Kang, B.W.; Park, J.U.; Kim, G.Y.; Choi, B.T.; Choi, Y.H.; et al. Apoptosis induction of human prostate carcinoma cells by cordycepin through reactive oxygen speciesmediated mitochondrial death pathway. Int. J. Oncol. 2013, 42, 1036–1044. [Google Scholar]
- Lu, H.; Li, X.; Zhang, J.; Shi, H.; Zhu, X.; He, X. Effects of cordycepin on HepG2 and EA.hy926 cells: Potential antiproliferative, antimetastatic and anti-angiogenic effects on hepatocellular carcinoma. Oncol. Lett. 2014, 7, 1556–1562. [Google Scholar]
- Hardie, D.G. AMP-activated/SNF1 protein kinases: conserved guardians of cellular energy. Nat. Rev. Mol. Cell. Biol. 2007, 8, 774–785. [Google Scholar] [CrossRef]
- Rehman, G.; Shehzad, A.; Khan, A.L.; Hamayun, M. Role of AMP-Activated Protein Kinase in Cancer Therapy. Arch. Pharm. 2014, 347, 457–468. [Google Scholar] [CrossRef]
- Nigg, E.A. Cyclin-dependent protein kinases: Key regulators of the eukaryotic cell cycle. Bioessays 1995, 17, 471–480. [Google Scholar] [CrossRef]
- Mitrea, D.M.; Yoon, M.K.; Ou, L.; Kriwacki, R.W. Disorder-function relationships for the cell cycle regulatory proteins p21 and p27. Biol. Chem. 2012, 393, 259–274. [Google Scholar]
- Yoon, M.K.; Mitrea, D.M.; Ou, L.; Kriwacki, R.W. Cell cycle regulation by the intrinsically disordered proteins p21 and p27. Biochem. Soc. Trans. 2012, 40, 981–988. [Google Scholar] [CrossRef]
- Kang, N.; Zhang, J.H.; Qiu, F.; Tashiro, S.; Onodera, S.; Ikejima, T. Inhibition of EGFR signaling augments oridonin-induced apoptosis in human laryngeal cancer cells via enhancing oxidative stress coincident with activation of both the intrinsic and extrinsic apoptotic pathways. Cancer Lett. 2010, 294, 147–158. [Google Scholar] [CrossRef]
- Kelly, P.N.; Strasser, A. The role of Bcl-2 and its pro-survival relatives in tumourigenesis and cancer therapy. Cell. Death Differentiation 2011, 18, 1414–1424. [Google Scholar] [CrossRef]
- Strasser, A.; Cory, S.; Adams, J.M. Deciphering the rules of programmed cell death to improve therapy of cancer and other diseases. The EMBO J. 2011, 30, 3667–3683. [Google Scholar] [CrossRef]
- Hu, W.; Kavanagh, J.J. Anticancer therapy targeting the apoptotic pathway. Lancet Oncol. 2003, 4, 721–729. [Google Scholar] [CrossRef]
- Spencer, S.L.; Sorger, P.K. Measuring and modeling apoptosis in single cells. Cell 2011, 144, 926–939. [Google Scholar] [CrossRef]
- Thomadaki, H.; Scorilas, A.; Tsiapalis, C.M.; Havredaki, M. The role of cordycepin in cancer treatment via induction or inhibition of apoptosis: Implication of polyadenylation in a cell type specific manner. Cancer Chemother. Pharmacol. 2008, 61, 251–265. [Google Scholar]
- Jeong, J.W.; Jin, C.Y.; Park, C.; Hong, S.H.; Kim, G.Y.; Jeong, Y.K.; Lee, J.D.; Yoo, Y.H.; Choi, Y.H. Induction of apoptosis by cordycepin via reactive oxygen species generation in human leukemia cells. Toxicol. In Vitro 2011, 25, 817–824. [Google Scholar] [CrossRef]
- He, W.; Zhang, M.F.; Ye, J.; Jiang, T.T.; Fang, X.; Song, Y. Cordycepin induces apoptosis by enhancing JNK and p38 kinase activity and increasing the protein expression of Bcl-2 pro-apoptotic molecules. J. Zhejiang Univ. Sci. B 2010, 11, 654–660. [Google Scholar]
- Burlacu, A. Regulation of apoptosis by Bcl-2 family proteins. J. Cell. Mol. Med. 2003, 7, 249–257. [Google Scholar] [CrossRef]
- Wei, M.C.; Zong, W.X.; Cheng, E.H.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacGregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef]
- Lindsay, J.; Esposti, M.D.; Gilmore, A.P. Bcl-2 proteins and mitochondria--specificity in membrane targeting for death. Biochim. Biophys. Acta 2011, 1813, 532–539. [Google Scholar] [CrossRef]
- Walensky, L.D. BCL-2 in the crosshairs: Tipping the balance of life and death. Cell Death Differ. 2006, 13, 1339–1350. [Google Scholar] [CrossRef]
- Mao, W.P.; Ye, J.L.; Guan, Z.B.; Zhao, J.M.; Zhang, C.; Zhang, N.N.; Jiang, P.; Tian, T. Cadmium induces apoptosis in human embryonic kidney (HEK) 293 cells by caspase-dependent and -independent pathways acting on mitochondria. Toxicol. In Vitro 2007, 21, 343–354. [Google Scholar] [CrossRef]
- Yoshikawa, N.; Nakamura, K.; Yamaguchi, Y.; Kagota, S.; Shinozuka, K.; Kunitomo, M. Antitumour activity of cordycepin in mice. Clin. Exp. Pharmacol. Physiol. 2004, 31, S51–53. [Google Scholar] [CrossRef]
- Gao, J.; Lian, Z.Q.; Zhu, P.; Zhu, H.B. Lipid-lowering effect of cordycepin (3'-deoxyadenosine) from Cordyceps militaris on hyperlipidemic hamsters and rats. Yao Xue Xue Bao 2011, 46, 669–676. [Google Scholar]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, X.-A.; Xiang, S.-S.; Li, H.-F.; Wu, X.-S.; Li, M.-L.; Shu, Y.-J.; Zhang, F.; Cao, Y.; Ye, Y.-Y.; Bao, R.-F.; et al. Cordycepin Induces S Phase Arrest and Apoptosis in Human Gallbladder Cancer Cells. Molecules 2014, 19, 11350-11365. https://doi.org/10.3390/molecules190811350
Wang X-A, Xiang S-S, Li H-F, Wu X-S, Li M-L, Shu Y-J, Zhang F, Cao Y, Ye Y-Y, Bao R-F, et al. Cordycepin Induces S Phase Arrest and Apoptosis in Human Gallbladder Cancer Cells. Molecules. 2014; 19(8):11350-11365. https://doi.org/10.3390/molecules190811350
Chicago/Turabian StyleWang, Xu-An, Shan-Shan Xiang, Huai-Feng Li, Xiang-Song Wu, Mao-Lan Li, Yi-Jun Shu, Fei Zhang, Yang Cao, Yuan-Yuan Ye, Run-Fa Bao, and et al. 2014. "Cordycepin Induces S Phase Arrest and Apoptosis in Human Gallbladder Cancer Cells" Molecules 19, no. 8: 11350-11365. https://doi.org/10.3390/molecules190811350