
Fe, Mn and 238U Accumulations in Phragmites australis Naturally Growing at the Mill Tailings Pond; Iron Plaque Formation Possibly Related to Root-Endophytic Bacteria Producing Siderophores

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Collection of P. australis
2.2. Analysis of Element Concentrations in Plants, and Calculations of Translocation Factor and Bioconcentration Factor
2.3. Observation of Iron Localization in P. australis Nodal Roots
2.4. Isolation of Endophytic Bacteria from P. australis Nodal Roots, Evaluation of Siderophore-Production Activity by CAS-Fe Assay and Identification
2.4.1. Isolation of Endophytic Bacteria
2.4.2. Evaluation of Siderophore-Production Activity of Endophytic Bacteria on CAS-Fe Medium and Identification
2.5. Confirmation of Precipitation by Bacterial Culture Filtrate with Fe Ions and Phenolic Production by HPLC
2.6. Statistical Analysis
3. Results
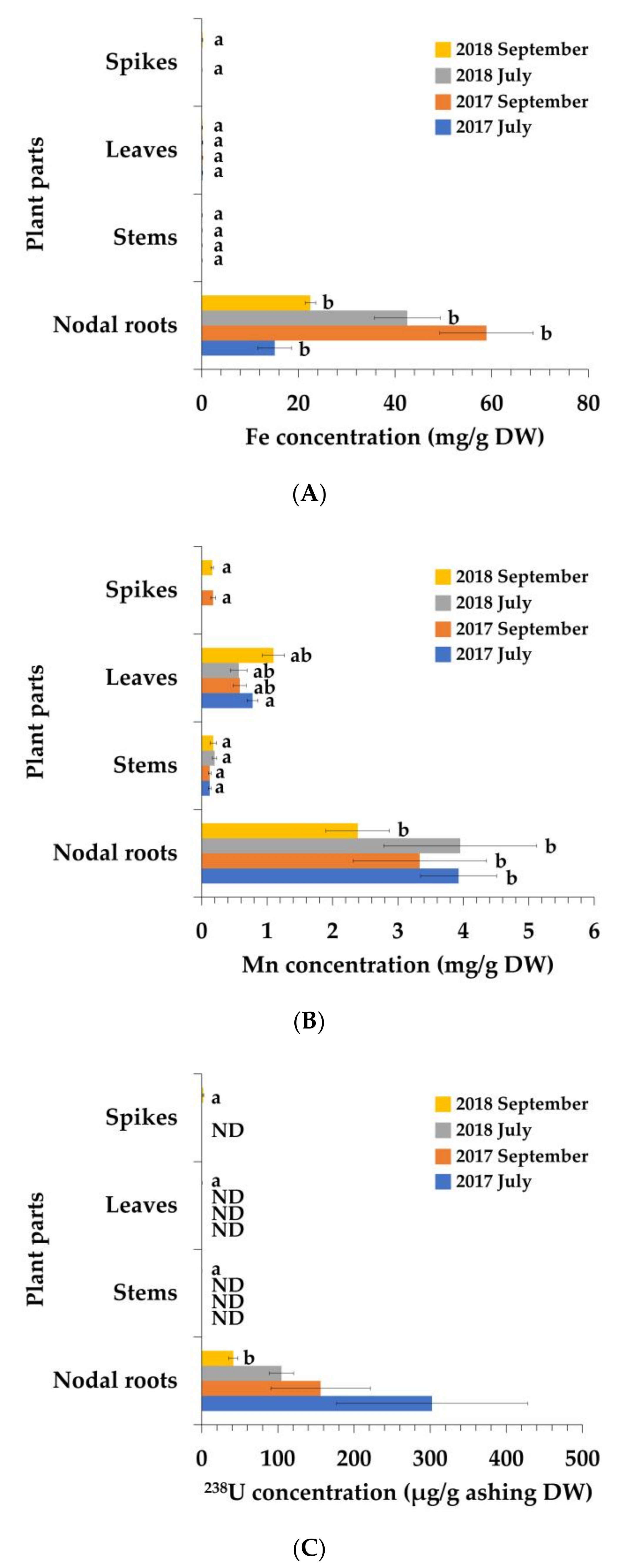
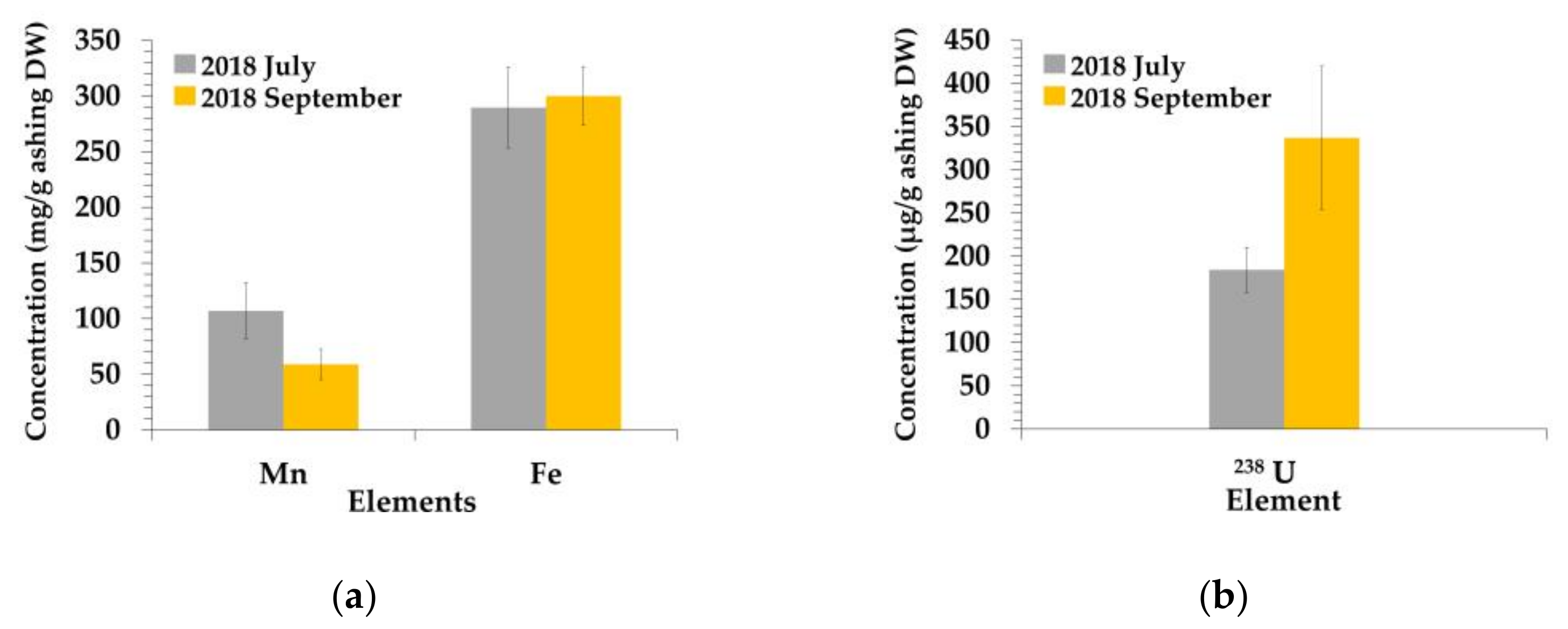
3.1. Elemental Concentrations of P. australis
3.2. Translocation Factors (TFs) and Bioconcentration Factors (BCFs)
3.3. Fe Localization in the Nodal Roots of P. australis
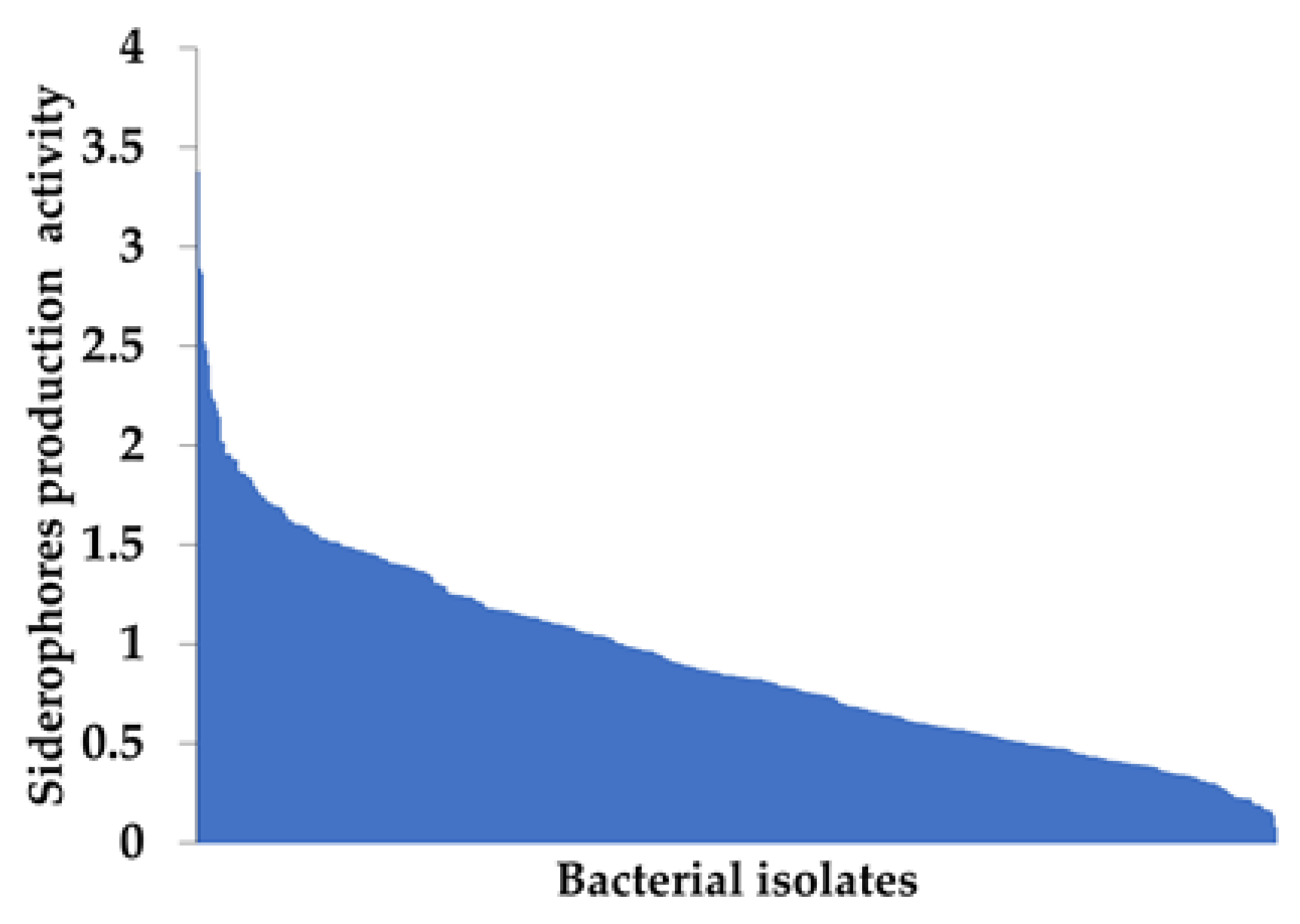
3.4. Siderophore Production Activity by Root-Endophytic Bacteria Isolated from P. australis
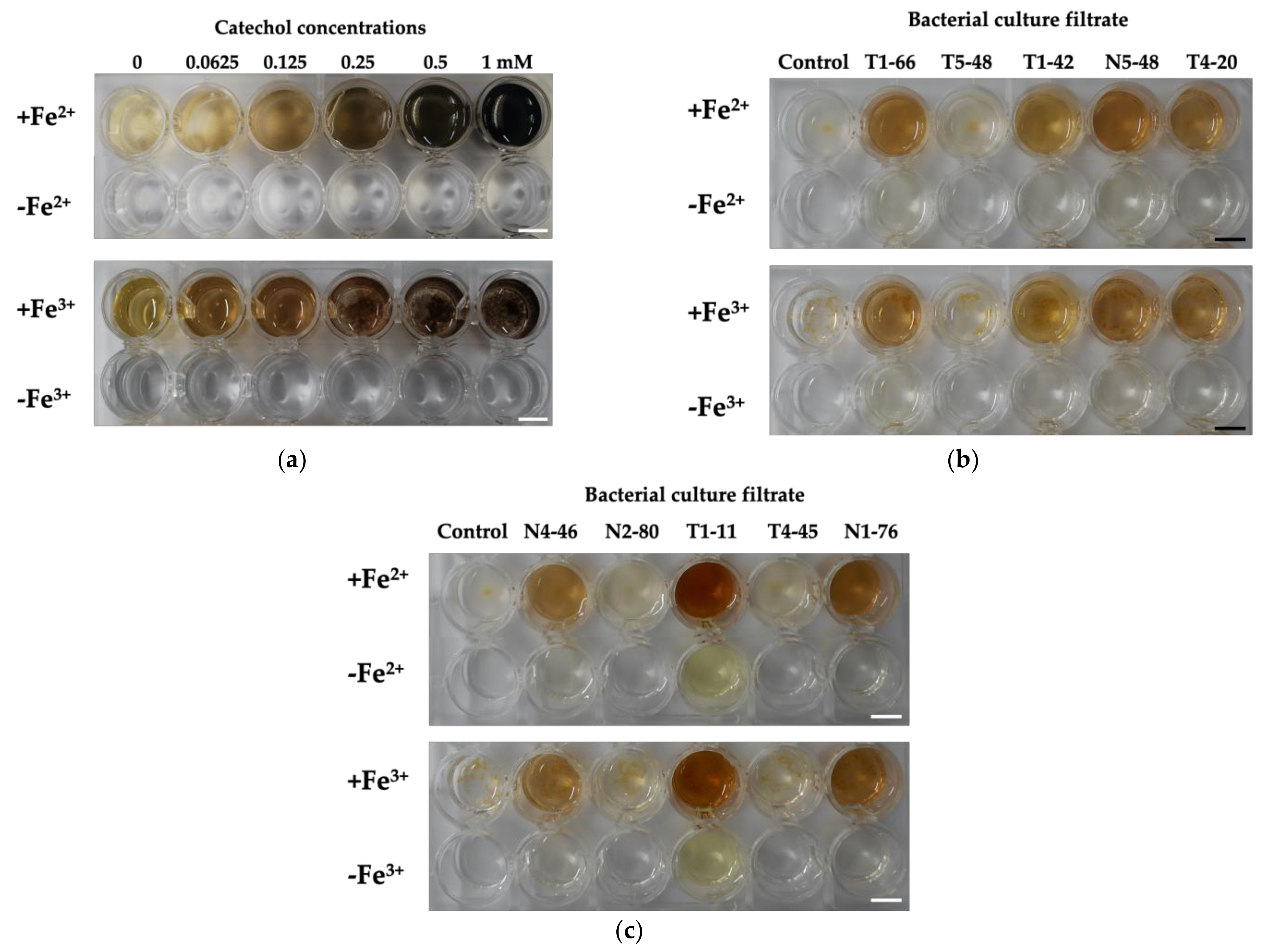
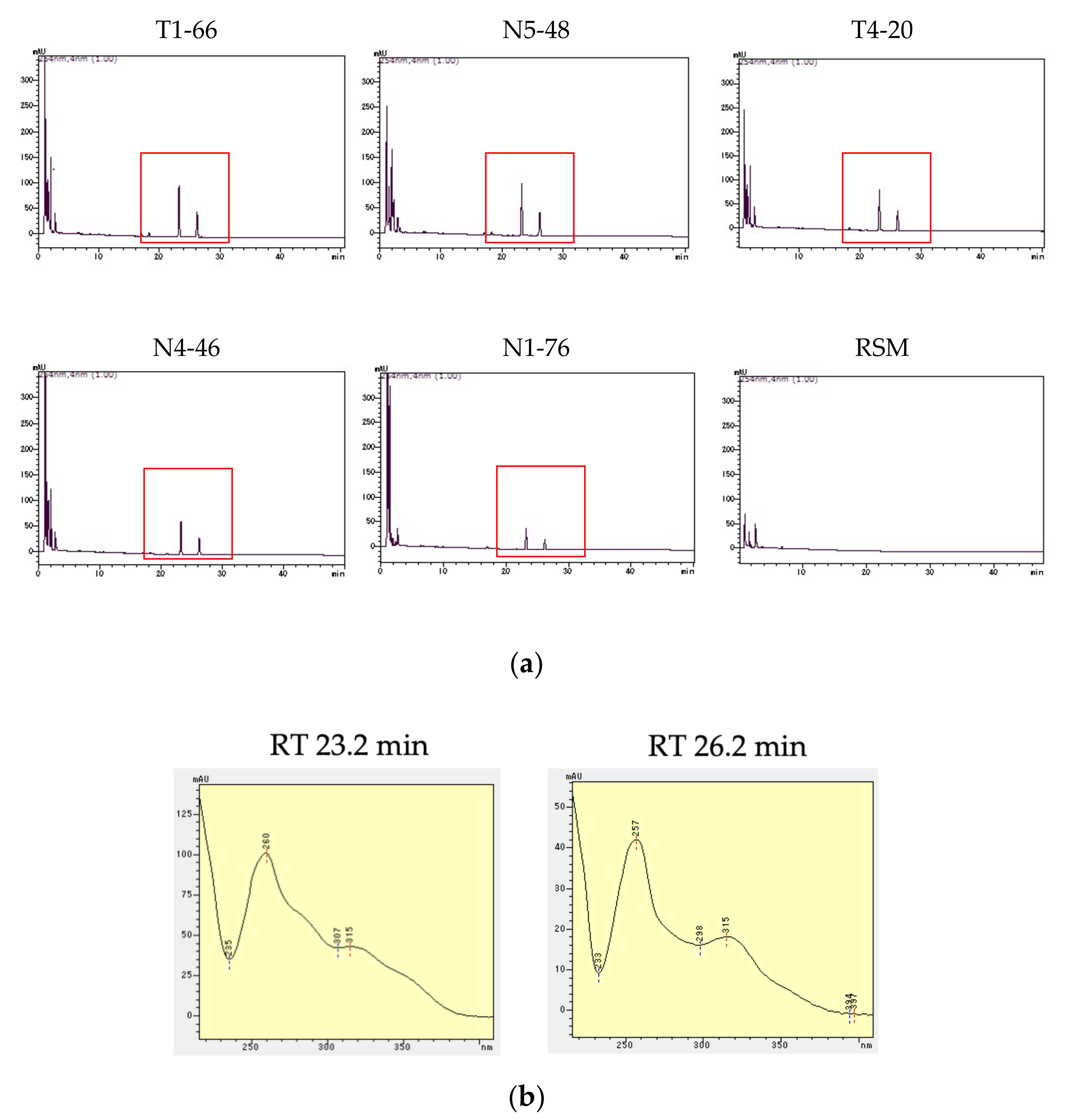
3.5. Confirmation of Precipitation by Bacterial Culture Filtrate with Fe Ions and Phenolic Production by HPLC
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dushenkov, V.; Kumar, P.B.A.N.; Motto, H.; Raskin, I. Rhizofiltration: The use of plants to remove heavy metals from aqueous streams. Environ. Sci. Technol. 1995, 29, 1239–1245. [Google Scholar] [CrossRef]
- Janson, C.E.; Kenson, R.E.; Tucker, L.H. Treatment of heavy metals in wastewaters. What wastewater-treatment method is most cost-effective for electroplating and finishing operations? Here are the alternatives. Environ. Prog. 1982, 1, 212–216. [Google Scholar] [CrossRef]
- Sheoran, A.S.; Sheoran, V. Heavy metal removal mechanism of acid mine drainage in wetlands: A critical review. Miner. Eng. 2006, 19, 105–116. [Google Scholar] [CrossRef]
- Dunbabin, J.S.; Bowmer, K.H. Potential use of constructed wetlands for treatment of industrial wastewaters containing metals. Sci. Total Environ. 1992, 111, 151–168. [Google Scholar] [CrossRef]
- Prokop, Z.; Cupr, P.; Zlevorova-Zlamalikova, V.; Komárek, J.; Dusek, L.; Holoubek, I. Mobility, bioavailability, and toxic effects of cadmium in soil samples. Environ. Res. 2003, 91, 119–126. [Google Scholar] [CrossRef]
- Rezania, S.; Park, J.; Rupani, P.F.; Darajeh, N.; Xu, X.; Shahrokhishahraki, R. Phytoremediation potential and control of Phragmites australis as a green phytomass: An overview. Environ. Sci. Pollut. Res. 2019, 26, 7428–7441. [Google Scholar] [CrossRef] [PubMed]
- Weis, J.S.; Weis, P. Metal uptake, transport and release by wetland plants: Implications for phytoremediation and restoration. Environ. Int. 2004, 30, 685–700. [Google Scholar] [CrossRef]
- Nagayasu, T.; Taki, T.; Fukushima, S. History and current situation of mine water treatment in Ningyo-toge uranium mine. JAEA Technol. 2017, 31, 1–87. (In Japanese) [Google Scholar] [CrossRef]
- Shitaka, Y.; Takayuki, T.; Sato, K. Evaluation of the variation of 226Ra and Ca uptake by plant and their relationship. J. Nucl. Sci. Technol. 2002, 39, 958–961. [Google Scholar] [CrossRef] [Green Version]
- Vymazal, J. Emergent plants used in free water surface constructed wetlands: A review. Ecol. Eng. 2013, 61, 582–592. [Google Scholar] [CrossRef]
- Armstrong, W. Aeration in higher plants. Adv. Bot. Res. 1980, 7, 225–332. [Google Scholar] [CrossRef]
- Ederli, L.; Reale, L.; Ferranti, F.; Pasqualini, S. Responses induced by high concentration of cadmium in Phragmites australis roots. Physiol. Plant. 2004, 121, 66–74. [Google Scholar] [CrossRef]
- Peverly, J.H.; Surface, J.M.; Wang, T. Growth and trace metal absorption by Phragmites australis in wetlands constructed for landfill leachate treatment. Ecol. Eng. 1995, 5, 21–35. [Google Scholar] [CrossRef]
- Wang, W.; Dudel, E.G. Fe plaque-related aquatic uranium retention via rhizofiltration along a redox-state gradient in a natural Phragmites australis Trin ex Steud. wetland. Environ. Sci. Pollut. Res. 2017, 24, 12185–12194. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, C. Cadmium distribution and its effects on molybdate-containing hydroxylases in Phragmites australis. Aquat. Bot. 2007, 86, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Wang, C. Zinc distribution and zinc-binding forms in Phragmites australis under zinc pollution. J. Plant Physiol. 2008, 165, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, K.; Tsuchiya, M.; Nakata, S.; Tanabe, A.; Fukawa, S.; Kanai, M.; Miwa, E. Detoxification of cadmium (Cd) by a novel Cd-associated and Cd-induced molecule in the stem of common reed. J. Plant Physiol. 2013, 170, 1553–1560. [Google Scholar] [CrossRef]
- Christensen, K.K.; Sand-Jensen, K. Precipitated iron and manganese plaques restrict root uptake of phosphorus in Lobelia dortmanna. Can. J. Bot. 1998, 76, 2158–2163. [Google Scholar]
- Hansel, C.M.; Fendorf, S.; Sutton, S.; Newville, M. Characterization of Fe plaque and associated metals on the roots of mine-waste impacted aquatic plants. Environ. Sci. Technol. 2001, 35, 3863–3868. [Google Scholar] [CrossRef]
- Greipsson, S.; Crowder, A.A. Amelioration of copper and nickel toxicity by iron plaque on roots of rice (Oryza sativa). Can. J. Bot. 1992, 70, 824–830. [Google Scholar] [CrossRef]
- Tripathi, R.D.; Tripathi, P.; Dwivedi, S.; Kumar, A.; Mishra, A.; Chauhan, P.S.; Norton, G.J.; Nautiyal, C.S. Roles for root iron plaque in sequestration and uptake of heavy metals and metalloids in aquatic and wetland plants. Metallomics 2014, 6, 1789–1800. [Google Scholar] [CrossRef]
- Zaets, I.; Kozyrovska, N. Heavy metal resistance in plants: A putative role of endophytic bacteria. In Toxicity of Heavy Metals to Legumes and Bioremediation; Zaidi, A., Wani, P.A., Khan, M.S., Eds.; Springer: Vienna, Austria, 2012; pp. 203–217. [Google Scholar]
- Varma, A. Preface. In Microbial Siderophores; Varma, A., Chincholkar, S., Eds.; Springer: Heidelberg, Germany, 2007; pp. V–VI. [Google Scholar]
- Braud, A.; Jézéquel, K.; Léger, M.A.; Lebeau, T. Siderophore production by using free and immobilized cells of two pseudomonads cultivated in a medium enriched with Fe and/or toxic metals (Cr, Hg, Pb). Biotechnol. Bioeng. 2006, 94, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Whiting, S.N.; de Souza, M.P.; Terry, N. Rhizosphere bacteria mobilize Zn for hyperaccumulation by Thlaspi caerulescens. Environ. Sci. Technol. 2001, 35, 3144–3150. [Google Scholar] [CrossRef]
- Babu, A.G.; Kim, J.D.; Oh, B.T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thu-ringiensis GDB-1. J. Hazard. Mater. 2013, 250, 477–483. [Google Scholar] [CrossRef]
- Nagata, S.; Yamaji, K.; Nomura, N.; Ishimoto, H. Root endophytes enhance stress-tolerance of Cicuta virosa L. growing in a mining pond of eastern Japan. Plant Species Biol. 2014, 30, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Mikutta, C. X-ray absorption spectroscopy study on the effect of hydroxybenzoic acids on the formation and structure of ferrihydrite. Geochim. Cosmochim. Acta 2011, 75, 5122–5139. [Google Scholar] [CrossRef]
- Das, A.; Prasad, R.; Srivastava, A.; Giang, P.H.; Bhatnagar, K.; Varma, A. Fungal siderophores: Structure, functions and regulation. In Microbial Siderophores; Varma, A., Chincholkar, S.B., Eds.; Springer: Heidelberg, Germany, 2007; pp. 8–9. [Google Scholar]
- Bijlsma, J.; de Bruijn, W.J.C.; Hageman, J.A.; Goos, P.; Velikov, K.P.; Vincken, J.P. Revealing the main factors and two-way interactions contributing to food discolouration caused by iron-catechol complexation. Sci. Rep. 2020, 10, 8288. [Google Scholar] [CrossRef]
- Ejima, H.; Richardson, J.J.; Caruso, F. Metal-phenolic networks as a versatile platform to engineer nanomaterials and bio-interfaces. Nano Today 2017, 12, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Education, Culture, Sports, Science and Technology. No.14 Uranium Analysis, 2nd ed.; Japan Chemical Analysis Center: Chiba, Japan, 2002. (In Japanese)
- Mendez, M.O.; Maier, R.M. Phytostabilization of mine tailings in arid and semiarid environments—An emerging remediation technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, M.; Singh, S.P. A comparative study of cadmium phytoextraction by accumulator and weed species. Environ. Pollut. 2005, 133, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Stacey, M.G.; Patel, A.; McClain, W.E.; Mathieu, M.; Remley, M.; Rogers, E.E.; Gassmann, W.; Blevins, D.G.; Stacey, G. The Arabidopsis AtOPT3 protein functions in metal homeostasis and movement of iron to developing seeds. Plant Physiol. 2008, 146, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Alexander, D.B.; Zuberer, D.A. Use of chrome azurol S reagents to evaluate siderophore production by rhizosphere bacteria. Biol. Fertil. Soils 1991, 12, 39–45. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Fukuda, A.; Hagiwara, H.; Ishimura, T.; Kouduka, M.; Ioka, S.; Amano, Y.; Tsunogai, U.; Suzuki, Y.; Mizuno, T. Geomicrobiological properties of ultra-deep granitic groundwater from the Mizunami underground research laboratory (MIU), central Japan. Microb. Ecol. 2010, 60, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microb. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buyer, J.S.; Sikora, L.J.; Chaney, R.L. A new growth medium for the study of siderophore-mediated interactions. Biol. Fertil. Soils 1989, 8, 97–101. [Google Scholar] [CrossRef]
- Yamaji, K.; Ichihara, Y. The role of catechin and epicatechin in chemical defense against damping-off fungi of current-year Fagus crenata seedlings in natural forest. For. Pathol. 2012, 42, 1–7. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 201–223. [Google Scholar]
- Ali, N.A.; Bernal, M.P.; Ater, M. Tolerance and bioaccumulation of cadmium by Phragmites australis grown in the presence of elevated concentrations of cadmium, copper, and zinc. Aquat. Bot. 2004, 80, 163–176. [Google Scholar] [CrossRef]
- Favas, P.J.C.; Pratas, J.; Mitra, S.; Sarkar, S.K.; Venkatachalam, P. Biogeochemistry of uranium in the soil-plant and water-plant systems in an old uranium mine. Sci. Total. Environ. 2016, 568, 350–368. [Google Scholar] [CrossRef]
- Otte, M.L.; Rozema, J.; Koster, L.; Haarsma, M.S.; Broekman, R.A. Iron plaque on roots of Aster tripolium L.: Interaction with zinc uptake. New Phytol. 1989, 111, 309–317. [Google Scholar] [CrossRef]
- Park, T.; Kim, W.I.; Kim, B.J.; Lee, H.; Choi, I.S.; Park, J.H.; Cho, W.K. Salt-induced, continuous deposition of supramolecular iron (III)–tannic acid complex. Langmuir 2018, 34, 12318–12323. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.; Park, E.; Lee, H. Plant-inspired pyrogallol-containing functional materials. Adv. Funct. Mater. 2019, 29, 1903022. [Google Scholar] [CrossRef]
- Saha, R.; Saha, N.; Donofrio, R.S.; Bestervelt, L.L. Microbial siderophores: A mini review. J. Basic Microbiol. 2013, 53, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Budzikiewicz, H. Siderophores of fluorescent pseudomonads. Z. Naturforsch. C 1997, 52, 713–720. [Google Scholar] [CrossRef]
- Teintze, M.; Hossain, M.B.; Barnes, C.L.; Leong, J.; Van der Helm, D. Structure of ferric pseudobactin: A siderophore from a plant growth promoting Pseudomonas. Biochemistry 1981, 20, 6446–6457. [Google Scholar] [CrossRef] [PubMed]
- Cox, C.D.; Rinehart, K.L., Jr.; Moore, M.L.; Cook, J.C., Jr. Pyochelin: Novel structure of an iron-chelating growth promoter for Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1981, 78, 4256–4260. [Google Scholar] [CrossRef] [Green Version]
- Mossialos, D.; Meyer, J.-M.; Budzikiewicz, H.; Wolff, U.; Koedam, N.; Baysse, C.; Anjaiah, V.; Cornelis, P. Quinolobactin, a new siderophore of Pseudomonas fluorescens ATCC 17400, the production of which is repressed by the cognate pyoverdine. Appl. Environ. Microbiol. 2000, 66, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Risse, D.; Beiderbeck, H.; Taraz, K.; Budzikiewicz, H.; Gustine, D. Corrugatin, a lipopeptide siderophore from Pseudomonas corrugata. Z. Naturforsch. C 1998, 53, 295–304. [Google Scholar] [CrossRef]
- Matthijs, S.; Budzikiewicz, H.; Schäfer, M.; Wathelet, B.; Cornelis, P. Ornicorrugatin, a new siderophore from Pseudomonas fluorescens AF76. Z. Naturforsch. C 2008, 63, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Anthoni, U.; Christophersen, C.; Nielsen, P.H.; Gram, L.; Petersen, B.O. Pseudomonine, an isoxazolidone with siderophoric activity from Pseudomonas fluorescens AH2 isolated from Lake Victorian Nile perch. J. Nat. Prod. 1995, 58, 1786–1789. [Google Scholar] [CrossRef]
- Rosconi, F.; Davyt, D.; Martínez, V.; Martínez, M.; Abin-Carriquiry, J.A.; Zane, H.; Butler, A.; de Souza, E.M.; Fabiano, E. Identification and structural characterization of serobactins, a suite of lipopeptide siderophores produced by the grass endophyte Herbaspirillum seropedicae. Environ. Microbiol. 2013, 15, 916–927. [Google Scholar] [CrossRef]
- Smith, M.J.; Shoolery, J.N.; Schwyn, B.; Holden, I.; Neilands, J.B. Rhizobactin, a structurally novel siderophore from Rhizobium meliloti. J. Am. Chem. Soc. 1985, 107, 1739–1743. [Google Scholar] [CrossRef]
- Dilworth, M.J.; Carson, K.C.; Giles, R.G.F.; Byrne, L.T.; Glenn, A.R. Rhizobium leguminosarum bv. viciae produces a novel cyclic trihydroxamate siderophore, vicibactin. Microbiology 1998, 144, 781–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, E.P.; Boghozian, R.; Little, J.L.; Lowman, D.W.; Chakraborty, R. Characterization of ‘Schizokinen’; a dihydroxamate-type siderophore produced by Rhizobium leguminosarum IARI 917. Biometals 2006, 19, 637–649. [Google Scholar] [CrossRef]
- Patel, H.N.; Chakraborty, R.N.; Desai, S.B. Isolation and partial characterization of phenolate siderophore from Rhizobium leguminosarum IARI 102. FEMS Microbiol. Lett. 1988, 56, 131–134. [Google Scholar] [CrossRef]
- Modi, M.; Shah, K.S.; Modi, V.V. Isolation and characterization of catechol-like siderophore from cowpea Rhizobium RA-1. Arch. Microbiol. 1985, 141, 156–158. [Google Scholar] [CrossRef]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Concentration of Elements | ||
|---|---|---|---|
| Fe (mg/L) | Mn (mg/L) | 238U (µg/L) | |
| 2017 * | 8.54 ± 4.42 | 0.55 ± 0.21 | 45.8 ± 26.1 |
| 2018 ** | 15.2 ± 0.80 | 1.21 ± 0.21 | 14.9 ± 2.72 |
| Year | Water Temperature (°C) | pH | ORP (mV) |
|---|---|---|---|
| 2017 * | 15.8 ± 3.66 | 6.69 ± 0.18 | 26.3 ± 35.1 |
| 2018 ** | 16.4 ± 3.27 | 6.79 ± 0.07 | 12.0 ± 68.1 |
| Elements | July, 2017 | September, 2017 | ||
| Stems/Nodal Roots | Leaves/Nodal Roots | Stems/Nodal Roots | Leaves/Nodal Roots | |
| Fe | 0.05 × 10−1 ± 0.01 × 10−1 | 0.01 ± 0.00 | 0.01 × 10−1 ± 0.01 × 10−1 | 0.03 × 10−1 ± 0.01 × 10−1 ** |
| Mn | 0.03 ± 0.00 | 0.21 ± 0.03 ** | 0.05 ± 0.01 | 0.22 ± 0.06 * |
| 238U | — | — | — | — |
| Elements | July, 2018 | September, 2018 | ||
| Stems/Nodal Roots | Leaves/Nodal Roots | Stems/Nodal Roots | Leaves/Nodal Roots | |
| Fe | 0.02 × 10−1 ± 0.01 × 10−1 | 0.04 × 10−1 ± 0.01 × 10−1 | 0.04× 10−1 ± 0.01×10−1 | 0.07 × 10−1 ± 0.01 × 10−1 * |
| Mn | 0.06 ± 0.01 | 0.19 ± 0.05 | 0.08 ± 0.03 | 0.53 ± 0.10 * |
| 238U | — | — | 0.04×10−1 ± 0.01×10−1 * | 0.01×10−1 ± 0.02 × 10−1 |
| Elements | July, 2017 | September, 2017 | ||||
| Nodal Roots | Stems | Leaves | Nodal Roots | Stems | Leaves | |
| Fe | 1770 ± 409 a | 7.28 ± 0.23 b | 19.7 ± 1.54 b | 6900 ± 1130 a | 7.64 ± 0.81 b | 21.1 ± 2.56 b |
| Mn | 7140 ± 1060 a | 224 ± 36.2 b | 1420 ± 152 b | 6060 ± 1860 a | 222 ± 40.5 b | 1060 ± 179 ab |
| 238U | 6610 ± 2740 | — | — | 3410 ± 1430 | — | — |
| Elements | July, 2018 | September, 2018 | ||||
| Nodal Roots | Stems | Leaves | Nodal Roots | Stems | Leaves | |
| Fe | 2800 ± 448 a | 4.61 ± 0.53 b | 10.7 ± 3.43 b | 1480 ± 71.7 a | 6.09 ± 1.18 b | 9.70 ± 0.49 b |
| Mn | 3270 ± 967 a | 163 ± 27.0 b | 468 ± 105 ab | 1970 ± 401 a | 146 ± 40.1 b | 906 ± 140 ab |
| 238U | 7030 ± 1070 | — | — | 2780 ± 411 a | 10.4 ± 2.16 b | 30.1 ± 6.84 b |
| Bacterial Isolates | Siderophore Production Activity * | Species Name |
|---|---|---|
| T1-66 | 3.16 ± 0.01 | Pseudomonas rhodesiae strain VTT E-031889 |
| T5-48 | 3.15 ± 0.03 | Herbaspirillum sp. ZX2 |
| T1-42 | 3.12 ± 0.04 | Pseudomonas putida strain S975 |
| N5-48 | 3.10 ± 0.05 | Pseudomonas rhodesiae strain VTT E-031889 |
| T4-20 | 3.04 ± 0.02 | Pseudomonas rhodesiae strain VTT E-031889 |
| N4-46 | 2.87 ± 0.04 | Rhizobium nepotum |
| N2-80 | 2.83 ± 0.05 | Herbaspirillum sp. AU2339 |
| T1-11 | 2.71 ± 0.09 | Pseudomonas extremaustralis strain OX0322 |
| T4-45 | 2.33 ± 0.27 | Herbaspirillum sp. Os43 |
| N1-76 | 2.17 ± 0.16 | Pseudomonas protegens strain FDAARGOS_307 |
| Isolates | Siderophore Production Activity * (Clear-Zone Diameter, mm) | Precipitation with Fe Ions ** | Phenolic Production *** |
|---|---|---|---|
| T1-66 | 37.7 ± 0.07 | + | + |
| T5-48 | 26.4 ± 1.10 | − | − |
| T1-42 | 25.3 ± 0.61 | + | − |
| N5-48 | 35.6 ± 0.34 | + | + |
| T4-20 | 33.4 ± 0.59 | + | + |
| N4-46 | 32.6 ± 0.15 | + | + |
| N2-80 | 27.3 ± 0.43 | − | − |
| T1-11 | 38.1 ± 0.13 | + | − |
| T4-45 | 25.6 ± 0.20 | − | − |
| N1-76 | 32.1 ± 0.66 | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamoto, Y.; Doyama, K.; Haruma, T.; Lu, X.; Tanaka, K.; Kozai, N.; Fukuyama, K.; Fukushima, S.; Ohara, Y.; Yamaji, K. Fe, Mn and 238U Accumulations in Phragmites australis Naturally Growing at the Mill Tailings Pond; Iron Plaque Formation Possibly Related to Root-Endophytic Bacteria Producing Siderophores. Minerals 2021, 11, 1337. https://doi.org/10.3390/min11121337
Nakamoto Y, Doyama K, Haruma T, Lu X, Tanaka K, Kozai N, Fukuyama K, Fukushima S, Ohara Y, Yamaji K. Fe, Mn and 238U Accumulations in Phragmites australis Naturally Growing at the Mill Tailings Pond; Iron Plaque Formation Possibly Related to Root-Endophytic Bacteria Producing Siderophores. Minerals. 2021; 11(12):1337. https://doi.org/10.3390/min11121337
Chicago/Turabian StyleNakamoto, Yukihiro, Kohei Doyama, Toshikatsu Haruma, Xingyan Lu, Kazuya Tanaka, Naofumi Kozai, Kenjin Fukuyama, Shigeru Fukushima, Yoshiyuki Ohara, and Keiko Yamaji. 2021. "Fe, Mn and 238U Accumulations in Phragmites australis Naturally Growing at the Mill Tailings Pond; Iron Plaque Formation Possibly Related to Root-Endophytic Bacteria Producing Siderophores" Minerals 11, no. 12: 1337. https://doi.org/10.3390/min11121337