
Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review


1
Faculty of Health Sciences, University of Maribor, Žitna ulica 15, 2000 Maribor, Slovenia
2
Chemicals Office of the Republic of Slovenia, Ajdovščina 4, 1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(12), 2550; https://doi.org/10.3390/microorganisms9122550
Submission received: 26 November 2021
/
Revised: 6 December 2021
/
Accepted: 8 December 2021
/
Published: 10 December 2021
(This article belongs to the Special Issue Multidrug-Resistant Bacteria in the Environment, Their Resistance and Transfer Mechanisms)
Abstract
:Disinfectants are used to reduce the concentration of pathogenic microorganisms to a safe level and help to prevent the transmission of infectious diseases. However, bacteria have a tremendous ability to respond to chemical stress caused by biocides, where overuse and improper use of disinfectants can be reflected in a reduced susceptibility of microorganisms. This review aims to describe whether mutations and thus decreased susceptibility to disinfectants occur in bacteria during disinfectant exposure. A systematic literature review following PRISMA guidelines was conducted with the databases PubMed, Science Direct and Web of Science. For the final analysis, 28 sources that remained of interest were included. Articles describing reduced susceptibility or the resistance of bacteria against seven different disinfectants were identified. The important deviation of the minimum inhibitory concentration was observed in multiple studies for disinfectants based on triclosan and chlorhexidine. A reduced susceptibility to disinfectants and potentially related problems with antibiotic resistance in clinically important bacterial strains are increasing. Since the use of disinfectants in the community is rising, it is clear that reasonable use of available and effective disinfectants is needed. It is necessary to develop and adopt strategies to control disinfectant resistance.
1. Introduction
Disinfectants, defined as biocides “Main group 1” [1], are an essential tool in combatting the spread of infectious diseases. When used properly and according to the instructions, disinfectants can help prevent pathogens’ transmission and spread, especially in nosocomial infections. With the rise of life-threatening infections with antibiotic-resistant bacteria and newly emerging viruses, the use of disinfectants and virucidal sanitizing agents has increased [2,3].
Disinfectants contain one or more biocidal active substances by which harmful organisms are chemically or biologically deterred, rendered harmless or destroyed [4]. In the healthcare sector, in addition to hand hygiene, the disinfection of surfaces is just as crucial to effectively protect patients, healthcare workers and visitors from the transmission of pathogens [5,6]. Biocidal products are also used in everyday hygiene, where consumers are offered a wide range of antibacterial cleaners, hygienic dishwashers, anti-sweat textiles, hygiene wipes and hand disinfectants. Most of them contain biocidal active substances in various amounts [7,8]. Although personal and household hygiene is often equated with antimicrobial products, regular handwashing without disinfectants is far more essential and sufficient [9]. However, overuse and improper use of disinfectants can accumulate to be reflected in disinfectant resistance, potentially changing our way of life, from compromising food security to threatening our healthcare systems [6].
Antimicrobial resistance has aroused great interest in the scientific and medical community in the case of antibiotics [10,11,12,13], but less interest has been paid to disinfectants, widely used, mainly in clinical settings [14,15]. Overuse and, more importantly, misuse of disinfectants may reduce the susceptibility of target organisms to clinically important antimicrobials due to cross-resistance and/or co-resistance mechanisms [16,17,18,19]. Therefore, it is necessary to pay more attention to the most broadly used disinfectants, i.e., human personal hygiene products, surface/material disinfectants, and algaecides [20,21]. It is advised to use comparable products without these biocidal agents to avoid unnecessary selection pressure of disinfectants on bacteria. Preferably, if a common handwashing agent can deactivate bacteria, there is an incentive to promote such use [22,23].
Decreased susceptibility or even resistance against disinfectants can occur due to various inside and outside cellular mechanisms resulting from bacterial phenotypic and genotypic adaptation [3]. Unlike antibiotics, the mode of action of disinfectants can be unspecific, targeting different processes or sites in bacterial cells and inflicting cell damage to multiple bacteria; hence resistance development is unlikely [24,25,26]. Nevertheless, bacterial resistance to disinfectants can be created by the mutation or amplification of an endogenous chromosomal gene, by acquiring resistant determinants on chromosomal genetic elements like plasmids, transposons, and integrons [23,27], or due to changes in cell envelope permeability, increased efflux pump expression and the specific mechanisms of phenotypic traits [28]. Another means of bacterial adaptation presents through disinfectant inactivation or neutralization [29] and biofilm formation.
Decreased susceptibility and acquired resistance to disinfectants has been documented [20,30,31,32], primarily against less reactive active ingredients, such as quaternary ammonium compounds (QAC), biguanides, phenols [2], benzalkonium chloride, triclosan (TCS; polychlorinated phenoxy phenol), chlorhexidine (CHX) [33], didecyl dimethyl ammonium chloride (DDAC) [34], tetracycline and chloramphenicol [35].
This review aims to describe whether mutations and thus decreased susceptibility occur in bacteria during long-term use and exposure to disinfectants.
2. Materials and Methods
2.1. Data Sources and Search Strategy
We performed a systematic review using the examination, analysis and synthesis of literature and the compilation method. We followed the PRISMA guidelines [36]. The search was performed using search terms in English: (susceptibility OR resistance) AND (disinfectants OR biocides) AND (bacteria OR microorganisms). A literature search was conducted with the databases PubMed, Science Direct and Web of Science. We used the following search limits: research papers published in English related to the research topic until June 2020. We used the same search terms, search limits, inclusion, and exclusion criteria in all the databases. Predetermined inclusion and exclusion criteria were applied as presented in Table 1.
2.2. Study Selection
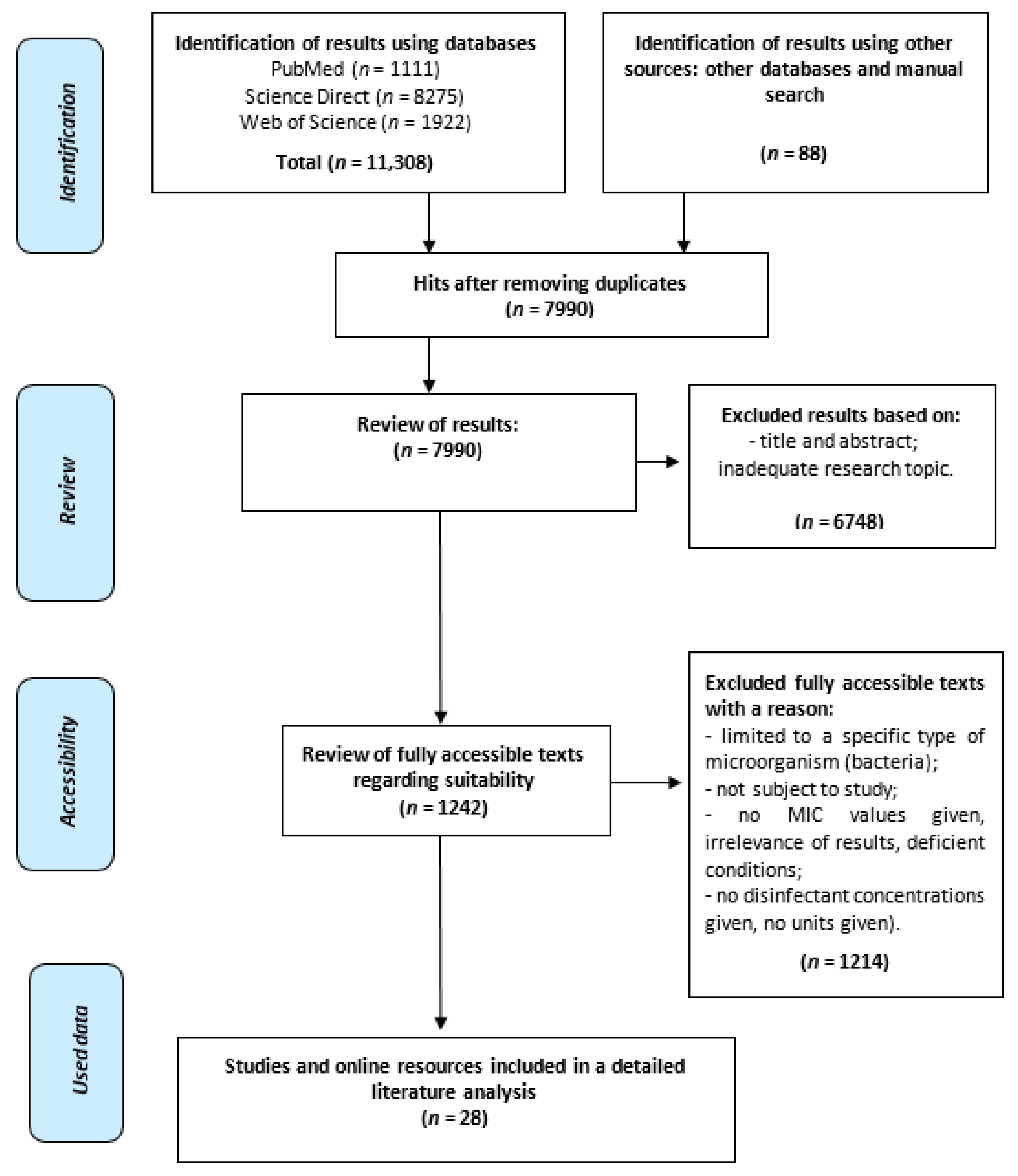
After revision of the databases, the results (n = 11,308) were exported and compiled with Mendeley’s reference management software. We also performed hand searching and included 88 articles. Mendeley’s automated process removed duplicates (n = 3318), followed by a manual search to identify and remove additional duplicates. The authors screened all abstracts (n = 7990). The search focused on articles describing bacteria that developed a substantial decrease in disinfectant susceptibility with known biocide ingredients. There were multiple reasons for excluding studies, mostly for lacking MIC values or disinfectant concentrations. For the final analysis and the review, 28 sources that remained of interest were included and screened based on their full text by two independent reviewers (See Figure 1).
2.3. Data Extraction and Analysis
The relevant data were first extracted by MP and checked by UR. The characteristics of the identified relevant sources were presented in a table, where we described the findings from the review and analysis of the relevant literature. The main findings from the identified sources were highlighted. The extracted data included: (1) disinfectant category; (2) publication author(s), year, country, journal; (3) study aim/purpose; (4) main results of the identified research study.
3. Results
Articles describing the resistance of bacteria against seven different disinfectants were identified, namely: triclosan (10 articles), peracetic acid (2 articles), hydrogen peroxide (3 articles), ethanol and isopropanol (1 article), formaldehyde and glutaraldehyde (2 articles), chlorhexidine (4 articles), benzalkonium chloride and didecyldimonium chloride (5 articles). For this review, the definition of bacterial resistance to a disinfectant is based on an importantly decreased susceptibility in different tests (e.g., disk diffusion test, minimum inhibitory concentration MIC) reported by the clinical microbiology or research laboratories described in the reviewed studies. All 28 selected publications that met the search criteria are classified in Table 2.
Multiple studies have shown an increased MIC from approximately 4- to 60-fold for specific bacteria for the disinfectant triclosan, making it epidemiologically relevant for increased bacterial adaptability and resistance [16,37,38,39,40,41,42,43,44,45]. The mechanisms for the elevated MIC were various mutations at the genetic level. For E. coli these were: deletion of the ycjD gene [45], mutation at codon 93 of the fabI gene, and mutation of the MarR transcription activator within the marRAB operon c, which regulates the operation of efflux pumps [41,43]. For P. aeruginosa, there was deletion of the fabV gene, leading to a decreased fatty acid synthesis and consequent inhibition of the production of acyl-homoserine lactones and other virulence factors, such as LasA/LasB, alkaline proteases, phospholipases, lipases, exotoxin A, rhamnolipid and pyocyanin, and a reduced pathogenicity [46]. It also affects the MexCDOprJ gene, PAO1, which encodes 12 RND pumps [47,48]. For S. aureus, intracellular malonyl-CoA inhibits the activity of the transcriptional repressor FapR, which directly interacts with the fabI gene, physiologically regulating its expression. This results in the most common mutations, polymorphisms, within the coding regions of C34T and MO035 in the sa-FabI region [49,50]. For S. enterica there was: a mutation in the fabI gene, mutation of the AcrAB and TolC genes that regulate efflux pumps, and inactivation of the transcriptional regulators ramA and marA [51,52,53].
For the disinfectant peracetic acid, the MIC was raised four-fold in one of the three bacteria tested (P. aeruginosa), while in the others, there were no significant changes in susceptibility. According to the described example, it could be classified as relevant in the indication of resistance, although the results are currently deficient due to the lack of multiple studies and unequal conditions [54,55]. For E. coli, mutations in the genes erm (B), tet (M), and tet (L) were observed [56].
For the disinfectant hydrogen peroxide, the MIC was also relevantly elevated in only one of the three bacteria studied (A. baumannii) and can be treated as a possible indicator of resistance here as well. However, due to deficient studies and unequal conditions, no conclusion regarding resistance can be made [57,58,59]. The cause of the elevated MIC were gene mutations. In all bacteria, mutations were in genes that regulate catalase (Kat), alkyl hydroperoxide reductase, and DNA-binding proteins that allow the catalase-reversible mechanism’s inhibitory effect on SpxB expression [60,61].
For the disinfectant chlorhexidine, the MIC has risen by almost 32–150 times in multiple relevant studies reviewed, making it epidemiologically relevant for increased bacterial adaptability and resistance, and a research/clinically relevant biocide [62,63,64,65,66]. The causes of the elevated MICs were gene mutations. For Enterococcus, mutation of the efrA and efrB genes that alter the expression of the EfrAB efflux pump of the ABC family, and hydrophobicity of the bacterial surface were observed [67,68]. For S. aureus and MRSA, mutations of qacA, qacB, smr and norA genes were observed [69,70,71,72]. For P. aeruginosa, mutation of efflux pump genes, such as MexCD-OprJ and oprH-phoPQ initiated by the stress response factor AlgU [69,70,71,72,73,74], and a decreased regulation of genes that encode proteins involved in membrane transport, oxidative phosphorylation, electron transport, and DNA repair were observed [75,76,77].
For the disinfectant benzalkonium chloride in multiple relevant studies reviewed, in three of the four bacteria, the MIC rises only 1–4-fold. This could make it epidemiologically relevant for increased bacterial adaptability and resistance, and a research/clinically relevant biocide [68,78,79,80,81]. The cause of the elevated MIC were gene mutations. For P. aeruginosa, mutations of efflux pump genes such as MDR mexA-mexB-oprM and mexC-mexD-oprJ were observed [82,83]. For E. coli, a mutation in the sugE gene located in the 94 regions of a chromosome that phenotypically inhibits a groEL mutation were observed [84,85]. For S. aureus, mutations of six different genes (i.e., qacA / B, qacC (smr), qacG, qacH and qacJ) that contribute to the development of resistance to QAC were observed [86,87,88,89].
For disinfectants containing alcohols, aldehydes and iodine compounds, no relevant changes of MIC values were reported [90,91,92,93,94].
The MIC values for most commonly used biocides against clinically important bacteria are presented in Table 3. The bacteria considered resistant had an increased MIC at least two times the average MIC in the first column.
4. Discussion
4.1. Criteria to Identify Resistant Strains
In order to understand resistance, there is an emphasis to distinguish between intrinsic and extrinsic resistance. Inherent resistance, known as natural resistance, is chromosomally encoded resistance, which determines the basic spectrum of effects of a disinfectant and the phenotypic resistance, e.g., biofilms. Extrinsic or acquired resistance develops through mutation by incorporating mobile genetic elements (horizontal gene transfer), transferable plasmids and other cell elements [114,115]. A clear distinction also needs to be made between phenotypic adaptation, which is reversible when exposure to the biocides ends, and acquired resistance, being genetically determined and usually stable [115]. When studying antibiotic resistance, the European Committee on Antimicrobial Susceptibility Testing has decided to: “define separate dividing points for the detection of bacteria with resistance mechanisms and the monitoring of resistance development using wild-type cut-off values (WCV) or epidemiological cut-off values (ECOFF or ECV) and the guidance of therapy via clinical breakpoints” [101,116]. As defined by the European Committee on Antimicrobial Susceptibility Testing, the ecological concept of antibiotic resistance states that ECOFFs are defined based on the normal distribution of MICs in a given bacterial species. Any isolate with a MIC above the epidemiological cut-off value (ECOFF), which is the upper limit of the normal distribution of the MIC for a given antimicrobial agent and a particular species, is considered resistant [43,96].
In the case of studying biocide resistance, however, no limits have been set so far, and there are no clear criteria to determine whether a microbe is susceptible to the biocide or not. Therefore, we can use the average MIC values obtained from individual laboratory studies conducted under relatively similar conditions. The observed relevant increase in the MIC value can indicate a decreased susceptibility or even resistance. When interpreting the results, the in-use concentration of the disinfectants used must be considered since the in-use concentration may also be higher than the actual measured MIC values. In this case, we cannot talk about the resistance but only about a decreased susceptibility.
4.2. Most Common Bacterial Mechanisms to Develop Resistance against Disinfectants
Bacteria control and overcome the effect of disinfectants in different ways (Table 4), such as restricted permeability of the cell wall, the expression of efflux systems, enzymatic degradation, changes in target sites, and the formation of biofilms [23,117]. Changes in cell surface hydrophobicity, ultrastructure, protein composition, and fatty acid modifications appear to occur [118,119]. For example, inactivation of the lipooligosaccharide biosynthesis genes causes resistance in A. baumannii [120]. Modifying the outer membrane proteins and an increased expression of cellular structures may increase the sensitivity to disinfectants [118,119,121]. Impermeability of the outer membrane occurs because of the lipopolysaccharide component, which increases the penetration of disinfectants and affects the size and expression of pores, thereby preventing entry and affecting sensitivity [122]. The hydrophilic porin channels on the outer membrane regulate the passage of solutes and are a significant barrier to hydrophilic substance penetration [123]. They also have a negative charge, which can cause the disinfectant molecules to bounce away from the bacterial cell.
Bacteria can also grow as biofilms, endospores, and within cellular macrophages. In most natural habitats, microorganisms grow and survive as associated biofilms [124]. Monocultures of several different species or mixed phenotypes of a particular species can form biofilms. It is a community of nonmobile microorganisms that are irreversibly attached to a surface and inserted into a polymeric extracellular matrix. The insensitivity of biofilms to disinfectants is due to altered microbial growth rates, which can be attributed to nutrient depletion in the biofilm, and disinfectant binding to the biofilm, which is neutralized or degraded [35]. Such an organization may moderate the concentration of antimicrobial disinfectants and antibiotics to which deeper biofilm cells are exposed. Such cells accidentally grow slowly, starve, and express stress phenotypes, including regulating efflux pumps and flushing out disinfectants [125].
Slightly less effective mechanisms involve the enzymatic degradation or inactivation of disinfectants when concentrations of agents, such as formaldehyde, chlorhexidine, and quaternary ammonium compounds, are lower than those used in clinical trials practice [126]. The exposure of bacteria to minimal inhibitory concentrations of disinfectants results in the induced expression of neutralizing enzymes, which is crucial for the biodegradation of disinfectants [127]. Examples of the neutralization of disinfectants have been given in several species of bacteria, for example, Pseudomonas fluorescens TN4 isolated from sludge was able to degrade DDAC, which belongs to the group of quaternary ammonium compounds. The isolate was also able to degrade other QACs by the N-dealkylation process [128].
One major cause of bacterial resistance is the active transport of substances to the cell exterior, the so-called efflux with proteins. Efflux pump mechanisms perform essential physiological functions [97]. Although existing in all living cells, those found in bacterial and mammalian cells are especially important for clinicians and pharmacologists since they constitute an important cause of antimicrobial resistance. Multidrug Resistance (MDR) efflux pumps present an ongoing research topic in antibiotic resistance and are also responsible for disinfectant resistance mechanisms [129]. One of the fundamental mechanisms of action is the efflux pump’s influence and the modulation of its genes. These efflux systems existed in bacteria long before the use of disinfectants and antibiotics in humans to treat infections. The mechanism involves the secretion of toxic compounds through a bacterial cell wall with a membrane-bound protein composed of at least three components. The increased expression of these pumps can raise the minimum inhibitory concentration to a high level, resulting in resistance to disinfectants [130] and greater sensitivity and cross-resistance to antibiotics [40]. Research data show that pump expression reduces the efficacy of various classes of disinfectants, including chlorhexidine digluconate, hydrogen peroxide, benzalkonium chloride, chloroxylenol, iodine compounds, triclosan, quaternary ammonium compounds, phenolic parabens and intercalates [131,132]. Among the best-studied systems of genes that regulate the secretion of biocides are mexAB-oprM, mexCD-oprJ and mexEF-oprN in P. aeruginosa [133], acrAB-tolC, acrEF-tolC and emrE in E. coli [134], smeDEF in bacteria Stenotrophomonas maltophilia [135], and norA and mepA in S. aureus. In the highly resistant nosocomial bacterium A. baumannii, the efflux activity is regulated by the quacA and quacB genes [57,77,136]. Bacteria use the same pumps to remove antibiotics and biocides. Thus, they can select antibiotic-resistant mutants that over-regulate such pumps [137].
Another important factor contributing to the development of disinfectant resistance is the mode of action of disinfectants. Biocides have a broader spectrum of activity and may have multiple targets, while antibiotics tend to have specific intracellular targets [29]. However, in the case of biocides with a particular antimicrobial mechanism (e.g., quaternary ammonium compounds—QAC’s), the development of antimicrobial resistance against disinfectants, and cross-resistance to antibiotics, are especially well documented [138].

{kind=link}
Table 4.
Disinfectant and bacterial mechanisms for the most commonly used disinfectants.
| Disinfectant Category | Active Ingredient | Use in Clinical Setting | Disinfectant Working Mechanism | Bacterial Adaptation to Disinfectant | Ref. |
|---|---|---|---|---|---|
| Alcohol | Ethic Alcohol (Ethanol) | 70–95% Ethanol solution | Denaturation of bacterial membrane proteins and dissolving lipid components such as antiparallel β and 310 helical turns of proteins, C-H deformations in lipids, inhibition of nutrient transport via membrane-bound ATPases, alteration of membrane pH and membrane potential. | Horizontal gene transfer, transformation and transduction and core genome mutations in the chromosome nucleotide position on the rpoB gene β subunit of RNA polymerase. | Alcohol working mechanism: [6,139,140,141] Alcohol adaptation: [91,142] |
| Aldehydes | Formaldehyde | 5% Formaldehyde solution | Cross-linking of protein’s free amino groups and inhibition of transport processes, RNA, and DNA. | Inactivation of formaldehyde through a metabolic system dependent on pterin cofactors, sugar phosphates, and those dependent on glutathione. Three separate enzymes catalyze successive stages of formaldehyde oxidation to CO2. These are the enzyme Gfa, alcohol dehydrogenase, and thioesterase. The specifically transmissible plasmid adhC gene encodes a glutathione-dependent formaldehyde dehydrogenase that causes inactivation. | Formaldehyde working mechanism: [3,29] Formaldehyde adaptation: [143,144,145] |
| Biguanides | Chlorhexidine (gluconate/diacetate) | 0.5% Alcohol solution (70%) | Inhibition of cytoplasmic membrane function and membrane-bound enzymes and leakage of intracellular components; inhibitor of both membrane-bound and soluble ATPase as well as of net K+ uptake, also collapses the membrane potential and has the potential for ATPase inactivation. | Induced gene expression of efflux pumps with upregulation and downregulation of coding genes (for an MFS transporter and HlyD-like periplasmic adaptor protein), active ingredient inactivation and alteration of the cell wall, increase in cell envelope components such as lipopolysaccharide or phospholipid caused by progressive mutations. | Chlorhexidine working mechanism: [29,33] Chlorhexidine adaptation: [29,33,43,65,146] |
| Bisphenol | Triclosan | Inhibition of enoyl-acyl carrier protein (ACP) reductase (FabI enzyme) in E. coli, P. aeruginosa, S. aureus and its homologue InhA in M. smegmatis, M. tuberculosis. | Target mutations, increased target expression (overexpressed genes mufA1 and mufM), active cell excretion, enzyme inactivation/ degradation. Increased concentration of branched chain fatty acids in the cell membrane occurs and multiple amino acids are changed in the fabI gene along with an increased concentration of the FabI protein through heterologous duplication and increased activity of ENR isoenzymes. | Triclosan working mechanism: [19,39,134,147,148] Triclosan adaptation: [30,149,150,151] | |
| Halogen releasing agents | Povidone-Iodine | 1–10% Iodine solution | Intracytoplasmic protein oxidation (cysteine and methionine), nucleotide and fatty acid function disruption, inhibition of production and release of bacterial exotoxins such as α-hemolysin, phospholipase C and enzymes such as elastase and β-glucuronide. | Formation of a biofilm and thickening of the cell wall. | Halogen releasing agents working mechanism: [29,152,153] Halogen releasing agents adaptation: [153,154] |
| Peroxygens | Hydrogen Peroxide | 3–6% Hydrogen Peroxide | H2O2 acts as an oxidant by producing hydroxyl or ferryl free radicals which disrupt the function of lipids, proteins-sulfhydryl (SH) and sulfur (SS) bonds and DNA. | Gene katA role of catalase, and peroxidase enzymes which neutralize H2O2. Bacterial cells form thick biofilm formations. | Hydrogen peroxide agents working mechanism: [29,155] Hydrogen peroxide agents adaptation: [155,156,157] |
| Quaternary Ammonium Compounds | Benzalkonium Chloride | 0.01–5% Benzalkonium chloride | Cationic amphiphilic properties destabilize the pathogen’s surface by forming electrostatic interactions with negatively charged components. Cytoplasmic membrane damage of phospholipid components occurs, distortion and protoplast lysis occur under osmotic stress. Leakage of low molecular weight components and eventual cell wall lysis. | Downregulation of membrane porins, overexpression or modification of efflux pumps (Mrdl EmrE MdfA) with mutations of the Mex system, horizontal gene transfer of transposon elements (Tn6188) and stress factors, biofilm formation, and biodegradation by dealkylation. | Benzalkonium chloride releasing agents working mechanism: [29,78,158] Benzalkonium chloride releasing agents adaptation: [82,159] |
5. Conclusions
Antimicrobial resistance in healthcare facilities has been occurring and regularly increasing over the last ten years. Growing evidence from in vitro studies has shown that bacteria have a tremendous ability to respond to chemical stress caused by biocides by several different mechanisms [160]. The main reason for emerging resistance is attributed mainly to the overuse, abuse and misuse of disinfectants [160,161,162]. Relevant increases in MIC concentrations, changes at the genetic level, and clearly altered mechanisms were observed in studies of several bacterial species in the presence of disinfectants. Through the most relevant of the reviewed articles, we can define the results for disinfectants based on triclosan and chlorhexidine, where the critical deviation of the MIC was observed in multiple studies.
Given the ongoing problems with multiple antibiotic resistance in clinically important bacteria strains and the potential for increased resistance to disinfectants, the use of which is rising in the community, it is clear that the prudent use of available and effective antimicrobials is needed. It is essential to develop and adopt strategies to control disinfectant resistance, for which the following factors will make a significant contribution. To solve the disinfectant resistance problem, it is essential to comprehensively summarize the disinfectant resistance mechanisms and to understand the resistance influencing factors [163]. It is also necessary to establish ECOFF values for biocides, without which any research is challenging and, to some extent, inaccurate. Harmonized methods for biocide susceptibility testing need to be developed.
Further studies are needed to establish a link between disinfectant exposure and resistance development, as many studies in clinical or external settings are currently limited. The rotation of disinfectants, where one disinfectant should be replaced by another having a different mechanism of action, is recommended [164]. The same types of disinfectants are used both in healthcare institutions and among the general population; therefore, their prudent use and consumption, as we know in the case of antibiotics, are complicated to control. Since selective pressure caused by disinfectants is exerted on both commensal and pathogenic bacteria [165,166], monitoring for resistant genes in nonpathogenic or commensal bacteria would make sense. Health-related infections acquired in the community need to be researched annually. More attention should be paid to the correct use of disinfectants by the general public, although supervising the proper use of disinfectants among the general population is very difficult to implement.
The risks and benefits of using disinfectants in the environment need to be weighed to determine whether additional precautions are required to guide the development and use of disinfectants [167]. If bacterial resistance increases and develops against many regularly used disinfectants in clinical and industrial settings, overuse in reflection of the COVID-19 pandemic could place an additional burden on global public health [168].
Author Contributions
U.R.: substantial contributions to conception and design, data acquisition, analysis and interpretation; drafting the article and revising it critically for important intellectual content; final approval of the version to be published. M.P.: substantial contributions to conception and design, data acquisition, analysis and interpretation; drafting the article. S.K.: substantial contributions to conception and design, critically revising the article. D.D.: substantial contributions to conception and design, data acquisition, analysis and interpretation; drafting the article and revising it critically for important intellectual content; final approval of the version to be published. S.Š.T.: substantial contributions to conception and design; final approval of the version to be published. All authors have read and agreed to the published version of the manuscript.
Funding
We are very grateful to Sanlas Holding GmbH, Austria who acted as a co-financer of the project. The research was also financially supported by the national research program (P2-0118).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- ECHA. Biocidal Products Regulation—Product Types. Available online: https://echa.europa.eu/regulations/biocidal-products-regulation/product-types (accessed on 10 February 2021).
- Maillard, J.-Y. Bacterial resistance to biocides in the healthcare environment: Should it be of genuine concern? J. Hosp. Infect. 2007, 65, 60–72. [Google Scholar] [CrossRef]
- Maillard, J.Y. Bacterial target sites for biocide action. J. Appl. Microbiol. 2002, 92, 16S–27S. [Google Scholar] [CrossRef] [PubMed]
- Pohl, K.; Dulio, V.; Botta, F.; Schwarzbauer, J.; Rüdel, H. Environmental monitoring of biocides in Europe: Compartment-specific strategies-Workshop report. In Proceedings of the Environmental Monitoring of Biocides in Europe: Compartment-Specific strategies, Berlin, Germany, 25–26 June 2015. [Google Scholar]
- Dumas, O.; Wiley, A.S.; Henneberger, P.K.; Speizer, F.E.; Zock, J.P.; Varraso, R.; Le Moual, N.; Boggs, K.M.; Camargo, C.A., Jr. Determinants of disinfectant use among nurses in US healthcare facilities. Am. J. Ind. Med. 2017, 60, 131–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutala, W.A.; Weber, D.J. Disinfection and sterilization in health care facilities: What clinicians need to know. Clin. Infect. Dis. 2004, 39, 702–709. [Google Scholar] [CrossRef] [Green Version]
- Hahn, S.; Schneider, K.; Gartiser, S.; Heger, W.; Mangelsdorf, I. Consumer exposure to biocides-identification of relevant sources and evaluation of possible health effects. Environ. Health 2010, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.; Lee, J.; Uhm, Y.; Kim, S.; Han, E.J.; Choi, K.; Choi, J.; Ban, Q.-m.; Cho, T.; Kim, A.Y. Korean consumers’ awareness of the risks of chemicals in daily consumer products. Environ. Sci. Eur. 2019, 31, 97. [Google Scholar] [CrossRef]
- Staniford, L.J.; Schmidtke, K.A. A systematic review of hand-hygiene and environmental-disinfection interventions in settings with children. BMC Public Health 2020, 20, 195. [Google Scholar] [CrossRef]
- Beyth, N.; Houri-Haddad, Y.; Domb, A.; Khan, W.; Hazan, R. Alternative antimicrobial approach: Nano-antimicrobial materials. Evid.-Based Complementary Altern. Med. 2015, 2015, 246012. [Google Scholar] [CrossRef] [Green Version]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.; Harper, D. Alternatives to antibiotics—A pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Rios, A.C.; Moutinho, C.G.; Pinto, F.C.; Del Fiol, F.S.; Jozala, A.; Chaud, M.V.; Vila, M.M.; Teixeira, J.A.; Balcão, V.M. Alternatives to overcoming bacterial resistances: State-of-the-art. Microbiol. Res. 2016, 191, 51–80. [Google Scholar] [CrossRef]
- Sakudo, A.; Yagyu, Y.; Onodera, T. Disinfection and sterilization using plasma technology: Fundamentals and future perspectives for biological applications. Int. J. Mol. Sci. 2019, 20, 5216. [Google Scholar] [CrossRef] [Green Version]
- Gebel, J.; Exner, M.; French, G.; Chartier, Y.; Christiansen, B.; Gemein, S.; Goroncy-Bermes, P.; Hartemann, P.; Heudorf, U.; Kramer, A. The role of surface disinfection in infection prevention. GMS Hyg. Infect. Control. 2013, 8, Doc10. [Google Scholar] [CrossRef]
- Gomes, I.; Malheiro, J.; Mergulhão, F.; Maillard, J.-Y.; Simões, M. Comparison of the efficacy of natural-based and synthetic biocides to disinfect silicone and stainless steel surfaces. Pathog. Dis. 2016, 74, ftw014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, S.; Latimer, J.; Bazaid, A.; McBain, A.J. Altered competitive fitness, antimicrobial susceptibility, and cellular morphology in a triclosan-induced small-colony variant of Staphylococcus aureus. Antimicrob. Agents Chemother. 2015, 59, 4809–4816. [Google Scholar] [CrossRef] [Green Version]
- Russell, A. Biocide use and antibiotic resistance: The relevance of laboratory findings to clinical and environmental situations. Lancet Infect. Dis. 2003, 3, 794–803. [Google Scholar] [CrossRef]
- Wieland, N.; Boss, J.; Lettmann, S.; Fritz, B.; Schwaiger, K.; Bauer, J.; Hölzel, C. Susceptibility to disinfectants in antimicrobial-resistant and-susceptible isolates of Escherichia coli, Enterococcus faecalis and Enterococcus faecium from poultry–ESBL/AmpC-phenotype of E. coli is not associated with resistance to a quaternary ammonium compound, DDAC. J. Appl. Microbiol. 2017, 122, 1508–1517. [Google Scholar]
- Yazdankhah, S.P.; Scheie, A.A.; Høiby, E.A.; Lunestad, B.-T.; Heir, E.; Fotland, T.Ø.; Naterstad, K.; Kruse, H. Triclosan and antimicrobial resistance in bacteria: An overview. Microb. Drug Resist. 2006, 12, 83–90. [Google Scholar] [CrossRef]
- European Commission. Assessment of the Antibiotic Resistance Effects of Biocides; European Commission: Brussels, Belgium, 2009. [Google Scholar]
- European Commission. Biocides Overview. Available online: https://ec.europa.eu/health/biocides/overview_en (accessed on 10 February 2021).
- Babeluk, R.; Jutz, S.; Mertlitz, S.; Matiasek, J.; Klaus, C. Hand hygiene–evaluation of three disinfectant hand sanitizers in a community setting. PLoS ONE 2014, 9, e111969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, J.S. Biocide resistance mechanisms. Int. Biodeterior. Biodegrad. 2003, 51, 133–138. [Google Scholar] [CrossRef]
- Goudarzi, M.; Navidinia, M. Overview perspective of bacterial strategies of resistance to biocides and antibiotics. Arch. Clin. Infect. Dis. 2019, 14, e65744. [Google Scholar]
- Romero, J.L.; Grande Burgos, M.J.; Pérez-Pulido, R.; Gálvez, A.; Lucas, R. Resistance to antibiotics, biocides, preservatives and metals in bacteria isolated from seafoods: Co-selection of strains resistant or tolerant to different classes of compounds. Front. Microbiol. 2017, 8, 1650. [Google Scholar] [CrossRef] [Green Version]
- Weber, D.J.; Rutala, W.A. Use of germicides in the home and the healthcare setting: Is there a relationship between germicide use and antibiotic resistance? Infect. Control. Hosp. Epidemiol. 2006, 27, 1107–1119. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Zeng, J.; Zhou, X.; Li, Y. Drug resistance and gene transfer mechanisms in respiratory/oral bacteria. J. Dent. Res. 2018, 97, 1092–1099. [Google Scholar] [CrossRef]
- Poole, K. Mechanisms of bacterial biocide and antibiotic resistance. J. Appl. Microbiol. 2002, 92, 55S–64S. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: Activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carey, D.E.; McNamara, P.J. The impact of triclosan on the spread of antibiotic resistance in the environment. Front. Microbiol. 2015, 5, 780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kampf, G. Biocidal agents used for disinfection can enhance antibiotic resistance in gram-negative species. Antibiotics 2018, 7, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wand, M.E.; Bock, L.J.; Bonney, L.C.; Sutton, J.M. Mechanisms of increased resistance to chlorhexidine and cross-resistance to colistin following exposure of Klebsiella pneumoniae clinical isolates to chlorhexidine. Antimicrob. Agents Chemother. 2017, 61, e01162-16. [Google Scholar] [CrossRef] [Green Version]
- Cieplik, F.; Jakubovics, N.S.; Buchalla, W.; Maisch, T.; Hellwig, E.; Al-Ahmad, A. Resistance toward chlorhexidine in oral bacteria—Is there cause for concern? Front. Microbiol. 2019, 10, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, S.E.; Maillard, J.-Y.; Russell, A.; Catrenich, C.; Charbonneau, D.; Bartolo, R. Development of bacterial resistance to several biocides and effects on antibiotic susceptibility. J. Hosp. Infect. 2003, 55, 98–107. [Google Scholar] [CrossRef]
- Gilbert, P.; Allison, D.; McBain, A. Biofilms in vitro and in vivo: Do singular mechanisms imply cross-resistance? J. Appl. Microbiol. 2002, 92, 98S–110S. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Bayston, R.; Ashraf, W.; Smith, T. Triclosan resistance in methicillin-resistant Staphylococcus aureus expressed as small colony variants: A novel mode of evasion of susceptibility to antiseptics. J. Antimicrob. Chemother. 2007, 59, 848–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, A.; Barbieri, R.; Read, R.; Church, D.; Adator, E.H.; Zaheer, R.; McAllister, T.A. Functional screening for triclosan resistance in a wastewater metagenome and isolates of Escherichia coli and Enterococcus spp. from a large Canadian healthcare region. PLoS ONE 2019, 14, e0211144. [Google Scholar] [CrossRef] [Green Version]
- Condell, O.; Sheridan, A.; Power, K.; Bonilla-Santiago, R.; Sergeant, K.; Renaut, J.; Burgess, C.; Fanning, S.; Nally, J. Comparative proteomic analysis of Salmonella tolerance to the biocide active agent triclosan. J. Proteom. 2012, 75, 4505–4519. [Google Scholar] [CrossRef]
- Copitch, J.L.; Whitehead, R.N.; Webber, M.A. Prevalence of decreased susceptibility to triclosan in Salmonella enterica isolates from animals and humans and association with multiple drug resistance. Int. J. Antimicrob. Agents 2010, 36, 247–251. [Google Scholar] [CrossRef]
- Cottell, A.; Denyer, S.; Hanlon, G.; Ochs, D.; Maillard, J.-Y. Triclosan-tolerant bacteria: Changes in susceptibility to antibiotics. J. Hosp. Infect. 2009, 72, 71–76. [Google Scholar] [CrossRef]
- Huang, Y.-H.; Lin, J.-S.; Ma, J.-C.; Wang, H.-H. Functional characterization of triclosan-resistant enoyl-acyl-carrier protein reductase (FabV) in Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 1903. [Google Scholar] [CrossRef] [Green Version]
- Levy, S. Active efflux, a common mechanism for biocide and antibiotic resistance. J. Appl. Microbiol. 2002, 92, 65S–71S. [Google Scholar] [CrossRef]
- Webber, M.A.; Randall, L.P.; Cooles, S.; Woodward, M.J.; Piddock, L.J. Triclosan resistance in Salmonella enterica serovar Typhimurium. J. Antimicrob. Chemother. 2008, 62, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.J.; Kim, J.A.; Pan, J.-G. Signature gene expression profile of triclosan-resistant Escherichia coli. J. Antimicrob. Chemother. 2010, 65, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef]
- Ellison, M.L.; Roberts, A.L.; Champlin, F.R. Susceptibility of compound 48/80-sensitized Pseudomonas aeruginosa to the hydrophobic biocide triclosan. FEMS Microbiol. Lett. 2007, 269, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lin, J.; Ma, J.; Cronan, J.E.; Wang, H. Triclosan resistance of Pseudomonas aeruginosa PAO1 is due to FabV, a triclosan-resistant enoyl-acyl carrier protein reductase. Antimicrob. Agents Chemother. 2010, 54, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Chuanchuen, R.; Karkhoff-Schweizer, R.R.; Schweizer, H.P. High-level triclosan resistance in Pseudomonas aeruginosa is solely a result of efflux. Am. J. Infect. Control. 2003, 31, 124–127. [Google Scholar] [CrossRef]
- D’Arezzo, S.; Lanini, S.; Puro, V.; Ippolito, G.; Visca, P. High-level tolerance to triclosan may play a role in Pseudomonas aeruginosa antibiotic resistance in immunocompromised hosts: Evidence from outbreak investigation. BMC Res. Notes 2012, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Ciusa, M.L.; Furi, L.; Knight, D.; Decorosi, F.; Fondi, M.; Raggi, C.; Coelho, J.R.; Aragones, L.; Moce, L.; Visa, P.; et al. A novel resistance mechanism to triclosan that suggests horizontal gene transfer and demonstrates a potential selective pressure for reduced biocide susceptibility in clinical strains of Staphylococcus aureus. Int. J. Antimicrob. Agents 2012, 40, 210–220. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, C.A.; Rybak, M.J. Efficacy of triclosan as an antimicrobial hand soap and its potential impact on antimicrobial resistance: A focused review. Pharmacotherapy 2015, 35, 328–336. [Google Scholar] [CrossRef]
- Nielsen, L.N.; Larsen, M.H.; Skovgaard, S.; Kastbjerg, V.; Westh, H.; Gram, L.; Ingmer, H. Staphylococcus aureus but not Listeria monocytogenes adapt to triclosan and adaptation correlates with increased fabI expression and agr deficiency. BMC Microbiol. 2013, 13, 177. [Google Scholar] [CrossRef] [Green Version]
- Spoering, A.L.; Lewis, K. Biofilms and planktonic cells of Pseudomonas aeruginosa have similar resistance to killing by antimicrobials. J. Bacteriol. 2001, 183, 6746–6751. [Google Scholar] [CrossRef] [Green Version]
- Van der Veen, S.; Abee, T. Mixed species biofilms of Listeria monocytogenes and Lactobacillus plantarum show enhanced resistance to benzalkonium chloride and peracetic acid. Int. J. Food Microbiol. 2011, 144, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Turolla, A.; Sabatino, R.; Fontaneto, D.; Eckert, E.M.; Colinas, N.; Corno, G.; Citterio, B.; Biavasco, F.; Antonelli, M.; Mauro, A. Defence strategies and antibiotic resistance gene abundance in enterococci under stress by exposure to low doses of peracetic acid. Chemosphere 2017, 185, 480–488. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Xu, Y.; Chang, Y.; Liu, C.; Jia, X.; Ling, B. Molecular characterization of reduced susceptibility to biocides in clinical isolates of Acinetobacter baumannii. Front. Microbiol. 2017, 8, 1836. [Google Scholar] [CrossRef]
- Pericone, C.D.; Overweg, K.; Hermans, P.W.; Weiser, J.N. Inhibitory and bactericidal effects of hydrogen peroxide production by Streptococcus pneumoniae on other inhabitants of the upper respiratory tract. Infect. Immun. 2000, 68, 3990–3997. [Google Scholar] [CrossRef] [Green Version]
- Wesgate, R.; Grasha, P.; Maillard, J.-Y. Use of a predictive protocol to measure the antimicrobial resistance risks associated with biocidal product usage. Am. J. Infect. Control. 2016, 44, 458–464. [Google Scholar] [CrossRef] [Green Version]
- Oogai, Y.; Kawada-Matsuo, M.; Komatsuzawa, H. Staphylococcus aureus SrrAB affects susceptibility to hydrogen peroxide and co-existence with Streptococcus sanguinis. PLoS ONE 2016, 11, e0159768. [Google Scholar] [CrossRef] [Green Version]
- Pesakhov, S.; Benisty, R.; Sikron, N.; Cohen, Z.; Gomelsky, P.; Khozin-Goldberg, I.; Dagan, R.; Porat, N. Effect of hydrogen peroxide production and the Fenton reaction on membrane composition of Streptococcus pneumoniae. Biochim. Biophys. Acta (BBA) Biomembr. 2007, 1768, 590–597. [Google Scholar] [CrossRef] [Green Version]
- Akinkunmi, E.O.; Lamikanra, A. Susceptibility of community associated methicillin resistant Staphylococcus aureus isolated from faeces to antiseptics. J. Infect. Dev. Ctries. 2011, 6, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Braga, T.M.; Pomba, C.; Lopes, M.F.S. High-level vancomycin resistant Enterococcus faecium related to humans and pigs found in dust from pig breeding facilities. Vet. Microbiol. 2013, 161, 344–349. [Google Scholar] [CrossRef]
- Kampf, G. Adaptive bacterial response to low level chlorhexidine exposure and its implications for hand hygiene. Microb. Cell 2019, 6, 307. [Google Scholar] [CrossRef]
- Thomas, L.; Maillard, J.-Y.; Lambert, R.; Russell, A. Development of resistance to chlorhexidine diacetate in Pseudomonas aeruginosa and the effect of a’residual’concentration. J. Hosp. Infect. 2000, 46, 297–303. [Google Scholar] [CrossRef]
- Valenzuela, A.S.; Benomar, N.; Abriouel, H.; Cañamero, M.M.; López, R.L.; Gálvez, A. Biocide and copper tolerance in enterococci from different sources. J. Food Prot. 2013, 76, 1806–1809. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, H.; Izutani, N.; Kitagawa, R.; Maezono, H.; Yamaguchi, M.; Imazato, S. Evolution of resistance to cationic biocides in Streptococcus mutans and Enterococcus faecalis. J. Dent. 2016, 47, 18–22. [Google Scholar] [CrossRef]
- Nasr, A.M.; Mostafa, M.S.; Arnaout, H.H.; Elshimy, A.A.A. The effect of exposure to sub-inhibitory concentrations of hypochlorite and quaternary ammonium compounds on antimicrobial susceptibility of Pseudomonas aeruginosa. Am. J. Infect. Control. 2018, 46, e57–e63. [Google Scholar] [CrossRef] [Green Version]
- Hasanvand, A.; Ghafourian, S.; Taherikalani, M.; Jalilian, F.A.; Sadeghifard, N.; Pakzad, I. Antiseptic resistance in methicillin sensitive and methicillin resistant Staphylococcus aureus isolates from some major hospitals, Iran. Recent Pat. Anti-Infect. Drug Discov. 2015, 10, 105–112. [Google Scholar] [CrossRef]
- Kernberger-Fischer, I.A. Nanosilberbeschichtete Lebensmittelverpackungsfolie zur Reduktion von Keimen auf Schweinefleisch und Testung der Empfindlichkeit von MRSA-und MSSA-Isolaten gegenüber Silber und sieben weiteren Bioziden. Ph.D. Thesis, Stiftung Tierärztliche Hochschule Hannover, Hannover, Germany, 2018. [Google Scholar]
- Noguchi, N.; Nakaminami, H.; Nishijima, S.; Kurokawa, I.; So, H.; Sasatsu, M. Antimicrobial agent of susceptibilities and antiseptic resistance gene distribution among methicillin-resistant Staphylococcus aureus isolates from patients with impetigo and staphylococcal scalded skin syndrome. J. Clin. Microbiol. 2006, 44, 2119–2125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, N.; Suwa, J.; Narui, K.; Sasatsu, M.; Ito, T.; Hiramatsu, K.; Song, J.-H. Susceptibilities to antiseptic agents and distribution of antiseptic-resistance genes qacA/B and smr of methicillin-resistant Staphylococcus aureus isolated in Asia during 1998 and 1999. J. Med. Microbiol. 2005, 54, 557–565. [Google Scholar] [CrossRef]
- Buxbaum, A.; Kratzer, C.; Graninger, W.; Georgopoulos, A. Antimicrobial and toxicological profile of the new biocide Akacid plus®. J. Antimicrob. Chemother. 2006, 58, 193–197. [Google Scholar] [CrossRef]
- Morita, Y.; Tomida, J.; Kawamura, Y. Responses of Pseudomonas aeruginosa to antimicrobials. Front. Microbiol. 2014, 4, 422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nde, C.W.; Jang, H.-J.; Toghrol, F.; Bentley, W.E. Global transcriptomic response of Pseudomonas aeruginosa to chlorhexidine diacetate. Environ. Sci. Technol. 2009, 43, 8406–8415. [Google Scholar] [CrossRef]
- Sheng, W.-H.; Wang, J.-T.; Lauderdale, T.-L.; Weng, C.-M.; Chen, D.; Chang, S.-C. Epidemiology and susceptibilities of methicillin-resistant Staphylococcus aureus in Taiwan: Emphasis on chlorhexidine susceptibility. Diagn. Microbiol. Infect. Dis. 2009, 63, 309–313. [Google Scholar] [CrossRef]
- Vijayakumar, R.; Sandle, T.; Al-Aboody, M.S.; AlFonaisan, M.K.; Alturaiki, W.; Mickymaray, S.; Premanathan, M.; Alsagaby, S.A. Distribution of biocide resistant genes and biocides susceptibility in multidrug-resistant Klebsiella pneumoniae, Pseudomonas aeruginosa and Acinetobacter baumannii—A first report from the Kingdom of Saudi Arabia. J. Infect. Public Health 2018, 11, 812–816. [Google Scholar] [CrossRef]
- Fazlara, A.; Ekhtelat, M. The disinfectant effects of benzalkonium chloride on some important foodborne pathogens. Am. Eurasian. J. Agric. Environ. Sci. 2012, 12, 23–29. [Google Scholar]
- He, G.-X.; Landry, M.; Chen, H.; Thorpe, C.; Walsh, D.; Varela, M.F.; Pan, H. Detection of benzalkonium chloride resistance in community environmental isolates of staphylococci. J. Med. Microbiol. 2014, 63, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mc Cay, P.H.; Ocampo-Sosa, A.A.; Fleming, G.T. Effect of subinhibitory concentrations of benzalkonium chloride on the competitiveness of Pseudomonas aeruginosa grown in continuous culture. Microbiology 2010, 156, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Jiang, X.; Zhang, Y.; Ji, S.; Gao, W.; Shi, L. Effect of benzalkonium chloride adaptation on sensitivity to antimicrobial agents and tolerance to environmental stresses in Listeria monocytogenes. Front. Microbiol. 2018, 9, 2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tandukar, M.; Oh, S.; Tezel, U.; Konstantinidis, K.T.; Pavlostathis, S.G. Long-term exposure to benzalkonium chloride disinfectants results in change of microbial community structure and increased antimicrobial resistance. Environ. Sci. Technol. 2013, 47, 9730–9738. [Google Scholar] [CrossRef]
- Wickham, G. An investigation into the relative resistances of common bacterial pathogens to quaternary ammonium cation disinfectants. Biosci. Horiz. Int. J. Stud. Res. 2017, 10, hzx008. [Google Scholar] [CrossRef] [Green Version]
- Chung, Y.J.; Saier, M.H. Overexpression of the Escherichia coli sugE gene confers resistance to a narrow range of quaternary ammonium compounds. J. Bacteriol. 2002, 184, 2543–2545. [Google Scholar] [CrossRef] [Green Version]
- Nishino, K.; Yamaguchi, A. Analysis of a complete library of putative drug transporter genes in Escherichia coli. J. Bacteriol. 2001, 183, 5803–5812. [Google Scholar] [CrossRef] [Green Version]
- Abdelaziz, A.; Sonbol, F.; Elbanna, T.; El-Ekhnawy, E. Exposure to Sublethal Concentrations of Benzalkonium Chloride Induces Antimicrobial Resistance and Cellular Changes in Klebsiellae pneumoniae Clinical Isolates. Microb. Drug Resist. 2019, 25, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.M.; Ishino, M.; Kobayashi, N. Analysis of genomic diversity and evolution of the low-level antiseptic resistance gene smr in Staphylococcus aureus. Microb. Drug Resist. 2003, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Correa, J.; De Paulis, A.; Predari, S.; Sordelli, D.; Jeric, P. First report of qacG, qacH and qacJ genes in Staphylococcus haemolyticus human clinical isolates. J. Antimicrob. Chemother. 2008, 62, 956–960. [Google Scholar] [CrossRef] [Green Version]
- Truong-Bolduc, Q.; Dunman, P.; Strahilevitz, J.; Projan, S.; Hooper, D. MgrA is a multiple regulator of two new efflux pumps in Staphylococcus aureus. J. Bacteriol. 2005, 187, 2395–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gradel, K.O.; Randall, L.; Sayers, A.R.; Davies, R.H. Possible associations between Salmonella persistence in poultry houses and resistance to commonly used disinfectants and a putative role of mar. Vet. Microbiol. 2005, 107, 127–138. [Google Scholar] [CrossRef]
- Pidot, S.J.; Gao, W.; Buultjens, A.H.; Monk, I.R.; Guerillot, R.; Carter, G.P.; Lee, J.Y.; Lam, M.M.; Grayson, M.L.; Ballard, S.A. Increasing tolerance of hospital Enterococcus faecium to handwash alcohols. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [Green Version]
- Reimer, K.; Wichelhaus, T.; Schäfer, V.; Rudolph, P.; Kramer, A.; Wutzler, P.; Ganzer, D.; Fleischer, W. Antimicrobial effectiveness of povidone-iodine and consequences for new application areas. Dermatology 2002, 204, 114–120. [Google Scholar] [CrossRef]
- Tschudin-Sutter, S.; Frei, R.; Kampf, G.; Tamm, M.; Pflimlin, E.; Battegay, M.; Widmer, A.F. Emergence of glutaraldehyde-resistant Pseudomonas aeruginosa. Infect. Control. Hosp. Epidemiol. 2011, 32, 1173–1178. [Google Scholar] [CrossRef] [Green Version]
- Willing, B.P.; Pepin, D.M.; Marcolla, C.S.; Forgie, A.J.; Diether, N.E.; Bourrie, B.C. Bacterial resistance to antibiotic alternatives: A wolf in sheep’s clothing? Anim. Front. 2018, 8, 39–47. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5564, Triclosan. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Triclosan (accessed on 19 February 2021).
- Morrissey, I.; Oggioni, M.R.; Knight, D.; Curiao, T.; Coque, T.; Kalkanci, A.; Martinez, J.L.; Consortium, B. Evaluation of epidemiological cut-off values indicates that biocide resistant subpopulations are uncommon in natural isolates of clinically-relevant microorganisms. PLoS ONE 2014, 9, e86669. [Google Scholar] [CrossRef] [Green Version]
- Webber, M.; Piddock, L. The importance of efflux pumps in bacterial antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 9–11. [Google Scholar] [CrossRef]
- Penna, T.C.V.; Mazzola, P.G.; Martins, A.M.S. The efficacy of chemical agents in cleaning and disinfection programs. BMC Infect. Dis. 2001, 1, 16. [Google Scholar] [CrossRef] [Green Version]
- Sella, S.R.; Guizelini, B.P.; Ribeiro, H. Validation of peracetic acid as a sporicide for sterilization of working surfaces in biological safety cabinets. J. Microbiol. Infect. Dis. 2012, 2, 93–99. [Google Scholar] [CrossRef]
- Maertens, H.; De Reu, K.; Meyer, E.; Van Coillie, E.; Dewulf, J. Limited association between disinfectant use and either antibiotic or disinfectant susceptibility of Escherichia coli in both poultry and pig husbandry. BMC Vet. Res. 2019, 15, 310. [Google Scholar] [CrossRef] [Green Version]
- Mazzola, P.G.; Jozala, A.F.; Novaes, L.C.d.L.; Moriel, P.; Penna, T.C.V. Minimal inhibitory concentration (MIC) determination of disinfectant and/or sterilizing agents. Braz. J. Pharm. Sci. 2009, 45, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, Q.; Lu, S.; Niu, Y. Hydrogen peroxide: A potential wound therapeutic target. Med. Princ. Pract. 2017, 26, 301–308. [Google Scholar] [CrossRef]
- Zubko, E.I.; Zubko, M.K. Co-operative inhibitory effects of hydrogen peroxide and iodine against bacterial and yeast species. BMC Res. Notes 2013, 6, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handtke, S.; Albrecht, D.; Zühlke, D.; Otto, A.; Becher, D.; Schweder, T.; Riedel, K.; Hecker, M.; Voigt, B. Bacillus pumilus KatX2 confers enhanced hydrogen peroxide resistance to a Bacillus subtilis P katA:: katX2 mutant strain. Microb. Cell Factories 2017, 16, 72. [Google Scholar] [CrossRef]
- D’Ercole, S.; Catamo, G.; De Fazio, P.; Piccolomini, R. In vitro antimicrobial activity of glutaraldehyde plus O-phenylphenol association (ratio 2:1). Minerva Stomatol. 2002, 51, 29–33. [Google Scholar]
- Gradel, K.O.; Randall, L. Minimum Inhibitory Concentrations of and Adaptation to Five Disinfectants Commonly Used Against Salmonella in the Poultry Industry. Available online: https://www.isah-soc.org/userfiles/downloads/proceedings/2003/speakers/S32GradelDenmark.pdf (accessed on 3 February 2021).
- Stuart, M.C.; Kouimtzi, M.; Hill, S.R. WHO Model Formulary 2008; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Vijayakumar, R.; Sandle, T. A review on biocide reduced susceptibility due to plasmid-borne antiseptic-resistant genes—special notes on pharmaceutical environmental isolates. J. Appl. Microbiol. 2019, 126, 1011–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CDC. Guideline for Disinfection and Sterilization in Healthcare Facilities, 2008. William A. Rutala, David J. Weber, Healthcare Infection Control Practices Advisory Committee (HICPAC). 2019. Available online: https://www.cdc.gov/infectioncontrol/pdf/guidelines/disinfection-guidelines-H.pdf (accessed on 3 February 2021).
- Ramzi, A.; Oumokhtar, B.; Ez Zoubi, Y.; Filali Mouatassem, T.; Benboubker, M.; El Ouali Lalami, A. Evaluation of Antibacterial Activity of Three Quaternary Ammonium Disinfectants on Different Germs Isolated from the Hospital Environment. BioMed Res. Int. 2020, 2020, 6509740. [Google Scholar] [CrossRef] [PubMed]
- Kouidhi, B.; Zmantar, T.; Jrah, H.; Souiden, Y.; Chaieb, K.; Mahdouani, K.; Bakhrouf, A. Antibacterial and resistance-modifying activities of thymoquinone against oral pathogens. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 29. [Google Scholar] [CrossRef] [Green Version]
- To, M.S.; Favrin, S.; Romanova, N.; Griffiths, M.W. Postadaptational resistance to benzalkonium chloride and subsequent physicochemical modifications of Listeria monocytogenes. Appl. Environ. Microbiol. 2002, 68, 5258–5264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langsrud, S.; Sidhu, M.S.; Heir, E.; Holck, A.L. Bacterial disinfectant resistance—A challenge for the food industry. Int. Biodeterior. Biodegrad. 2003, 51, 283–290. [Google Scholar] [CrossRef]
- Abreu, A.C.; Tavares, R.R.; Borges, A.; Mergulhão, F.; Simões, M. Current and emergent strategies for disinfection of hospital environments. J. Antimicrob. Chemother. 2013, 68, 2718–2732. [Google Scholar] [CrossRef]
- Meyer, B.; Cookson, B. Does microbial resistance or adaptation to biocides create a hazard in infection prevention and control? J. Hosp. Infect. 2010, 76, 200–205. [Google Scholar] [CrossRef]
- MIC EUCAST. MIC and Inhibition Zone Diameter Distributions of Microorganisms without and with Phenotypically Evident Resistance Mechanisms. Available online: https://mic.eucast.org (accessed on 3 February 2021).
- Lambert, P. Cellular impermeability and uptake of biocides and antibiotics in Gram-positive bacteria and mycobacteria. J. Appl. Microbiol. 2002, 92, 46S–54S. [Google Scholar] [CrossRef]
- Tattawasart, U.; Maillard, J.-Y.; Furr, J.; Russell, A. Outer membrane changes in Pseudomonas stutzeri resistant to chlorhexidine diacetate and cetylpyridinium chloride. Int. J. Antimicrob. Agents 2000, 16, 233–238. [Google Scholar] [CrossRef]
- Winder, C.; Al-Adham, I.; Abdel Malek, S.; Buultjens, T.; Horrocks, A.; Collier, P. Outer membrane protein shifts in biocide-resistant Pseudomonas aeruginosa PAO1. J. Appl. Microbiol. 2000, 89, 289–295. [Google Scholar] [CrossRef]
- Olmeda-López, H.; Corral-Lugo, A.; McConnell, M.J. Effect of Subinhibitory Concentrations of Antibiotics and Disinfectants on ISAba-Mediated Inactivation of Lipooligosaccharide Biosynthesis Genes in Acinetobacter baumannii. Antibiotics 2021, 10, 1259. [Google Scholar] [CrossRef]
- Dubois-Brissonnet, F.; Naïtali, M.; Mafu, A.A.; Briandet, R. Induction of fatty acid composition modifications and tolerance to biocides in Salmonella enterica serovar Typhimurium by plant-derived terpenes. Appl. Environ. Microbiol. 2011, 77, 906–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, C.; Pereira, A.; Pereira, M.; Melo, L.; Simões, M. Physiological changes induced by the quaternary ammonium compound benzyldimethyldodecylammonium chloride on Pseudomonas fluorescens. J. Antimicrob. Chemother. 2011, 66, 1036–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [Green Version]
- Leggett, M.J.; McDonnell, G.; Denyer, S.P.; Setlow, P.; Maillard, J.Y. Bacterial spore structures and their protective role in biocide resistance. J. Appl. Microbiol. 2012, 113, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Baugh, S.; Ekanayaka, A.S.; Piddock, L.J.; Webber, M.A. Loss of or inhibition of all multidrug resistance efflux pumps of Salmonella enterica serovar Typhimurium results in impaired ability to form a biofilm. J. Antimicrob. Chemother. 2012, 67, 2409–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheldon Jr, A.T. Antiseptic “resistance”: Real or perceived threat? Clin. Infect. Dis. 2005, 40, 1650–1656. [Google Scholar] [CrossRef]
- Gilbert, P.; McBain, A.J. Potential impact of increased use of biocides in consumer products on prevalence of antibiotic resistance. Clin. Microbiol. Rev. 2003, 16, 189–208. [Google Scholar] [CrossRef] [Green Version]
- Nishihara, T.; Okamoto, T.; Nishiyama, N. Biodegradation of didecyldimethylammonium chloride by Pseudomonas fluorescens TN4 isolated from activated sludge. J. Appl. Microbiol. 2000, 88, 641–647. [Google Scholar] [CrossRef]
- Jamshidi, S.; Sutton, J.M.; Rahman, K.M. An overview of bacterial efflux pumps and computational approaches to study efflux pump inhibitors. Future Med. Chem. 2016, 8, 195–210. [Google Scholar] [CrossRef]
- Gnanadhas, D.P.; Marathe, S.A.; Chakravortty, D. Biocides–resistance, cross-resistance mechanisms and assessment. Expert Opin. Investig. Drugs 2013, 22, 191–206. [Google Scholar] [CrossRef]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Randall, L.; Cooles, S.; Coldham, N.; Penuela, E.; Mott, A.; Woodward, M.J.; Piddock, L.; Webber, M. Commonly used farm disinfectants can select for mutant Salmonella enterica serovar Typhimurium with decreased susceptibility to biocides and antibiotics without compromising virulence. J. Antimicrob. Chemother. 2007, 60, 1273–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morita, Y.; Murata, T.; Mima, T.; Shiota, S.; Kuroda, T.; Mizushima, T.; Gotoh, N.; Nishino, T.; Tsuchiya, T. Induction of mexCD-oprJ operon for a multidrug efflux pump by disinfectants in wild-type Pseudomonas aeruginosa PAO1. J. Antimicrob. Chemother. 2003, 51, 991–994. [Google Scholar] [CrossRef] [Green Version]
- Braoudaki, M.; Hilton, A.C. Low level of cross-resistance between triclosan and antibiotics in Escherichia coli K-12 and E. coli O55 compared to E. coli O157. FEMS Microbiol. Lett. 2004, 235, 305–309. [Google Scholar]
- Sanchez, P.; Moreno, E.; Martinez, J.L. The biocide triclosan selects Stenotrophomonas maltophilia mutants that overproduce the SmeDEF multidrug efflux pump. Antimicrob. Agents Chemother. 2005, 49, 781–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, A.D. Mechanisms of bacterial insusceptibility to biocides. Am. J. Infect. Control 2001, 29, 259–261. [Google Scholar] [CrossRef]
- Huet, A.A.; Raygada, J.L.; Mendiratta, K.; Seo, S.M.; Kaatz, G.W. Multidrug efflux pump overexpression in Staphylococcus aureus after single and multiple in vitro exposures to biocides and dyes. Microbiology 2008, 154, 3144–3153. [Google Scholar] [CrossRef] [Green Version]
- Wessels, S.; Ingmer, H. Modes of action of three disinfectant active substances: A review. Regul. Toxicol. Pharmacol. 2013, 67, 456–467. [Google Scholar] [CrossRef]
- Aboualizadeh, E.; Bumah, V.V.; Masson-Meyers, D.S.; Eells, J.T.; Hirschmugl, C.J.; Enwemeka, C.S. Understanding the antimicrobial activity of selected disinfectants against methicillin-resistant Staphylococcus aureus (MRSA). PLoS ONE 2017, 12, e0186375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, I.; Somerville, G.A.; Heilmann, C.; Sahl, H.-G.; Maurer, H.H.; Herrmann, M. Very low ethanol concentrations affect the viability and growth recovery in post-stationary-phase Staphylococcus aureus populations. Appl. Environ. Microbiol. 2006, 72, 2627–2636. [Google Scholar] [CrossRef] [Green Version]
- Rutala, W.A.; Weber, D.J. Disinfection and Sterilization in Health Care Facilities: An Overview and Current Issues. Infect. Dis. Clin. N. Am. 2016, 30, 609–637. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7134755/pdf/main.pdf (accessed on 3 February 2021). [CrossRef]
- Borana, R.; Bari, S. Ethanol tolerance in Escherichia coli DH5-Alpha developed by serial exposure to sublethal doses is conferred to wild strains by horizontal gene transfer. bioRxiv 2020. [Google Scholar] [CrossRef]
- Chen, N.H.; Djoko, K.Y.; Veyrier, F.J.; McEwan, A.G. Formaldehyde stress responses in bacterial pathogens. Front. Microbiol. 2016, 7, 257. [Google Scholar] [CrossRef] [Green Version]
- Cloete, T.E. Resistance mechanisms of bacteria to antimicrobial compounds. Int. Biodeterior. Biodegrad. 2003, 51, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.; Lai, Y.; Hartwell, H.J.; Moeller, B.C.; Doyle-Eisele, M.; Kracko, D.; Bodnar, W.M.; Starr, T.B.; Swenberg, J.A. Formation, accumulation, and hydrolysis of endogenous and exogenous formaldehyde-induced DNA damage. Toxicol. Sci. 2015, 146, 170–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coenye, T.; Van Acker, H.; Peeters, E.; Sass, A.; Buroni, S.; Riccardi, G.; Mahenthiralingam, E. Molecular mechanisms of chlorhexidine tolerance in Burkholderia cenocepacia biofilms. Antimicrob. Agents Chemother. 2011, 55, 1912–1919. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.J.; Li, J.; Roland, G.E.; Rock, C.O. Inhibition of the Staphylococcus aureus NADPH-dependent enoyl-acyl carrier protein reductase by triclosan and hexachlorophene. J. Biol. Chem. 2000, 275, 4654–4659. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Jin, M.; Nguyen, S.H.; Mao, L.; Li, J.; Coin, L.J.; Yuan, Z.; Guo, J. Non-antibiotic antimicrobial triclosan induces multiple antibiotic resistance through genetic mutation. Environ. Int. 2018, 118, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.; Roy, N.; Choi, K.; Lee, S.-W. Distribution of triclosan-resistant genes in major pathogenic microorganisms revealed by metagenome and genome-wide analysis. PLoS ONE 2018, 13, e0192277. [Google Scholar] [CrossRef] [Green Version]
- Pycke, B.F.; Crabbé, A.; Verstraete, W.; Leys, N. Characterization of triclosan-resistant mutants reveals multiple antimicrobial resistance mechanisms in Rhodospirillum rubrum S1H. Appl. Environ. Microbiol. 2010, 76, 3116–3123. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, H.P. Triclosan: A widely used biocide and its link to antibiotics. FEMS Microbiol. Lett. 2001, 202, 1–7. [Google Scholar] [CrossRef]
- Bigliardi, P.L.; Alsagoff, S.A.L.; El-Kafrawi, H.Y.; Pyon, J.-K.; Wa, C.T.C.; Villa, M.A. Povidone iodine in wound healing: A review of current concepts and practices. Int. J. Surg. 2017, 44, 260–268. [Google Scholar] [CrossRef]
- Lachapelle, J.-M.; Castel, O.; Casado, A.F.; Leroy, B.; Micali, G.; Tennstedt, D.; Lambert, J. Antiseptics in the era of bacterial resistance: A focus on povidone iodine. Clin. Pract. 2013, 10, 579. [Google Scholar] [CrossRef]
- Capriotti, K.; Pelletier, J.; Barone, S.; Capriotti, J. Efficacy of dilute povidone-iodine against multi-drug resistant bacterial biofilms, fungal biofilms and fungal spores. Clin. Res. Dermatol. 2018, 5, 1–5. [Google Scholar]
- Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.-Y. Use of hydrogen peroxide as a biocide: New consideration of its mechanisms of biocidal action. J. Antimicrob. Chemother. 2012, 67, 1589–1596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baureder, M.; Reimann, R.; Hederstedt, L. Contribution of catalase to hydrogen peroxide resistance in Enterococcus faecalis. FEMS Microbiol. Lett. 2012, 331, 160–164. [Google Scholar] [CrossRef]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Bondurant, S.; McKinney, T.; Bondurant, L.; Fitzpatrick, L. Evaluation of a benzalkonium chloride hand sanitizer in reducing transient Staphylococcus aureus bacterial skin contamination in health care workers. Am. J. Infect. Control. 2020, 48, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Gravel, J.; Paradis-Bleau, C.; Schmitzer, A.R. Adaptation of a bacterial membrane permeabilization assay for quantitative evaluation of benzalkonium chloride as a membrane-disrupting agent. Medchemcomm 2017, 8, 1408–1413. [Google Scholar] [CrossRef] [PubMed]
- Mc Carlie, S.; Boucher, C.E.; Bragg, R.R. Molecular basis of bacterial disinfectant resistance. Drug Resist. Updates 2020, 48, 100672. [Google Scholar] [CrossRef] [PubMed]
- Bragg, R.; Jansen, A.; Coetzee, M.; van der Westhuizen, W.; Boucher, C. Bacterial resistance to Quaternary Ammonium Compounds (QAC) disinfectants. Adv. Exp. Med. Biol. 2014, 808, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Roca, I.; Akova, M.; Baquero, F.; Carlet, J.; Cavaleri, M.; Coenen, S.; Cohen, J.; Findlay, D.; Gyssens, I.; Heuer, O.E.; et al. The global threat of antimicrobial resistance: Science for intervention. New Microbes New Infect. 2015, 6, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Tong, C.; Hu, H.; Chen, G.; Li, Z.; Li, A.; Zhang, J. Disinfectant resistance in bacteria: Mechanisms, spread, and resolution strategies. Environ. Res. 2021, 195, 110897. [Google Scholar] [CrossRef]
- Murtough, S.M.; Hiom, S.J.; Palmer, M.; Russell, A.D. Biocide rotation in the healthcare setting: Is there a case for policy implementation? J. Hosp. Infect. 2001, 48, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.A.; Sischo, W.M.; Jones, L.P.; Moore, D.A.; Ahmed, S.; Short, D.M.; Besser, T.E. Recent Emergence of Escherichia coli with Cephalosporin Resistance Conferred by blaCTX-M on Washington State Dairy Farms. Appl. Environ. Microbiol. 2015, 81, 4403–4410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A. Brock Biology of Microorganisms, 14th ed.; Pearson: London, UK, 2015. [Google Scholar]
- Hardy, K.; Sunnucks, K.; Gil, H.; Shabir, S.; Trampari, E.; Hawkey, P.; Webber, M. Increased usage of antiseptics is associated with reduced susceptibility in clinical isolates of Staphylococcus aureus. MBio 2018, 9, e00894-18. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J. Tackling Drug Resistant Infections Globally–Final Report and Recommendations; Government of the United Kingdom: London, UK, 2016.
Figure 1.
Flow diagram search strategy according to PRISMA recommendations.

Table 1.
Research strategy inclusion and exclusion criteria.
| Databases: | PubMed, Science Direct and Web of Science | |
|---|---|---|
| Inclusion Criteria | Exclusion Criteria | |
| Problem: |
|
|
| Intervention/ Treatment: |
|
|
| Outcome: |
|
|
| Types of research: |
|
|
| Search limits | ||
| Timeframe: | Until December 2020 | |
| Language: | English | |
Table 2.
Study characteristics of identified research studies about disinfectant resistance.
| Author, Year, Country, Journal | Study Aim/Purpose | Main Results of Identified Research Studies |
|---|---|---|
| Triclosan | ||
| Cottell, et al. (2009), Great Britain and Germany, Journal of Hospital Infection | Determining the minimum inhibitory concentrations of triclosan with broth- and agar-dilution methods. Using the British Society for Antimicrobial Chemotherapy guidelines, the antibiotic susceptibilities were determined. Exploring further linkages between triclosan exposure and the emergence or lack of bacterial antibiotic resistance. | Triclosan MICs were significantly higher for the mutant strains S. aureus T3 and E. coli TM3 compared with the parent strains E. coli ATCC 8739 and S. aureus NCIMB 9518. Significantly higher MIC was also observed in the triclosan-tolerant strain A. johnsonii RC compared to the sensitive counterpart. |
| Byung, et al. (2010), Great Britain, Journal of Antimicrobial Therapy | Studying defense mechanisms against triclosan in mutants derived from the E. coli strain, which carry the target triclosan-resistant enzyme, FabI (G93V). | The MIC of triclosan for the E. coli imp4231 FabI (G93V) mutant with different concentrations after 20 h of exposure resulted in 8 mg/L. This is approximately 400 times higher when compared with the parent strain, where the MIC was 0.02 mg /L. The highest MIC of triclosan (80 mg/L) was in E. coli IFN4. |
| Cameron, et al. (2019), India, PLoS One | Identifying the functional mechanisms of triclosan resistance in waste waters metagenomes and assessing the frequency of resistance in clinical isolates of E. coli and Enterococcus spp. | Three E. coli isolates had up to 64-fold higher triclosan MICs (2 to 8 mg/L) than the original MICs and were presumed ESBL producers. |
| D’Arezzo, et al. (2012), Italia, BMC Research Notes | Evaluating the antimicrobial activity of triclosan and chlorhexidine digluconate (also for two commercial disinfectants) against the epidemic strain of P. aeruginosa. | An extremely high level of triclosan resistance (MIC 2125 mg/L) was observed for the P. aeruginosa epidemic strain. The same strain was susceptible to chlorhexidine digluconate (MIC 12.5 mg/L). The epidemic strain survived for more than 120 h in the presence of 3400 mg/L (≈ 1.6 × MIC) of triclosan upon gradual adaptation. |
| Huang, et al. (2016), China, Frontiers of Microbiology | Demonstrating the functional characteristic of triclosan-resistant enoyl-acyl protein reductase carrier (FabV) in P. aeruginosa. | The plasmid-bearing strain P. aeruginosa PAO170 (1fabV) (pSRK-PI and pSRK-PV) was triclosan resistant; the triclosan MIC was above 2000 μg/mL. |
| Forbes, et al. (2015), Great Britain, Antimicrobial Agents and Chemotherapy | Antimicrobial triclosan susceptibility testing, cell mobility and morphology in small colonies of Staphylococci. | S. aureus produced small colony variants with decreased triclosan susceptibility (4/6 test bacteria) when exposed to triclosan. The susceptibility of strains P0 to P10 increased in MIC (4- to 31-fold) and MBC increased from 3- to 16-fold. ATCC 6538 R1 had a MIC of up to 31 mg/L. |
| Bayston, et al. (2007), Great Britain, Journal of Antimicrobial Chemotherapy | Determining the antimicrobial activity duration of triclosan-impregnated silicone. Reporting about the development of MRSA resistance during experimental exposure. | Two of the three MRSA strains showed impaired coagulase production and decreased deoxyribonuclease production. Triclosan MICs increased between 8- and 67-fold (MIC up to 4.0 mg/L). |
| Copitch, et al. (2010), Great Britain, International Journal of Antimicrobial Agents | Determining the reduced susceptibility to triclosan in the group of S. enterica isolates and to identify and describe the mechanisms of resistance. | The level of resistance to triclosan was generally low in S. enterica isolates (triclosan range MIC 0.25–4 mg/L). Increased efflux activity was observed in multidrug-resistant and triclosan-resistant strains when comparing them to the strains with a reduced susceptibility to triclosan alone. |
| Condell, et al. (2012), Ireland, Journal of Proteomics | In the study, the authors compared the proteomic profile of the susceptible serovar S. enterica Typhimurium with its isogenic triclosan tolerant strain to decode cellular mechanisms that promote biocide tolerance. | Changes in the proteome of Salmonella were observed when exposed to sublethal concentrations of triclosan, which gave insights into mechanisms for the response and tolerance. |
| Webber, et al. (2008), Great Britain, Journal of Antimicrobial Chemotherapy | The purpose of the study was to describe the mechanisms of triclosan resistance in S. enterica Typhimurium. | Three triclosan-resistant phenotypes were classified as low MIC (MIC <8 mg/L), medium MIC (MIC 16–32 mg/L), and high MIC mutants (MIC > 32 mg/L). The most resistant mutant was strain L702 with MIC 64 mg/L. |
| Peracetic acid | ||
| Van der Veen and Abee (2011), Netherlands, International Journal of Food Microbiology | Studying single and mixed-species biofilm formation (Listeria monocytogenes EGD-e and LR-991), with Lactobacillus plantarum WCFS1 as the secondary species. Determining resistance to benzalkonium chloride and peracetic acid. | Resistance to benzalkonium chloride is higher in single and mixed-species biofilms than in planktonic grown cells. After exposure for 15 min to 100 μg/mL, mixed-species biofilms are more resistant to benzalkonium chloride than single-species biofilms. The resistance against peracetic acid treatments (15 min to 100 μg/mL) is also higher in single and mixed-species biofilms than planktonic grown cells, but the differences are less pronounced. |
| Spoering and Lewis (2001), USA, Journal of Bacteriology | Studying biofilms of P. aeruginosa wild-type strain PAO1 and comparing its resistance against biocide when comparing it to planktonic cells. | When comparing biofilms to logarithmic-phase planktonic cells, biofilms were considerably tolerant to the biocide. On the other hand, stationary-phase planktonic cells were more tolerant of peracetic acid than biofilms. The MBC for all three populations was 400 g/mL. |
| Hydrogen Peroxide | ||
| Pericone, et al. (2000), USA, Infection and Immunology | To determine if Streptococcus pneumoniae and Haemophilus influenza produce substances (hydrogen peroxide) that inhibit the other bacteria’s growth. | H. influenzae was most susceptible to growth inhibition, and killing by H2O2 (MIC, 0.4 mM; MBC, 0.5 mM). N. meningitides was also relatively sensitive (MIC, 0.4 mM; MBC, 5.0 mM). M. catarrhalis was shown to be relatively insensitive (MIC, 1.1 mM; MBC, 160 mM) like Pneumococcus (MIC, 1.6 mM; MBC, 80 mM), which could explain the ability to survive endogenously produced hydrogen peroxide. |
| Wesgate, et al. (2016), Great Britain, American Journal of Infection Control | Exposure of Staphylococcus aureus and Escherichia coli to triclosan, chlorhexidine gluconate solution, hydrogen peroxide and a hydrogen peroxide-based product. Determination of the susceptibility by combining standard efficacy protocols. | Significant increases in antimicrobial insusceptibility (MIC 69-fold; MBC 74-fold increase) were observed when exposing S. aureus for 5 min to 0.0004% triclosan.). A more than 30-fold increase in MIC was observed after E. coli exposure to bisphenol. No changes in the susceptibility profile (less than two-fold) were observed when exposing bacteria to 0.00005% CHG, 0.001% H2O2 and 0.001% Oxy BAC. |
| Lin et al. (2017), China, Frontiers of Microbiology | Using a PCR reverse transcriptase method for the assessment of the association between efflux pump gene expression and a reduced sensitivity for triclosan, chlorhexidine, benzalkonium chloride, hydrogen peroxide and ethanol. | A variety of susceptibilities to biocides was observed by the tested isolates. MICs for triclosan ranged from 2–256 μg/mL, for chlorhexidine 8–128 μg/mL, for benzalkonium bromide 4–32 μg/mL, (1.6–13 mg/mL), for hydrogen peroxide 47–376 mM, and for ethanol (60–180 mg/mL) at 7.5–22.5% (vol/vol). |
| Ethanol and Isopropanol | ||
| Pidot, et al. (2018), Australia, Science Translational Medicine | In this study, the authors sought to compare the alcohol tolerance of 139 nosocomial E. faecium isolates obtained between 1997 and 2015. | Newer clinical isolates of E. faecium were more resistant to alcohol than their predecessors. Using a 70% isopropanol surface disinfectant, mutated E. faecium isolates were ten times more tolerant to disinfectant than isolates from decades ago. Strain ST796 had a reduced tolerance to isopropanol of 1.14 log10. Four hundred nucleotide positions mutated on two or more pairs of sequences. |
| Formaldehyde and Glutaraldehyde | ||
| Gradel, et al. (2005), Denmark, Veterinary Microbiology | The study’s main objective was to perform a preliminary examination to detect apparent differences between Salmonella serotypes and isolates, to link them with the resistance to disinfectants, for which there are extensive data regarding Danish fattening flocks. | In MICs of five disinfectants commonly used in the Danish or English poultry sector, few variations were observed. Most differences from the isolates having high MICs were determined when using formaldehyde, but only a few isolates differed from the high MIC isolates when using the other four disinfectants. |
| Tschudin-Sutter, et al. (2011), Switzerland, Infection Control and Hospital Epidemiology | Determining the effectiveness of the endoscope cleaning procedure with glutaraldehyde in a Danish hospital against P. aeruginosa strains. | In samples obtained by endoscopes, P. aeruginosa was detected. During the disinfection procedure, a disinfectant based on glutaraldehyde showed no activity against two Pseudomonas outbreak strains when used under standard conditions at the recommended concentration. After reviewing the medical chart, six patients with circulatory and lower respiratory infections had an epidemiological link to the Pseudomonas outbreak strain. The most resistant strain needed the use of concentrations almost three times higher to achieve the same microbicidal effects. |
| Chlorhexidine | ||
| Braga, et al. (2013), Portugal, Veterinary Microbiology | The isolation of Enterococcal strains from dust samples collected from Portuguese breeding pig establishments. Determining the sensitivity of strains to chlorhexidine, benzalkonium chloride, biguanide and QAC was studied. The presence of VRE in these samples was also investigated. | The maximum MIC value for benzalkonium chloride and chlorhexidine in VRE isolates was 4 µg/mL. This was also the highest MIC value for all the other 41 isolates. The exceptions were two vancomycin-intermediate isolates, with a MIC to chlorhexidine of 8 µg/mL. |
| Valenzuela, et al. (2013), Spain, Journal of Food Protection | Enterococci were isolated from different animal species and plant food wildflowers, animal infestations and clinical specimens. Determinations were made of the resistance incidence for biocides (quaternary ammonium compounds, bisphenol, biguanide) and copper sulphate. | Triclosan (250 mg/L) inhibited 98.16% of isolates. The greatest variability was observed for chlorhexidine (MICs from 2.5 to 2500 mg/L). For the inhibition of 74.57% of isolates from clinical samples, the required dose of chlorhexidine was 2500 mg/L. Inhibition of Enterococci by copper sulfate was observed in the range 4–16 mM. |
| Akinkunmi and Lamikanra (2012), Nigeria, Journal of Infection in Developing Countries | Examination of MRSA resistance to commonly used antibiotics and antiseptics in fecal sample isolates from children in the community | Among MRSA isolates, 68.8%, 75.0%, and 100% were more resistant to benzalkonium chloride, chlorhexidine, and cetrimide than S. aureus NCTC 6571. Among the methicillin-susceptible S. aureus isolates, 32.0%, 28.0%, and 56.0% were more resistant to benzalkonium chloride, chlorhexidine gluconate, and cetrimide than S. aureus NCTC 6571. MIC50 values for S. aureus were 8 mg/mL, 4 mg/mL and 32 mg/mL for benzalkonium chloride, chlorhexidine and cetrimide, respectively. |
| Thomas, et al. (2000), Great Britain, Journal of Hospital Infection | The aim was to investigate the effects of sub-MIC concentrations of CHX on gram-negative bacteria, particularly the P. aeruginosa strain, which is known to have an intrinsic resistance to CHX, and the susceptibility of CHX-resistant strains to antibiotics. | After the fourth subculture, growth occurred within 24 h with a further increase in the MIC in P. aeruginosa strains NCIMB 10421; the MIC was significantly increased from the original 10 µg/mL to more than 70 µg/mL. The significance of these findings is still unclear, as the concentration of CHX in clinical use is much higher than that at which the authors obtained resistance. |
| Benzalkonium chloride and didecyldimonium chloride | ||
| Nasr, et al. (2018), Egypt, American Journal of Infection Control | The study aimed to evaluate the effect of isolated Pseudomonas exposure, the susceptibility to antibiotics, and to subinhibitory concentrations of two disinfectants, didecyldimonium chloride and sodium hypochlorite. | The development of antibiotic and biocidal resistant Pseudomonas strains can occur when using concentrations of sodium hypochlorite and didecyldimonium chloride. This study emphasizes the need for strict adherence to standard hospital disinfection policies to achieve adequate prevention and control of healthcare-associated infections. The MICs for all isolates ranged from 0.01% to 0.02% for sodium hypochlorite and 0.012% for didecyldimonium chloride. |
| McCay, et al. (2010), Ireland, Microbiology | Determining whether exposure of the population of P. aeruginosa (NCIMB 10421) to higher BAC levels in long-term continuous culture would result in a cross-adaptation to antimicrobials. | A method to enrich a continuous culture of P. aeruginosa NCIMB 10421, the MIC 25 mg/L of benzalkonium chloride was added (D = 0.04 h–1.792 h). The derivative PA-29 (696 h) showed a >12-fold reduced susceptibility to the biocide, MIC > 350 mg/L. |
| Fazlara and Ekhtelat (2012), Iran, American-Eurasian Journal of Agriculture | Evaluation of the antibacterial effects of benzalkonium chloride commonly used in the food industry, on six major food-borne pathogens. | The benzalkonium chloride MIC and MBC ranged between 40 and 45 mg/L for E. coli. The most susceptible and resistant bacteria were L. monocytogenes and B. cereus (MIC 30 and 35 mg/L and MBC 140 and 160 mg/L, respectively) |
| He, et al. (2014), China, Journal of Medical Microbiology | Studying isolates of BAC-resistant Staphylococci from the community environment with isolation, identification, and detection of BAC resistance genes. | The analysis of resistance genes showed that 41 strains contained qacA/B, 30 strains qacC, 25 strains qacG, 16 strains qacH, and eight strains qacJ. Because the BAC biocide affects these genes, this indicates an associated resistance in Staphylococci. The maximum MIC value for 63 strains of S. aureus ranged up to 32 μg/mL for BAC. |
| Yu, et al. (2018), China, Frontiers in Microbiology | The study examined the effect of BAC adaptation on antimicrobial susceptibility and tolerance to environmental loads and the role of efflux pumps in the adaptation of L. monocytogenes. | In BAC adapted Listeria strains, 18 EtBr strains had a MIC of 200 μg/mL, in 5 strains the MIC was > 200 μg/mL and in 2 strains it was 100 μg/mL. |
| Ramzi, et al., (2020), Morocco, BioMed Research International | Studying the antibacterial activity of quaternary ammoniums synthetic disinfectants for hospital environment isolates: Escherichia coli, Klebsiella pneumoniae, Enterobacter cloacae, Pseudomonas aeruginosa, Acinetobacter baumannii, and Staphylococcus aureus. | The tested disinfectant demonstrated an antibacterial effect against S. aureus and S. aureus ATCC 29213 (MIC of 0.25 mg/mL); the disinfectant spray showed effects in E. coli, S. aureus, E. coli ATCC 25922, and P. aeruginosa ATCC 27853 (MIC of 4 mg/mL) and S. aureus ATCC 29213 (MIC 2 mg/mL). Phagosurf ND® inhibited the growth of S. aureus ATCC 29213 (MIC of 4 mg/mL). |
Table 3.
MIC of biocides and the expressed susceptibility/resistance of bacteria to disinfectants.
| Bacteria/Disinfectant | MIC—Average Cut Off Values from Studies (mg/L) (Number of Strains) | MIC—Outstanding Elevated Values (mg/L) (Strain type) | Study Data Quality and Limitations | Main Results from Identified Studies—Resistance Relevance/Rise of MIC |
|---|---|---|---|---|
| Triclosan (0.5–2%) [95] | ||||
| Escherichia coli | 2 mg/L (368 strains) [96] | 8 mg/L (E. coli imp4231 FabI (G93V)) [81] | / | Relevant MIC increase indicates resistance. |
| Escherichia coli | 2 mg/L (368 strains) [96] | 8 mg/L (multiple strains) [38] | / | Relevant MIC increase indicates resistance. |
| Escherichia coli | 2 mg/L (368 strains) [96] | 1000 mg/L (E. coli TM3) [41] | Lack of precise evidence, unclear standards and research conditions. | Relevant MIC increase does not indicate resistance due to low-quality study. |
| Pseudomonas aeruginosa | 1000–2000 mg/L (1–6 strains) [50] | 4250 mg/L (multiple strains) [50] | / | Relevant MIC increase indicates resistance. |
| Pseudomonas aeruginosa | 1000–2000 mg/L (1–6 strains) [50] | >2000 mg/L (P. aeruginosa PAO170 (1fabV)) [42] | Lack of precise MIC values, only information on a value higher than 2000 mg/L. | Relevant MIC increase can potentially indicate resistance, but there is insufficient data. |
| Staphylococcus aureus | 0.5 mg/L (1635 strains) [49] | 31 mg/L (S. aureus ATCC 6538 R1) [16] | / | Relevant MIC increase indicates resistance. |
| Methicillin resistant Staphylococcus aureus (MRSA) | 0.5 mg/L (1635 strains) [49] | 4 mg/L (MRSA F1855) [37] | Lack of data for MRSA strains. | Relevant MIC increase does not indicate resistance because of values only for S. aureus. |
| Salmonella enterica | 8 mg/L (901 strains) [47] | 0.25–4 mg/L (multiple strains) [40] | / | No relevant MIC increase does not indicate resistance. |
| Salmonella enterica | 8 mg/L (901 strains) [47] | No MIC values. Only an increase of 1000 × mentioned (multiple strains) [39] | Lack of more accurate numerical MIC values. | Lack of MIC values does not provide sufficient data indicating relevance for resistance. |
| Salmonella enterica | 8 mg/L (901 strains) [47] | 64 mg/L (S. enterica L702) [97] | / | Relevant MIC increase indicates resistance. |
| Peracetic acid (0.2–3%) [98,99] | ||||
| Pseudomonas aeruginosa | 100 mg/L (1 strain) [54] | 400 mg/L (P.aeruginosa PAO1) [54] | Lack of sufficient MIC data and test sets to determine relevance. | Relevant MIC increase can potentially indicate resistance, but there is insufficient data. |
| Bacillus atrophaeus and Escherichia coli | 1.25–5 mg/L in 3–6 mg/L (118 strains) [99,100] | MIC value 5 mg/L (multiple strains) (B. atrophaeus ATCC 9372) [99,100] | No MIC values in mg/L. | No relevant MIC increase does not indicate resistance. |
| Hydrogen peroxide (0.001–4%) [101,102,103] | ||||
| Staphylococcus aureus | 0.2 mM–938 mg/L (2 strains) [103] | No MIC values 0.300 mg/L (S. aureus NCIMB 9518) [59] | No MIC values in mg/L. | Lack of MIC values does not provide sufficient data indicating relevance for resistance. |
| Acinetobacter spp. | 47 mM–469 mg/L (48 strains) [57,101] | MIC value 13,000 mg/L (multiple strains) [57] | / | Relevant MIC increase indicates resistance. |
| Bacillus spp. | 0.2 mM–1,875 mg/L (3 strains) [101,104] | MIC value 2 mM (B. subtilis PkatA:katX2) [104] | / | Relevant MIC increase indicates resistance. |
| Alcohols (70%) [87,101] | ||||
| Enterococcus faecium Staphylococcus aureus | Od 43,750 mg/L do 87,500 mg/L (7 strains) [101] | No MIC values. The only number of bacteria reduced in log10 [91] | No MIC values in mg/L. | Lack of MIC values does not provide sufficient data indicating relevance for resistance. |
| Aldehydes (0.5–4.0%) [105,106] | ||||
| Salmonella enterica (Formaldehyde) | 0.3 mg/L (34 strains) [106] | 0.125 mg/L (multiple strains) [90] | / | No relevant MIC increase does not indicate resistance. |
| Pseudomonas aeruginosa (Glutaraldehyde) | 3750 mg/L (1 strain) [105] | No MIC values, only recognized increase by 3× (P. aeruginosa ATCC 15442) [93] | No sublethal MIC values and no MIC measurement units described. | Lack of MIC values does not provide sufficient data indicating relevance for resistance. |
| Chlorhexidine (0.5–4.0%) [107] | ||||
| Vancomycin-resistant Enterococcus (VRE) | 4–16 mg/L (5 strains) [1] | 8 mg/L (VRE VanA) [63] | / | No relevant MIC increase does not indicate resistance. |
| Methicillin resistant Staphylococcus aureus (MRSA) | 8–128 mg/L (282 strains) [69] | Only MIC50 and MIC90 values <32 mg/L (multiple strains) [62] | No MIC values, only MIC50 and MIC90. | Lack of MIC values does not provide sufficient data indicating relevance for resistance. |
| Enterococcus faecalis | 16 mg/L (56 strains) [96] | 2500 mg/L (multiple strains) [66] | / | Relevant MIC increase indicates resistance. |
| Enterococcus faecium | 32 mg/L (53 strains) [96] | 250 mg/L (multiple strains) [66] | / | Relevant MIC increase indicates resistance. |
| Pseudomonas aeruginosa | 8–64 mg/L (70 strains) [73,108] | 70 mg/L (P. aeruginosa NCIMB 10421) [65] | / | Relevant MIC increase indicates resistance. |
| Pseudomonas aeruginosa | 8–64 mg/L (70 strains) [73,108] | 1024 mg/L (P. aeruginosa NCTC 6749) [64] | / | Relevant MIC increase indicates resistance. |
| Benzalkonium chloride and didecyldimonium chloride (0.01–5%) [109] | ||||
| Pseudomonas spp. (Benzalkonium chloride) | 4–512 mg/L (11 strains) [77,110] | >350 mg/L (P. aeruginosa PA-29) [80] | / | No relevant MIC increase does not indicate resistance. |
| Bacillus spp. (Benzalkonium chloride) | 16 mg/L (1 strains) [96] | 140–160 mg/L (B. cereus ATCC 11778) [78] | Disinfectant concentration and disinfectant contact duration not provided. | Relevant MIC increase can potentially indicate resistance, but there is insufficient data. |
| Staphylococcus aureus (Benzalkonium chloride) | 16 mg/L (1635 strains) [96] 0.25–4 mg/L (2 strains) [111] | 32 mg/L (multiple strains) [79] | / | Relevant MIC increase indicates resistance. |
| Listeria monocytogenes (Benzalkonium chloride) | 4–10 mg/L (31 strains) [111,112] | 14 mg/L (multiple strains) [81] | / | Relevant MIC increase can potentially indicate resistance, although the MIC rise is minimal. |
| Pseudomonas spp. (Didecyldimonium chloride) | 5–40 mg/L (11 strains) [113] | 120 mg/L (multiple strains) [1] | / | Relevant MIC increase indicates resistance. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rozman, U.; Pušnik, M.; Kmetec, S.; Duh, D.; Šostar Turk, S. Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review. Microorganisms 2021, 9, 2550. https://doi.org/10.3390/microorganisms9122550
AMA Style
Rozman U, Pušnik M, Kmetec S, Duh D, Šostar Turk S. Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review. Microorganisms. 2021; 9(12):2550. https://doi.org/10.3390/microorganisms9122550
Chicago/Turabian StyleRozman, Urška, Marko Pušnik, Sergej Kmetec, Darja Duh, and Sonja Šostar Turk. 2021. "Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review" Microorganisms 9, no. 12: 2550. https://doi.org/10.3390/microorganisms9122550
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.