
Preservation of Underground Microbial Diversity in Ancient Subsurface Deposits (>6 Ma) of the Rio Tinto Basement

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Geological Settings
3. Materials and Methods
3.1. Sample Collection
3.2. Sample Preparation
3.3. ToF-SIMS Analysis
4. Results
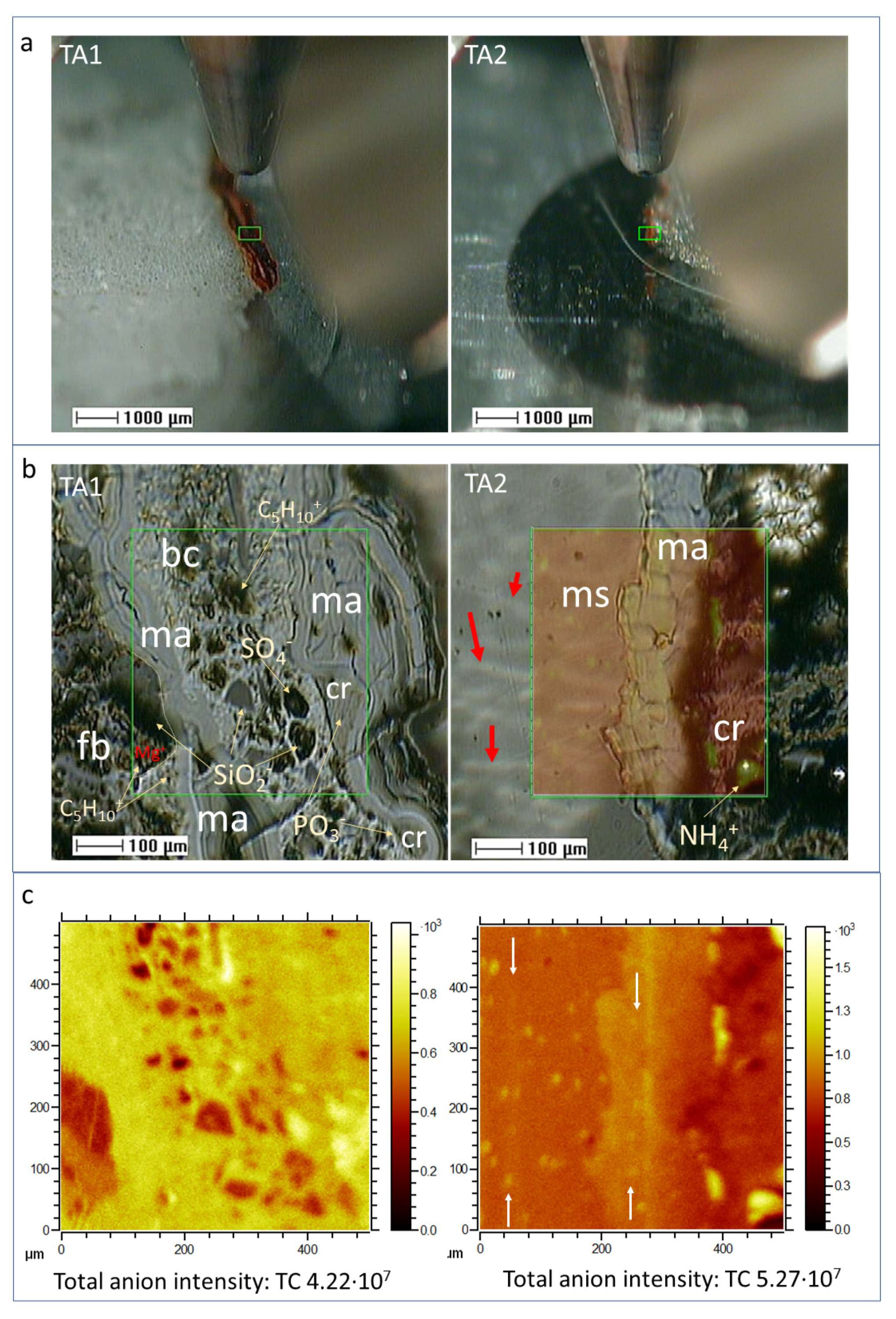
4.1. ToF-SIMS Image Analysis
4.1.1. Sample Microstructure
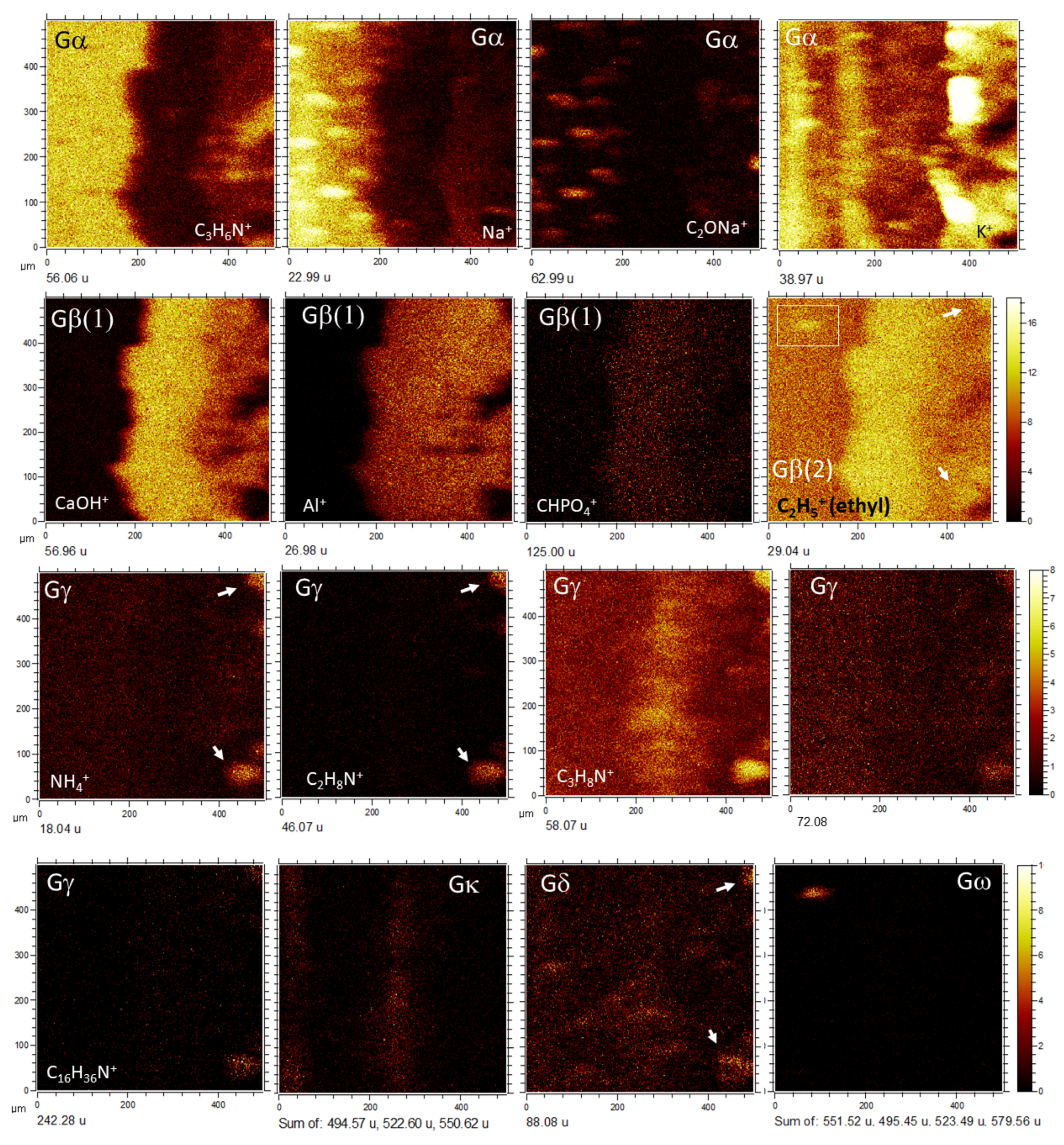
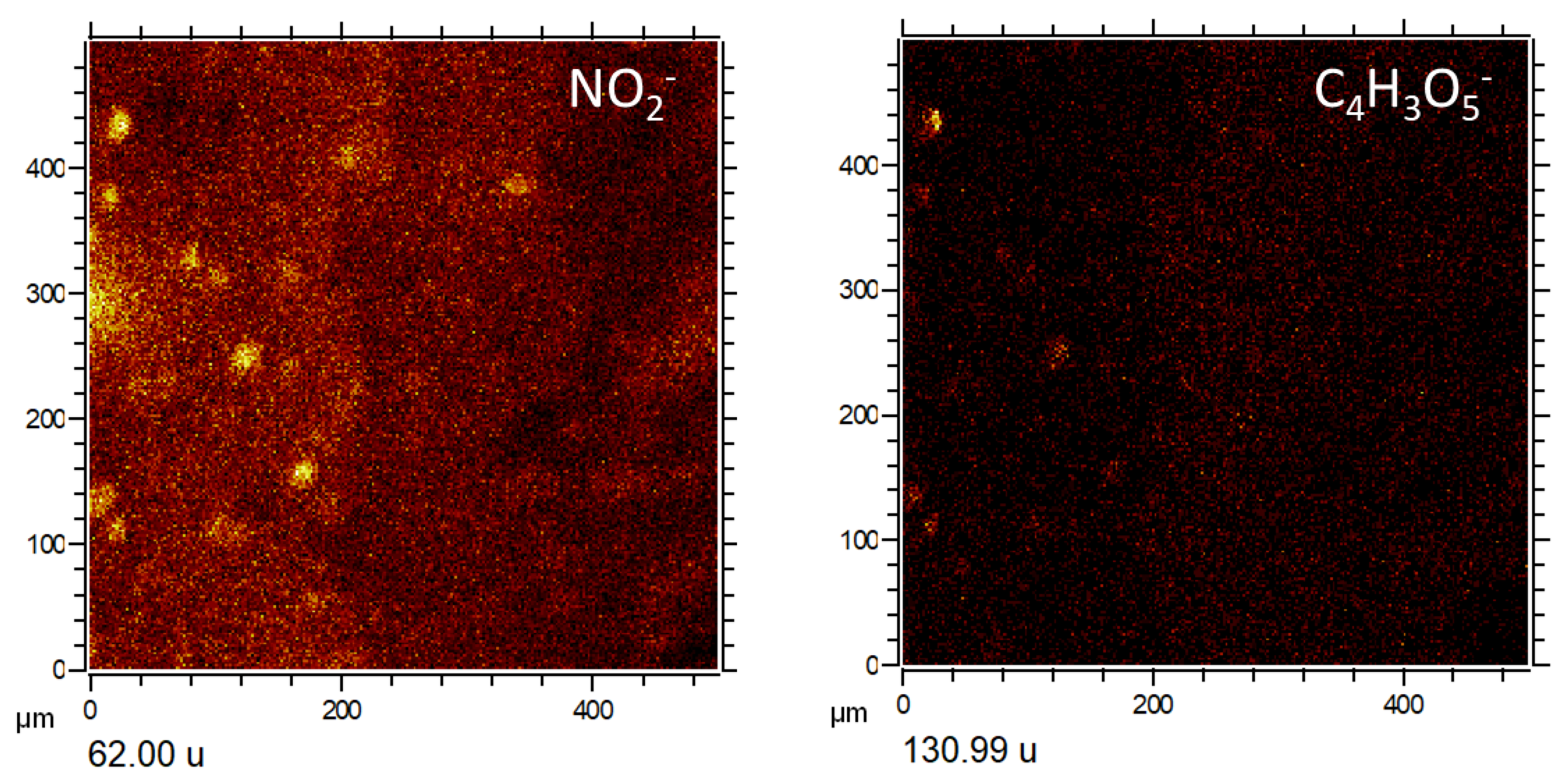
4.1.2. Distribution of Inorganic Ions
4.1.3. Morphological Groups
4.2. ToF-SIMS Spectral Identification
4.2.1. Hydrocarbons
4.2.2. Polycyclic Aromatic Hydrocarbons (PAHs)
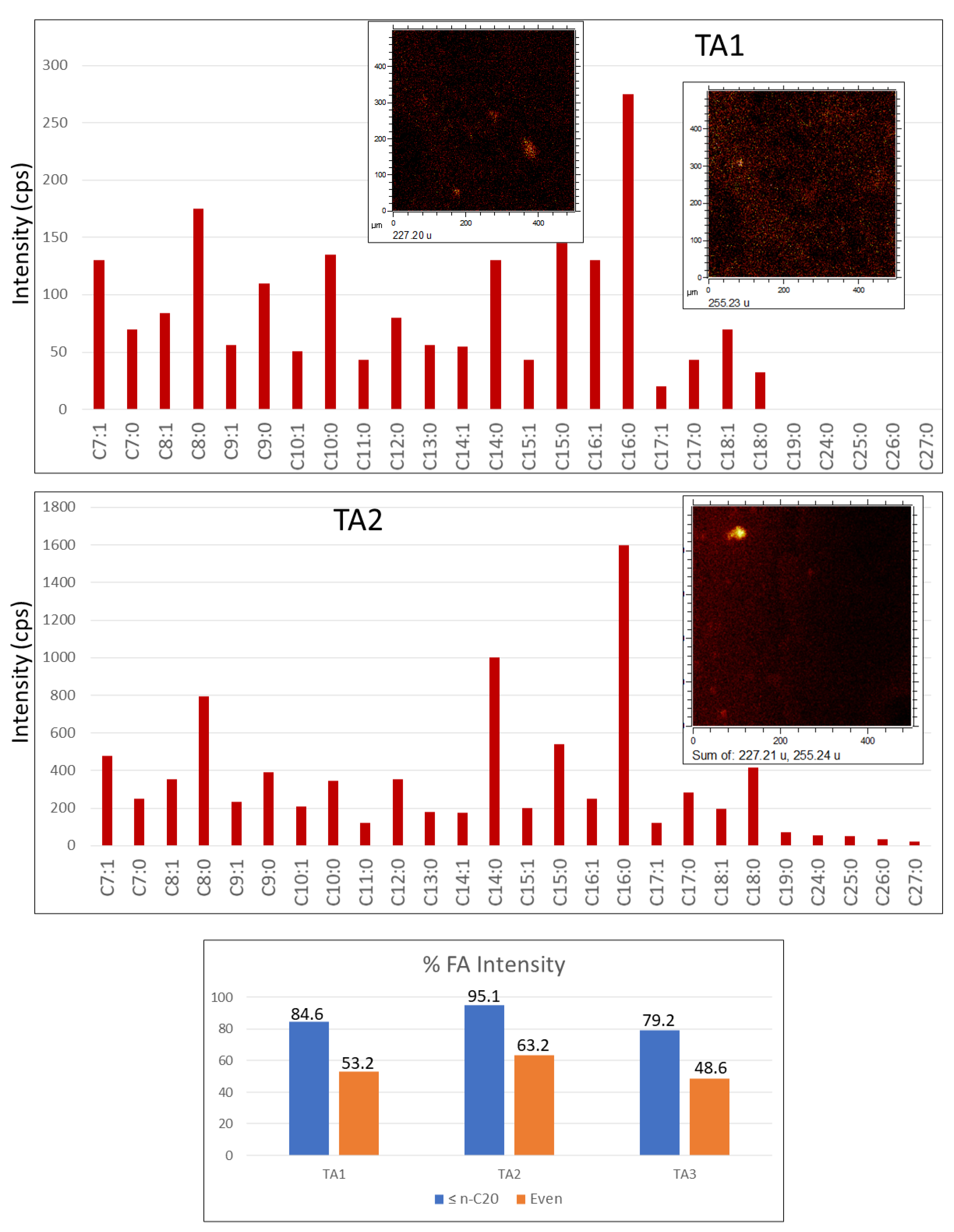
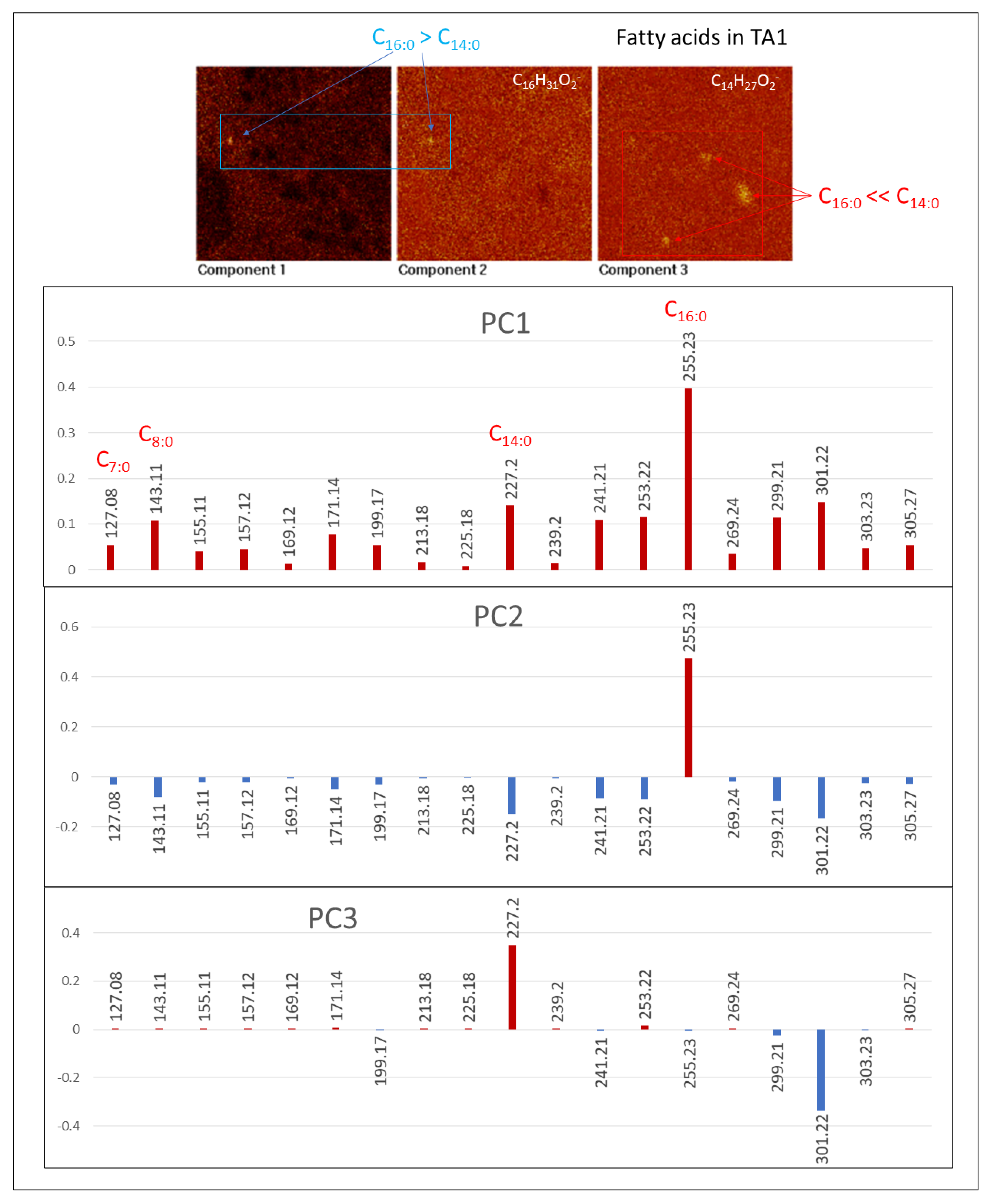
4.2.3. Fatty Acids (FAs)
4.2.4. Acylglycerols
4.2.5. Sphingolipids and Phospholipids
4.2.6. Peptide Fragments and Amino Acids
4.2.7. Hopanoids and Steroids
4.2.8. Other Compounds
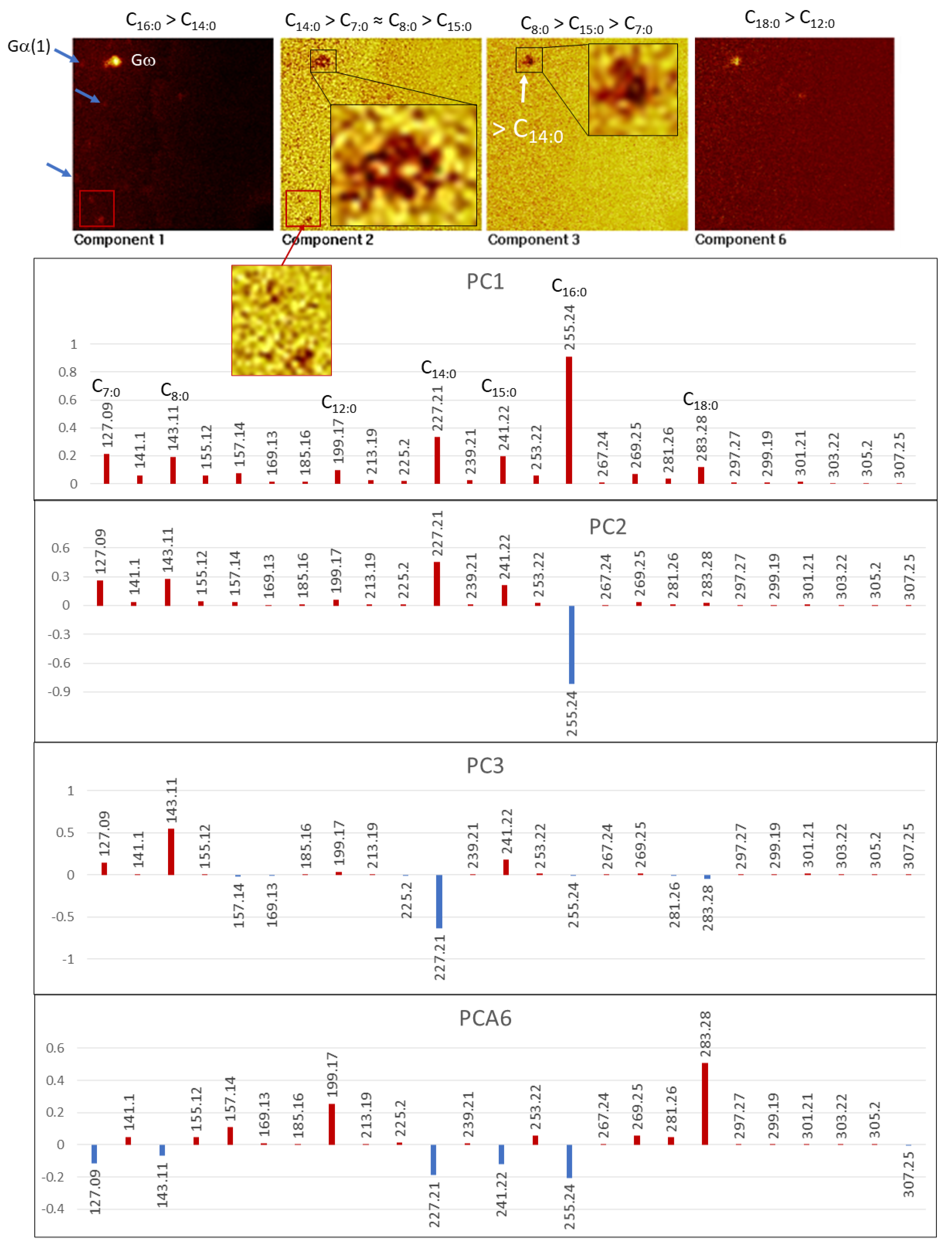
4.3. Principal Component Analysis (PCA) of ToF-SIMS Images
4.3.1. Major Anions
4.3.2. Amino Acid Fragments
4.3.3. [M − H]− FA Adducts
5. Discussion
5.1. Mineralization
5.2. (Bio)mineralization and Microbial Metabolism
5.3. Microstructure and Molecular Associations (MMAs)
5.4. Ancient Microbial Composition Recorded in the Rio Tinto Basement
5.5. Preservation of Biomolecules and Organic Compounds
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gómez-Ortiz, D.; Fernández-Remolar, D.C.; Granda, Á.; Quesada, C.; Granda, T.; Prieto-Ballesteros, O.; Molina, A.; Amils, R. Identification of the subsurface sulfide bodies responsible for acidity in Río Tinto source water, Spain. Earth Planet. Sci. Lett. 2014, 391, 36–41. [Google Scholar] [CrossRef]
- Essalhi, M.; Sizaret, S.; Barbanson, L.; Chen, Y.; Lagroix, F.; Demory, F.; Nieto, J.; Sáez, R.; Capitán, M.Á. A case study of the internal structures of gossans and weathering processes in the Iberian Pyrite Belt using magnetic fabrics and paleomagnetic dating. Miner. Depos. 2011, 46, 981–999. [Google Scholar] [CrossRef]
- Velasco, F.; Herrero, J.M.; Suárez, S.; Yusta, I.; Alvaro, A.; Tornos, F. Supergene features and evolution of gossans capping massive sulphide deposits in the Iberian Pyrite Belt. Ore Geol. Rev. 2013, 53, 181–203. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Morris, R.V.; Gruener, J.E.; Amils, R.; Knoll, A.H. The Rio Tinto Basin, Spain: Mineralogy, sedimentary geobiology, and implications for interpretation of outcrop rocks at Meridiani Planum, Mars. Earth Planet. Sci. Lett. 2005, 240, 149–167. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Gómez, F.; Prieto-Ballesteros, O.; Schelble, R.T.; Rodríguez, N.; Amils, R. Some ecological mechanisms to generate habitability in planetary subsurface areas by chemolithotrophic communities: The Río Tinto subsurface ecosystem as a model system. Astrobiology 2008, 8, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Escudero, C.; del Campo, A.; Ares, J.R.; Sánchez, C.; Martínez, J.M.; Gómez, F.; Amils, R. Visualizing microorganism-mineral interaction in the Iberian Pyrite Belt subsurface: The Acidovorax Case. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Amils, R.; Escudero, C.; Oggerin, M.; Puente-Sánchez, F.; Arce-Rodríguez, A.; Fernandez-Remolar, D.C.; Rodríguez, N.; Garcia-Villadangos, M.; Sanz, J.L.; Briones, C.; et al. Complete C, H, N, S and Fe cycles operating in Iberian Pyrite Belt deep rock matrix microbiome: Pivotal role of N cycle. Proc. Natl. Acad. Sci. USA. under review.
- Escudero, C.; Vera, M.; Oggerin, M.; Amils, R. Active microbial biofilms in deep poor porous continental subsurface rocks. Sci. Rep. 2018, 8, 1538. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; Puente-Sánchez, F.; Parro, V.; Amils, R. Nucleation of Fe-rich phosphates and carbonates on microbial cells and exopolymeric substances. Front. Microbiol. 2015, 6, 1024. [Google Scholar] [CrossRef] [PubMed]
- Puente-Sánchez, F.; Moreno-Paz, M.; Rivas, L.A.; Cruz-Gil, P.; García-Villadangos, M.; Gómez, M.J.; Postigo, M.; Garrido, P.; González-Toril, E.; Briones, C.; et al. Deep subsurface sulfate reduction and methanogenesis in the Iberian Pyrite Belt revealed through geochemistry and molecular biomarkers. Geobiology 2014, 12, 34–47. [Google Scholar] [CrossRef]
- Puente-Sánchez, F.; Sánchez-Román, M.; Amils, R.; Parro, V. Tessaracoccus lapidicaptus sp. nov., an actinobacterium isolated from the deep subsurface of the Iberian pyrite belt. Int. J. Syst. Evol. Microbiol. 2014, 64, 3546–3552. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Remolar, D.C.; Banerjee, N.; Gómez-Ortiz, D.; Izawa, M.; Amils, R. A mineralogical archive of the biogeochemical sulfur cycle preserved in the subsurface of the Río Tinto system. Am. Mineral. 2018, 103, 394–411. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Prieto-Ballesteros, O.; Rodríguez, N.; Gómez, F.; Amils, R.; Gómez-Elvira, J.; Stoker, C.R. Underground habitats in the Río Tinto basin: A model for subsurface life habitats on Mars. Astrobiology 2008, 8, 1023–1047. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Remolar, D.C.; Carrizo, D.; Harir, M.; Huang, T.; Sánchez-García, L.; Gómez-Ortiz, D.; Schmitt-Kopplin, P.; Malmberg, P. Unveiling microbial preservation under hyperacidic and oxidizing conditions in the Oligocene Rio Tinto deposits. Sci. Rep. under review.
- Fernandez-Remolar, D.C.; Gómez-Ortiz, D.; Huang, T.; Angles, A.; Shen, Y.; Hu, Q.; Amils, R.; Rodriguez, N.; Escudero, C.; Banerjee, N. The molecular record of metabolic activity in the subsurface of the Rio Tinto Mars analog. Astrobiology. under review.
- Fernández-Remolar, D.C.; Prieto-Ballesteros, O.; Gómez-Ortíz, D.; Fernández-Sampedro, M.; Sarrazin, P.; Gailhanou, M.; Amils, R. Río Tinto sedimentary mineral assemblages: A terrestrial perspective that suggests some formation pathways of phyllosilicates on Mars. Icarus 2011, 211, 114–138. [Google Scholar] [CrossRef]
- Leistel, J.M.; Marcoux, E.; Thiéblemont, D.; Quesada, C.; Sánchez, A.; Almodóvar, G.R.; Pascual, E.; Sáez, R. The volcanic-hosted massive sulphide deposits of the Iberian Pyrite Belt. Miner. Depos. 1998, 33, 2–30. [Google Scholar] [CrossRef]
- García-Navarro, E.; Fernández, C. Final stages of the Variscan orogeny at the southern Iberian massif: Lateral extrusion and rotation of continental blocks. Tectonics 2004, 23. [Google Scholar] [CrossRef]
- Jabaloy Sánchez, A.; Padrón-Navarta, J.A.; Gómez-Pugnaire, M.T.; López Sánchez-Vizcaíno, V.; Garrido, C.J. Alpine orogeny: Deformation and structure in the Southern Iberian Margin (Betics s.l.). In The Geology of Iberia: A Geodynamic Approach: Volume 3: The Alpine Cycle; Quesada, C., Oliveira, J.T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 453–486. [Google Scholar]
- Manzi, V.; Gennari, R.; Hilgen, F.; Krijgsman, W.; Lugli, S.; Roveri, M.; Sierro, F.J. Age refinement of the Messinian salinity crisis onset in the Mediterranean. Terra Nova 2013, 25, 315–322. [Google Scholar] [CrossRef]
- Sand, W.; Gehrke, T.; Jozsa, P.-G.; Schippers, A. (Bio)chemistry of bacterial leaching—Direct vs. indirect bioleaching. Hydrometallurgy 2001, 59, 159–175. [Google Scholar] [CrossRef]
- Amils, R.; Fernández-Remolar, D.C. Río Tinto: A geochemical and mineralogical terrestrial analogue of Mars. Life 2014, 4, 511–534. [Google Scholar] [CrossRef] [PubMed]
- Stoker, C.R.; Cannon, H.N.; Dunagan, S.E.; Lemke, L.G.; Glass, B.J.; Miller, D.; Gomez-Elvira, J.; Davis, K.; Zavaleta, J.; Winterholler, A.; et al. The 2005 MARTE robotic drilling experiment in Río Tinto, Spain: Objectives, approach, and results of a simulated mission to search for life in the Martian subsurface. Astrobiology 2008, 8, 921–945. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.; Harir, M.; Carrizo, D.; Schmitt-Kopplin, P.; Amils, R. Formation of shale-hosted massive sulfide deposits in the Iberian Pyrite Belt (Tharsis, Spain). J. Geophys. Res. 2018, 123, 1017–1040. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Santamaría, J.; Amils, R.; Parro, V.; Gómez-Ortíz, D.; Izawa, M.R.M.; Banerjee, N.R.; Pérez Rodríguez, R.; Rodríguez, N.; López-Martínez, N. Formation of iron-rich shelled structures by microbial communities. J. Geophys. Res. Biogeosciences 2015, 120, 147–168. [Google Scholar] [CrossRef]
- Patiny, L.; Borel, A. ChemCalc: A building block for tomorrow’s chemical infrastructure. J. Chem. Inf. Model. 2013, 53, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Strohalm, M.; Kavan, D.; Novák, P.; Volný, M.; Havlíček, V. mMass 3: A cross-platform software environment for precise analysis of mass spectrometric data. Anal. Chem. 2010, 82, 4648–4651. [Google Scholar] [CrossRef]
- Pence, H.E.; Williams, A. ChemSpider: An online chemical information resource. J. Chem. Educ. 2010, 87, 1123–1124. [Google Scholar] [CrossRef]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Ther. Drug Monit. 2005, 27, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS online tools for lipid research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef] [PubMed]
- Urbini, M.; Petito, V.; de Notaristefani, F.; Scaldaferri, F.; Gasbarrini, A.; Tortora, L. ToF-SIMS and principal component analysis of lipids and amino acids from inflamed and dysplastic human colonic mucosa. Anal. Bioanal. Chem. 2017, 409, 6097–6111. [Google Scholar] [CrossRef]
- Killops, S.D.; Killops, V. Introduction to Organic Chemistry, 2nd ed.; Blackwell Science: Oxford, UK, 2005; p. 393. [Google Scholar]
- Kögel-Knabner, I.; Amelung, W. 12.7—Dynamics, chemistry, and preservation of organic matter in soils. In Treatise on Geochemistry, 2nd ed.; Holland, H.D., Turekian, K.K., Eds.; Elsevier: Oxford, UK, 2014; pp. 157–215. [Google Scholar]
- Meyers, P.A. Organic geochemical proxies of paleoceanographic, paleolimnologic, and paleoclimatic processes. Org. Geochem. 1997, 27, 213–250. [Google Scholar] [CrossRef]
- Passarelli, M.K.; Winograd, N. Lipid imaging with time-of-flight secondary ion mass spectrometry (ToF-SIMS). Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2011, 1811, 976–990. [Google Scholar] [CrossRef]
- Singh, A.; Del Poeta, M. Sphingolipidomics: An important mechanistic tool for studying fungal pathogens. Front. Microbiol. 2016, 7, 501. [Google Scholar] [CrossRef] [PubMed]
- Phan, N.T.N.; Munem, M.; Ewing, A.G.; Fletcher, J.S. MS/MS analysis and imaging of lipids across Drosophila brain using secondary ion mass spectrometry. Anal. Bioanal. Chem. 2017, 409, 3923–3932. [Google Scholar] [CrossRef]
- Heim, C.; Sjövall, P.; Lausmaa, J.; Leefmann, T.; Thiel, V. Spectral characterisation of eight glycerolipids and their detection in natural samples using time-of-flight secondary ion mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 2741–2753. [Google Scholar] [CrossRef] [PubMed]
- Tyurin, V.A.; Tyurina, Y.Y.; Feng, W.; Mnuskin, A.; Jiang, J.; Tang, M.; Zhang, X.; Zhao, Q.; Kochanek, P.M.; Clark, R.S.B.; et al. Mass-spectrometric characterization of phospholipids and their primary peroxidation products in rat cortical neurons during staurosporine-induced apoptosis. J. Neurochem. 2008, 107, 1614–1633. [Google Scholar] [CrossRef] [PubMed]
- Siljeström, S.; Parenteau, M.N.; Jahnke, L.L.; Cady, S.L. A comparative ToF-SIMS and GC–MS analysis of phototrophic communities collected from an alkaline silica-depositing hot spring. Org. Geochem. 2017, 109, 14–30. [Google Scholar] [CrossRef]
- Kawecki, M.; Bernard, L. Database of proteinogenic amino acid reference spectra for Bismuth-cluster ToF-SIMS. I. Negative polarity. Surf. Sci. Spectra 2018, 25, 015001. [Google Scholar] [CrossRef]
- Kawecki, M.; Bernard, L. Database of proteinogenic amino acid reference spectra for Bismuth-cluster ToF-SIMS. II. Positive polarity. Surf. Sci. Spectra 2018, 25, 015002. [Google Scholar] [CrossRef]
- Srivastava, A.; Pitesky, M.E.; Steele, P.T.; Tobias, H.J.; Fergenson, D.P.; Horn, J.M.; Russell, S.C.; Czerwieniec, G.A.; Lebrilla, C.B.; Gard, E.E.; et al. Comprehensive assignment of mass spectral signatures from individual Bacillus atrophaeus spores in matrix-free laser desorption/ionization bioaerosol mass spectrometry. Anal. Chem. 2005, 77, 3315–3323. [Google Scholar] [CrossRef]
- Leefmann, T.; Heim, C.; Siljeström, S.; Blumenberg, M.; Sjövall, P.; Thiel, V. Spectral characterization of ten cyclic lipids using time-of-flight secondary ion mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Geier, B.; Sogin, E.M.; Michellod, D.; Janda, M.; Kompauer, M.; Spengler, B.; Dubilier, N.; Liebeke, M. Spatial metabolomics of in situ host–microbe interactions at the micrometre scale. Nat. Microbiol. 2020, 5, 498–510. [Google Scholar] [CrossRef] [PubMed]
- Kool, D.M.; Talbot, H.M.; Rush, D.; Ettwig, K.; Sinninghe Damsté, J.S. Rare bacteriohopanepolyols as markers for an autotrophic, intra-aerobic methanotroph. Geochim. Cosmochim. Acta 2014, 136, 114–125. [Google Scholar] [CrossRef]
- Hiraishi, A.; Ueda, Y.; Ishihara, J. Quinone profiling of bacterial communities in natural and synthetic sewage activated sludge for enhanced phosphate removal. Appl. Environ. Microbiol. 1998, 64, 992. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Remolar, D.C.; Sánchez-Román, M.; Hill, A.C.; Gómez-Ortíz, D.; Ballesteros, O.P.; Romanek, C.S.; Amils, R. The environment of early Mars and the missing carbonates. Meteorit. Planet. Sci. 2011, 46, 1447–1469. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Knoll, A.H. Fossilization potential of iron-bearing minerals in acidic environments of Rio Tinto, Spain: Implications for Mars exploration. Icarus 2008, 194, 72–85. [Google Scholar] [CrossRef]
- Li, X.; Hou, L.; Liu, M.; Zheng, Y.; Yin, G.; Lin, X.; Cheng, L.; Li, Y.; Hu, X. Evidence of nitrogen loss from anaerobic ammonium oxidation coupled with ferric iron reduction in an intertidal wetland. Environ. Sci. Technol. 2015, 49, 11560–11568. [Google Scholar] [CrossRef]
- In ‘t Zandt, M.H.; de Jong, A.E.E.; Slomp, C.P.; Jetten, M.S.M. The hunt for the most-wanted chemolithoautotrophic spookmicrobes. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef]
- Fdz-Polanco, F.; Fdz-Polanco, M.; Fernandez, N.; Urueña, M.A.; Garcia, P.A.; Villaverde, S. New process for simultaneous removal of nitrogen and sulphur under anaerobic conditions. Water Res. 2001, 35, 1111–1114. [Google Scholar] [CrossRef]
- Staples, L.W. Nitrate minerals. In Mineralogy; Encyclopedia of Earth Science; Springer US: Boston, MA, USA, 1983; p. 324. [Google Scholar]
- Fennessey, C.M.; Jones, M.E.; Taillefert, M.; DiChristina, T.J. Siderophores are not involved in Fe(III) solubilization during anaerobic Fe(III) Respiration by Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2010, 76, 2425. [Google Scholar] [CrossRef] [PubMed]
- Dehner, C.A.; Awaya, J.D.; Maurice, P.A.; DuBois, J.L. Roles of siderophores, oxalate, and ascorbate in mobilization of iron from hematite by the aerobic bacterium Pseudomonas mendocina. Appl. Environ. Microbiol. 2010, 76, 2041. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; Fernández-Remolar, D.; Amils, R.; Sánchez-Navas, A.; Schmid, T.; Martin-Uriz, P.S.; Rodríguez, N.; McKenzie, J.A.; Vasconcelos, C. Microbial mediated formation of Fe-carbonate minerals under extreme acidic conditions. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Remolar, D.C.; Preston, L.J.; Sánchez-Román, M.; Izawa, M.R.M.; Huang, L.; Southam, G.; Banerjee, N.R.; Osinski, G.R.; Flemming, R.; Gómez-Ortíz, D.; et al. Carbonate precipitation under bulk acidic conditions as a potential biosignature for searching life on Mars. Earth Planet. Sci. Lett. 2012, 351, 13–26. [Google Scholar] [CrossRef]
- Rivadeneyra, M.A.; Martín-Algarra, A.; Sánchez-Román, M.; Sánchez-Navas, A.; Martín-Ramos, J.D. Amorphous Ca-phosphate precursors for Ca-carbonate biominerals mediated by Chromohalobacter marismortui. ISME J. 2010, 4, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Sohlenkamp, C.; Geiger, O. Bacterial membrane lipids: Diversity in structures and pathways. FEMS Microbiol. Rev. 2016, 40, 133–159. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H.; Murphy, R.C.; Raetz, C.R.H.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–862. [Google Scholar] [CrossRef] [PubMed]
- Holloway, J.M.; Dahlgren, R.A. Nitrogen in rock: Occurrences and biogeochemical implications. Glob. Biogeochem. Cycles 2002, 16, 65-1–65-17. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Joergensen, R.G.; Ludwig, B. Pathways of nitrogen utilization by soil microorganisms—A review. Soil Biol. Biochem. 2010, 42, 2058–2067. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.; Rodríguez, N.; Gómez, F.; Amils, R. The geological record of an acidic environment driven by iron hydrochemistry: The Tinto River system. J. Geophys. Res. 2003, 108. [Google Scholar] [CrossRef]
- Preston, L.J.; Shuster, J.; FernÁNdez-Remolar, D.; Banerjee, N.R.; Osinski, G.R.; Southam, G. The preservation and degradation of filamentous bacteria and biomolecules within iron oxide deposits at Rio Tinto, Spain. Geobiology 2011, 9, 233–249. [Google Scholar] [CrossRef]
- Konhauser, K.; Riding, R. Bacterial biomineralization. In Fundamentals of Geobiology; Blackwell Publishing: Oxford, UK, 2012; pp. 105–130. [Google Scholar]
- Amils, R.; Fernandez Remolar, D. Rio Tinto: An extreme acidic environmental model of astrobiological interest. In Extremophiles as Astrobiological Models; Seckbach, J., Stan-Lotter, H., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 21–44. [Google Scholar]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Remolar, D.C.; Gómez-Ortiz, D.; Malmberg, P.; Huang, T.; Shen, Y.; Anglés, A.; Amils, R. Preservation of Underground Microbial Diversity in Ancient Subsurface Deposits (>6 Ma) of the Rio Tinto Basement. Microorganisms 2021, 9, 1592. https://doi.org/10.3390/microorganisms9081592
Fernández-Remolar DC, Gómez-Ortiz D, Malmberg P, Huang T, Shen Y, Anglés A, Amils R. Preservation of Underground Microbial Diversity in Ancient Subsurface Deposits (>6 Ma) of the Rio Tinto Basement. Microorganisms. 2021; 9(8):1592. https://doi.org/10.3390/microorganisms9081592
Chicago/Turabian StyleFernández-Remolar, David C., David Gómez-Ortiz, Per Malmberg, Ting Huang, Yan Shen, Angélica Anglés, and Ricardo Amils. 2021. "Preservation of Underground Microbial Diversity in Ancient Subsurface Deposits (>6 Ma) of the Rio Tinto Basement" Microorganisms 9, no. 8: 1592. https://doi.org/10.3390/microorganisms9081592