Pyrazinecarboxamides as Potential Elicitors of Flavonolignan and Flavonoid Production in Silybum marianum and Ononis arvensis Cultures In Vitro

Abstract
:1. Introduction

2. Results and Discussion

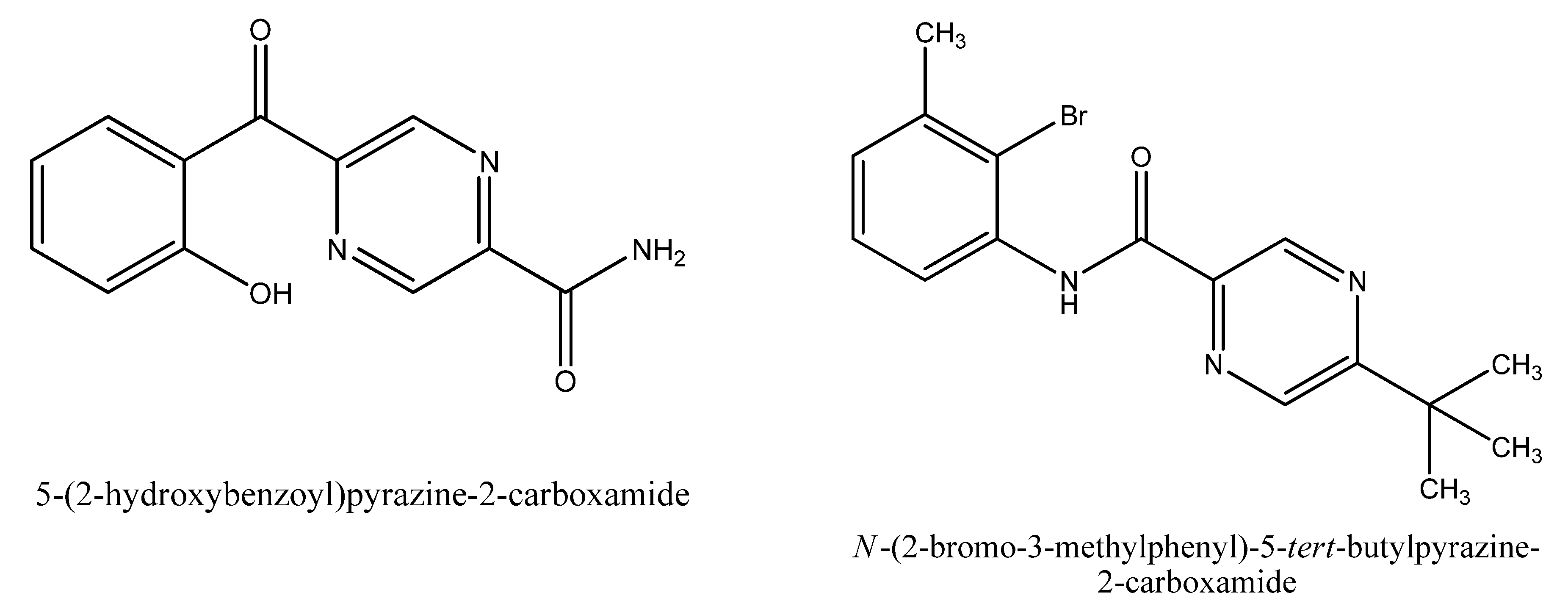{kind=link}
| Compound I (conc. mol/L) | Time of sampling (hours) | TAX (%) | SILYD (%) | SILYCHR (%) | ISOSILYB A (%) | ISOSILYB B (%) | SILYB A (%) | SILYB B (%) | SILYM COMP (%) |
|---|---|---|---|---|---|---|---|---|---|
| C1 = 1.159 × 10−3 | control | 0 | - | 0 | - | 0 | - | - | 0 |
| 6 | 0 | - | - | 0 | - | 0 | - | 0 | |
| 12 | 0 | 0.04 | 0.01 | - | - | - | - | 0.05 | |
| 24 | 0 | 0.04 | 0 | - | - | - | - | 0.04 | |
| 48 | 0 | - | 0 | 0 | 0 | - | - | 0 | |
| 72 | 0 | - | - | 0 | 0 | - | 0 | 0 | |
| 168 | 0 | 0 | 0 | 0 | 0 | 0 | - | 0 | |
| C2 = 1.159 × 10−4 | control | 0 | - | 0.01 | - | - | - | - | 0.01 |
| 6 | 0 | - | 0.01 | - | - | - | - | 0.01 | |
| 12 | 0 | - | 0.01 | - | 0 | - | - | 0.01 | |
| 24 | 0 | 0.03 | 0 | - | - | - | - | 0.03 | |
| 48 | 0 | 0.03 | 0 | - | - | - | - | 0.03 | |
| 72 | 0 | - | - | - | - | 0 | 0.01 | 0.01 | |
| 168 | 0 | 0.03 | 0.01 | 0.04 | - | - | - | 0.08 | |
| C3 = 1.159 × 10−5 | control | 0 | - | 0.02 | - | - | - | - | 0,02 |
| 6 | 0.02 | 0 | - | - | - | - | - | 0 | |
| 12 | 0 | 0.02 | 0 | 0 | - | - | - | 0.02 | |
| 24 | 0 | 0.03 | 0.01 | 0 | - | - | 0 | 0.04 | |
| 48 | 0 | 0.01 | - | - | - | - | - | 0.01 | |
| 72 | 0 | 0.01 | 0 | - | - | - | - | 0.01 | |
| 168 | 0 | 0 | 0 | - | - | - | - | 0 |
| Compound I (conc. mol/L) | Time of sampling (hours) | TAX (%) | SILYD (%) | SILYCHR (%) | ISOSILYB A (%) | ISOSILYB B (%) | SILYB A (%) | SILYB B (%) | SILYM COMP (%) |
|---|---|---|---|---|---|---|---|---|---|
| C1 = 1.159 × 10−3 | control | 0.01 | 0 | 0.03 | - | 0 | - | 0 | 0.06 |
| 6 | 0 | 0.02 | - | - | - | - | - | 0.02 | |
| 12 | 0 | 0.02 | - | - | - | - | - | 0.02 | |
| 24 | 0 | 0.11 | 0 | - | - | - | - | 0.11 | |
| 48 | 0 | 0.01 | 0 | 0.01 | - | - | - | 0.02 | |
| 72 | 0 | 0 | - | - | 0.01 | - | - | 0.01 | |
| 168 | 0 | 0.01 | - | - | 0 | - | - | 0.02 | |
| C2 = 1.159 × 10−4 | control | 0 | - | 0.01 | - | - | 0 | - | 0.01 |
| 6 | 0 | - | 0 | - | - | - | - | 0 | |
| 12 | 0 | - | - | - | 0 | 0.01 | - | 0.01 | |
| 24 | 0 | 0 | 0 | - | 0 | - | - | 0 | |
| 48 | 0.01 | 0 | - | - | - | 0 | 0 | ||
| 72 | 0 | - | 0 | 0 | 0 | - | - | 0 | |
| 168 | 0 | - | 0 | - | 0 | 0 | 0 | 0 | |
| C3 = 1.159 × 10−5 | control | 0 | - | 0.01 | - | - | - | - | 0.01 |
| 6 | 0 | 0 | - | - | - | - | - | 0 | |
| 12 | 0 | 0.01 | - | 0 | - | - | - | 0 | |
| 24 | 0 | - | - | - | 0 | - | - | 0.01 | |
| 48 | 0 | - | 0 | - | - | 0 | 0 | 0 | |
| 72 | 0 | - | 0 | - | - | 0 | - | 0 | |
| 168 | 0 | - | 0 | - | - | - | - | 0 |
2.1. Silybum marianum Callus Culture
2.2. Silybum marianum Suspension Culture
2.3. Flavonolignans in Nutrient Medium
| Compound I (concentration mol/L) | Time of sampling (hours) | TAXIFOLIN (%) | SILYCHRISTIN (%) | SILYDIANIN (%) |
|---|---|---|---|---|
| c1 = 1.159 × 10−3 | control | 0.02 | 0 | 0.01 |
| 6 | 0.01 | 0 | 0.09 | |
| 24 | 0.05 | 0 | 0 | |
| 48 | 0.01 | 0 | 0.01 | |
| 72 | 0 | 0 | 0.21 | |
| 168 | 0.03 | 0 | 0 | |
| c2 = 1.159 × 10 − 4 | 12 | 0.04 | 0 | 0 |
| 24 | 0 | 0 | 0 | |
| 72 | 0.06 | 0 | 0 | |
| c3 = 1.159 × 10 − 5 | 6 | 0.04 | 0 | 0 |
| 12 | 0 | 0 | 0 | |
| 24 | 0 | 0 | 0 | |
| 48 | 0 | 0 | 0 | |
| 168 | 0 | 0 | 0.05 |
2.4. Ononis arvensis Callus Culture
| Compound II (concentration mol/L) | Time of sampling (hours) | Flavonoid content (%) | SD |
|---|---|---|---|
| c1 = 8.36.10−4 | control | 0.01 | 0.002 |
| 6 | 0.025 * | 0.004 | |
| 12 | 0.06 * | 0.004 | |
| 24 | 0.023 * | 0.001 | |
| 48 | 0.057 * | 0.002 | |
| 72 | 0.017 * | 0.003 | |
| 168 | 0.003 | 0.001 | |
| c2 = 8.36.10−5 | control | 0.025 | 0.003 |
| 6 | 0.079 * | 0.007 | |
| 12 | 0.063 * | 0.002 | |
| 24 | 0.020 | 0.002 | |
| 48 | 0.008 | 0.003 | |
| 72 | 0.006 | 0.002 | |
| 168 | 0.016 | 0.003 | |
| c3 = 8.36.10−6 | control | 0.016 | 0.003 |
| 6 | 0.218 * | 0.005 | |
| 12 | 0.141 * | 0.001 | |
| 24 | 0.011 | 0.004 | |
| 48 | 0.123 * | 0.002 | |
| 72 | 0.156 * | 0.002 | |
| 168 | 0.241 * | 0.003 |
2.5. Ononis arvensis Suspension Culture
| Compound II (concentration mol/L) | Time of sampling (hours) | Flavonoid content (%) | SD |
|---|---|---|---|
| c1 = 8.36.10−4 | control | 0.008 | 0.002 |
| 6 | 0.017 * | 0.002 | |
| 12 | 0.004 | 0.003 | |
| 24 | 0.010 | 0.005 | |
| 48 | 0.210 * | 0.003 | |
| 72 | 0.003 | 0.001 | |
| 168 | 0.002 | 0.001 | |
| c2 = 8.36.10−5 | control | 0.045 | 0.005 |
| 6 | 0.002 | 0.002 | |
| 12 | 0.006 | 0.002 | |
| 24 | 0.001 | 0.001 | |
| 48 | 0.004 | 0.002 | |
| 72 | 0.020 | 0.004 | |
| 168 | 0.041 | 0.003 | |
| c3 = 8.36.10−6 | control | 0.002 | 0.001 |
| 6 | 0.044 * | 0.004 | |
| 12 | 0.105 * | 0.007 | |
| 24 | 0.073 * | 0.005 | |
| 48 | 0.118 * | 0.006 | |
| 72 | 0.059 * | 0.003 | |
| 168 | 0.039 * | 0.002 |
3. Experimental
3.1. Plant Material
3.2. Elicitors
3.3. Analysis of Flavonolignans and Flavonoids
4. Conclusions
Acknowledgments
References
- Verpoorte, R.; van der Heijden, R.; ten Hoopen, H.G.J.; Memelink, J. Metabolic engineering of plant secondary metabolite pathways for the production of fine chemicals. Biotechnol. Lett. 1999, 21, 467–479. [Google Scholar] [CrossRef]
- Tumova, L.; Psotova, R. UV-radiation and the flavonoid content in callus culture of Ononis arvensis L. Herba Polonica 1998, 44, 22–26. [Google Scholar]
- Dolezal, M.; Zitko, J.; Kesetovicova, D.; Kunes, J.; Svobodova, M. Substituted N-phenylpyrazine-2-carboxamides: Synthesis and antimycobacterial evaluation. Molecules 2009, 14, 4180–4189. [Google Scholar] [CrossRef]
- Dolezal, M.; Tumova, L.; Kesetovicova, D.; Tuma, J.; Kralova, K. Substituted N-phenylpyrazine-2-carboxamides, their synthesis and evaluation as herbicides and abioticitic elicitors. Molecules 2007, 12, 2587–2598. [Google Scholar]
- Tumova, L.; Tuma, J.; Dolezal, M.; Megusar, K. Substituted pyrazinecarboxamides as abiotic elicitors of the flavonolignan production in Silybum marianum Culture in vitro. Molecules 2010, 15, 331–340. [Google Scholar] [CrossRef]
- Tumova, L.; Gallova, K.; Rimakova, J.; Dolezal, M.; Tuma, J. The effect of substituted amide of pyrazine-2-carboxylic acids on flavonolignan production in Silybum marianum culture in vitro. Acta Phys. Planta. 2005, 27, 357–362. [Google Scholar] [CrossRef]
- Dolezal, M.; Jampilek, J.; Osicka, Z.; Kunes, J.; Buchta, V.; Vichova, P. Substituted 5-aroylpyrazine-2-carboxylic acid derivatives: Synthesis and biological activity. Farmaco 2003, 58, 1105–1111. [Google Scholar] [CrossRef]
- Wichlt, M. Herbal Drugs and Phytopharmaceuticals; Medfarm Scietific Publishers: Stuttgart, Germany, 2000; pp. 121–123. [Google Scholar]
- Wichtl, M. Herbal Drugs and Phytopharmaceuticals; Medfarm Scietific Publishers: Stuttgart, Germany, 2000; pp. 355–357. [Google Scholar]
- Gazak, R.; Walterova, D.; Kren, V. Silybin and silymarin—New and emerging application ion in medicine. Curr. Med. Chem. 2007, 14, 315–338. [Google Scholar] [CrossRef]
- Šeršeň, F.; Vencel, T.; Annus, J. Silymarin and its components scavenge phenylglyoxylic ketyl radicals. Fitoterapia 2006, 77, 525–529. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with Tobacco tissue cultures. Physiol Plant 1962, 15, 437–497. [Google Scholar] [CrossRef]
- Czech Pharmacopeia; Grada Publishing a.s.: Prague, Czech Republic, 2009.
- Czech Pharmacopeia; Grada Publishing a.s: Prague, Czech Republi, 2005.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tumova, L.; Tuma, J.; Dolezal, M. Pyrazinecarboxamides as Potential Elicitors of Flavonolignan and Flavonoid Production in Silybum marianum and Ononis arvensis Cultures In Vitro. Molecules 2011, 16, 9142-9152. https://doi.org/10.3390/molecules16119142
Tumova L, Tuma J, Dolezal M. Pyrazinecarboxamides as Potential Elicitors of Flavonolignan and Flavonoid Production in Silybum marianum and Ononis arvensis Cultures In Vitro. Molecules. 2011; 16(11):9142-9152. https://doi.org/10.3390/molecules16119142
Chicago/Turabian StyleTumova, Lenka, Jiri Tuma, and Martin Dolezal. 2011. "Pyrazinecarboxamides as Potential Elicitors of Flavonolignan and Flavonoid Production in Silybum marianum and Ononis arvensis Cultures In Vitro" Molecules 16, no. 11: 9142-9152. https://doi.org/10.3390/molecules16119142