
Yeasts Associated with the Small-Intestinal Contents and Epithelium of Pon Yang Kham (Charolais Crossbred) Fattening Beef Cattle

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Yeast Isolation
2.3. Yeast Identification
2.4. Phylogenetic Analysis
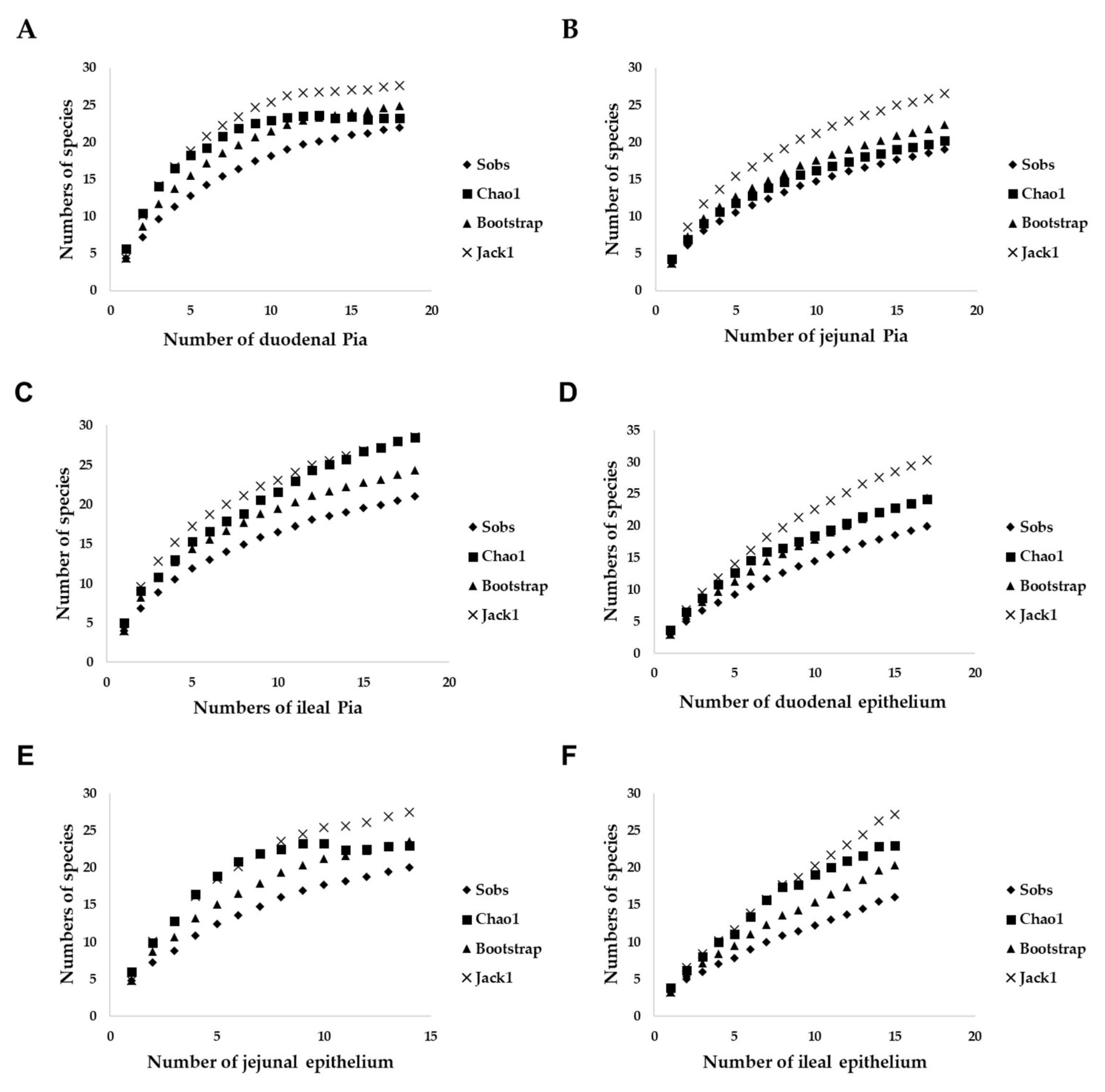
2.5. Biodiversity Analyses
3. Results
3.1. Sample Collection and Yeast Isolation
3.2. Identification of Yeasts from Pia
3.3. Comparison of Yeasts from Pia of Three Parts of the Small Intestine
3.4. Identification of Yeasts from the Small-Intestinal Epithelium
3.5. Comparison of Yeasts Obtained from the Small-Intestinal Epithelium in All Three Parts
3.6. Comparison of the Yeast Communities of the Small-Intestinal Pia and Epithelium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Pongpaew, P.; Klaypradit, W.; Srikalong, P.; Ingkasupart, P.; Kerdpiboon, S. Fatty acid composition of pon yang kham beef tallow. In Proceedings of the International Congresses of Meat Science and Technology (ICoMST), Melbourne, Australia, 21 August 2018. [Google Scholar]
- Wiwatthanapornchai, S. Analysis of suitable farm size for fattening Pon Yang Kham cattle. Mod. Appl. Sci. 2019, 13, 7. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. WJG 2015, 21, 8787. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fecteau, M.-E.; Pitta, D.W.; Vecchiarelli, B.; Indugu, N.; Kumar, S.; Gallagher, S.C.; Fyock, T.L.; Sweeney, R.W. Dysbiosis of the fecal microbiota in cattle infected with Mycobacterium avium subsp. paratuberculosis. PLoS ONE 2016, 11, e0160353. [Google Scholar] [CrossRef] [PubMed]
- Koboziev, I.; Webb, C.R.; Furr, K.L.; Grisham, M.B. Role of the enteric microbiota in intestinal homeostasis and inflammation. Free Radic. Biol. Med. 2014, 68, 122–133. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.M.; Vieira, A.T.; Vinolo, M.A.R.; Oliveira, F.A.; Curi, R.; Martins, F.d.S. The central role of the gut microbiota in chronic inflammatory diseases. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, F.; Tremaroli, V.; Nielsen, J.; Bäckhed, F. Assessing the human gut microbiota in metabolic diseases. Diabetes 2013, 62, 3341–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisgaard, H.; Li, N.; Bonnelykke, K.; Chawes, B.L.K.; Skov, T.; Paludan-Müller, G.; Stokholm, J.; Smith, B.; Krogfelt, K.A. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J. Allergy Clin. Immunol. 2011, 128, 646–652. [Google Scholar] [CrossRef]
- Natividad, J.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How colonization by microbiota in early life shapes the immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Hooper, L.V.; Midtvedt, T.; Gordon, J.I. How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr. 2002, 22, 283–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firkins, J.; Yu, Z. Ruminant nutrition symposium: How to use data on the rumen microbiome to improve our understanding of ruminant nutrition. J. Anim. Sci. 2015, 93, 1450–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäumler, A.J.; Sperandio, V. Interactions between the microbiota and pathogenic bacteria in the gut. Nature 2016, 535, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, C.; Peter, G. Biodiversity and Ecophysiology of Yeasts; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Pinheiro, R.E.E.; Rodrigues, A.M.D.; Santos, J.T.d.O.; Costa, J.d.A.; Pereyra, C.M.; Torres, A.M.; Rosa, C.A.; Santos, A.R.d.O.; Muratori, M.C.S. Occurrence and diversity of yeast species isolated from fish feed and tambatinga gut. Lat. Am. J. Aquat. Res. 2018, 46, 837–842. [Google Scholar] [CrossRef]
- Sirisan, V.; Pattarajinda, V.; Vichitphan, K.; Leesing, R. Isolation, identification and growth determination of lactic acid-utilizing yeasts from the ruminal fluid of dairy cattle. Lett. Appl. Microbiol. 2013, 57, 102–107. [Google Scholar] [CrossRef]
- Fernandes, T.; Carvalho, B.; Mantovani, H.; Schwan, R.; Ávila, C. Identification and characterization of yeasts from bovine rumen for potential use as probiotics. J. Appl. Microbiol. 2019, 127, 845–855. [Google Scholar] [CrossRef]
- Lee, J.-H.; Kumar, S.; Lee, G.-H.; Chang, D.-H.; Rhee, M.-S.; Yoon, M.-H.; Kim, B.-C. Methanobrevibacterboviskoreani sp. nov., isolated from the rumen of Korean native cattle. Int. J. Syst. Evol. Microbiol. 2013, 63, 4196–4201. [Google Scholar] [CrossRef] [Green Version]
- Van Uden, N.; Carmo-sousa, L.D. Yeasts from the bovine caecum. J. Gen. Microbiol. 1957, 16, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Van Uden, N.; Sousa, L.D.C.; Farinha, M. On the intestinal yeast flora of horses, sheep, goats and swine. Microbiology 1958, 19, 435–445. [Google Scholar] [CrossRef] [Green Version]
- Sokół, I.; Gaweł, A.; Bobrek, K. The prevalence of yeast and characteristics of the isolates from the digestive tract of clinically healthy turkeys. Avian Dis. 2018, 62, 286–290. [Google Scholar] [CrossRef]
- Seyedmousavi, S.; Bosco, S.d.M.; De Hoog, S.; Ebel, F.; Elad, D.; Gomes, R.R.; Jacobsen, I.D.; Jensen, H.E.; Martel, A.; Mignon, B. Fungal infections in animals: A patchwork of different situations. Med. Mycol. 2018, 56, S165–S187. [Google Scholar] [CrossRef]
- Vohra, A.; Syal, P.; Madan, A. Probiotic yeasts in livestock sector. Anim. Feed Sci. Technol. 2016, 219, 31–47. [Google Scholar] [CrossRef]
- Zheng, X.-W.; Yan, Z.; Han, B.-Z.; Zwietering, M.H.; Samson, R.A.; Boekhout, T.; Nout, M.R. Complex microbiota of a Chinese “Fen” liquor fermentation starter (Fen-Daqu), revealed by culture-dependent and culture-independent methods. Food Microbiol. 2012, 31, 293–300. [Google Scholar] [CrossRef]
- Davies, D.R.; Theodorou, M.K.; Lawrence, M.I.; Trinci, A.P. Distribution of anaerobic fungi in the digestive tract of cattle and their survival in faeces. Microbiology 1993, 139, 1395–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Barba, J.L.; Maldonado-Barragán, A.; Jiménez Díaz, R. Small-scale total DNA extraction from bacteria and yeast for PCR applications. Anal. Biochem. 2005, 347, 333–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Van Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J. Phylogenetic relationships among yeasts of the ‘Saccharomyces complex’determined from multigene sequence analyses. FEMS Yeast Res. 2003, 3, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Fell, J.W.; Boekhout, T.; Fonseca, A.; Scorzetti, G.; Statzell-Tallman, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1351–1371. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Colwell, R.K.; Chao, A.; Gotelli, N.J.; Lin, S.-Y.; Mao, C.X.; Chazdon, R.L.; Longino, J.T. Models and estimators linking individual-based and sample-based rarefaction, extrapolation and comparison of assemblages. J. Plant Ecol. 2012, 5, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Brake, D.; Titgemeyer, E.; Bailey, E.; Anderson, D. Small intestinal digestion of raw cornstarch in cattle consuming a soybean hull-based diet is improved by duodenal casein infusion. J. Anim. Sci. 2014, 92, 4047–4056. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.; Young, A.; Jorgensen, N. Effect of dietary corn starch intake on pancreatic amylase and intestinal maltase and pH in cattle. J. Anim. Sci. 1981, 52, 1177–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macrae, J.C. Carbohydrate Digestion in the Intestinal Tract of the Mature Sheep. Ph.D. Thesis, Newcastle University, Newcastle upon Tyne, UK, 1967. [Google Scholar]
- Lennox, A.M.; Garton, G. The absorption of long-chain fatty acids from the small intestine of the sheep. Br. J. Nutr. 1968, 22, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Pierzynowski, S.; Barej, W.; Mikołajczyk, M.; Zabielski, R. The influence of light fermented carbohydrates on the exocrine pancreatic secretion in cows. J. Anim. Physiol. Anim. Nutr. 1988, 60, 234–238. [Google Scholar] [CrossRef]
- Pitt, J.I. Resistance of Some Food Spoilage Yeasts to Preservatives. Food Technol. Austral. 1974, 6, 238–241. [Google Scholar]
- Aono, R. Taxonomic distribution of alkali-tolerant yeasts. Syst. Appl. Microbiol. 1990, 13, 394–397. [Google Scholar] [CrossRef]
- Kurita, O.; Yamazaki, E. Growth under alkaline conditions of the salt-tolerant yeast Debaryomyces hansenii IFO10939. Curr. Microbiol. 2002, 45, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Guinan, J.; Villa, P.; Thangamani, S. Secondary bile acids inhibit Candida albicans growth and morphogenesis. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cafarchia, C.; Camarda, A.; Romito, D.; Campolo, M.; Quaglia, N.; Tullio, D.; Otranto, D. Occurrence of yeasts in cloacae of migratory birds. Mycopathologia 2006, 161, 229–234. [Google Scholar] [CrossRef]
- Lund, A. Yeasts and moulds in the bovine rumen. Microbiology 1974, 81, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves Dos Santos, M.T.P.; Benito, M.J.; Córdoba, M.d.G.; Alvarenga, N.; Ruiz-Moyano Seco de Herrera, S. Yeast community in traditional Portuguese Serpa cheese by culture-dependent and -independent DNA approaches. Int. J. Food Microbiol. 2017, 262, 63–70. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Purahong, W.; Wubet, T.; Hyde, K.D.; Zhang, W.; Xu, H.; Zhang, G.; Fu, C.; Liu, M.; Xing, Q. Direct comparison of culture-dependent and culture-independent molecular approaches reveal the diversity of fungal endophytic communities in stems of grapevine (Vitis vinifera). Fungal Divers. 2018, 90, 85–107. [Google Scholar] [CrossRef]
- Hamad, I.; Ranque, S.; Azhar, E.I.; Yasir, M.; Jiman-Fatani, A.A.; Tissot-Dupont, H.; Raoult, D.; Bittar, F. Culturomics and amplicon-based metagenomic approaches for the study of fungal population in human gut microbiota. Sci. Rep. 2017, 7, 1–8. [Google Scholar]
- Al-Yasiri, M.H.; Normand, A.-C.; Piarroux, R.; Ranque, S.; Mauffrey, J.-F. Gut yeast communities in Larus michahellis from various breeding colonies. Med Mycol. 2017, 55, 436–444. [Google Scholar]
- Pacynska, J. Yeast-like fungi and yeasts isolated from healthy breeding horses. Pol. J. Vet. Sci. 2013, 16, 69. [Google Scholar]
- Różański, P.; Ślaska, B.; Różańska, D. Prevalence of yeasts in English full blood mares. Mycopathologia 2013, 175, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, E.H.; Fontenelle, R.O.; Brilhante, R.S.; Cordeiro, R.A.; Monteiro, A.J.; Sidrim, J.J.; Rocha, M.F. The anatomical distribution and antimicrobial susceptibility of yeast species isolated from healthy dogs. Vet. J. 2009, 182, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Kam, A.P.; Xu, J. Diversity of commensal yeasts within and among healthy hosts. Diagn. Microbiol. Infect. Dis. 2002, 43, 19–28. [Google Scholar] [CrossRef]
- Bassetti, M.; Merelli, M.; Righi, E.; Diaz-Martin, A.; Rosello, E.M.; Luzzati, R.; Parra, A.; Trecarichi, E.M.; Sanguinetti, M.; Posteraro, B. Epidemiology, species distribution, antifungal susceptibility, and outcome of candidemia across five sites in Italy and Spain. J. Clin. Microbiol. 2013, 51, 4167–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diekema, D.; Arbefeville, S.; Boyken, L.; Kroeger, J.; Pfaller, M. The changing epidemiology of healthcare-associated candidemia over three decades. Diagn. Microbiol. Infect. Dis. 2012, 73, 45–48. [Google Scholar] [CrossRef]
- Pappas, P.G. Invasive candidiasis. Infect. Dis. Clin. 2006, 20, 485–506. [Google Scholar] [CrossRef] [PubMed]
- Krcmery, V.; Barnes, A. Non-albicans Candida spp. causing fungaemia: Pathogenicity and antifungal resistance. J. Hosp. Infect. 2002, 50, 243–260. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of invasive candidiasis: A persistent public health problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagas-Neto, T.C.; Chaves, G.M.; Colombo, A.L. Update on the genus Trichosporon. Mycopathologia 2008, 166, 121. [Google Scholar] [CrossRef]
- Rodrigues, G.S.; De Faria, R.; Guazzelli, L.; Oliveira, F.M.; Severo, L. Nosocomial infection due to Trichosporon asahii: Clinical revision of 22 cases. Rev. Iberoam. De Micol. 2006, 23, 85. [Google Scholar] [CrossRef]
- Girmenia, C.; Pagano, L.; Martino, B.; D’Antonio, D.; Fanci, R.; Specchia, G.; Melillo, L.; Buelli, M.; Pizzarelli, G.; Venditti, M. Invasive infections caused by Trichosporon species and Geotrichum capitatum in patients with hematological malignancies: A retrospective multicenter study from Italy and review of the literature. J. Clin. Microbiol. 2005, 43, 1818–1828. [Google Scholar] [CrossRef] [Green Version]
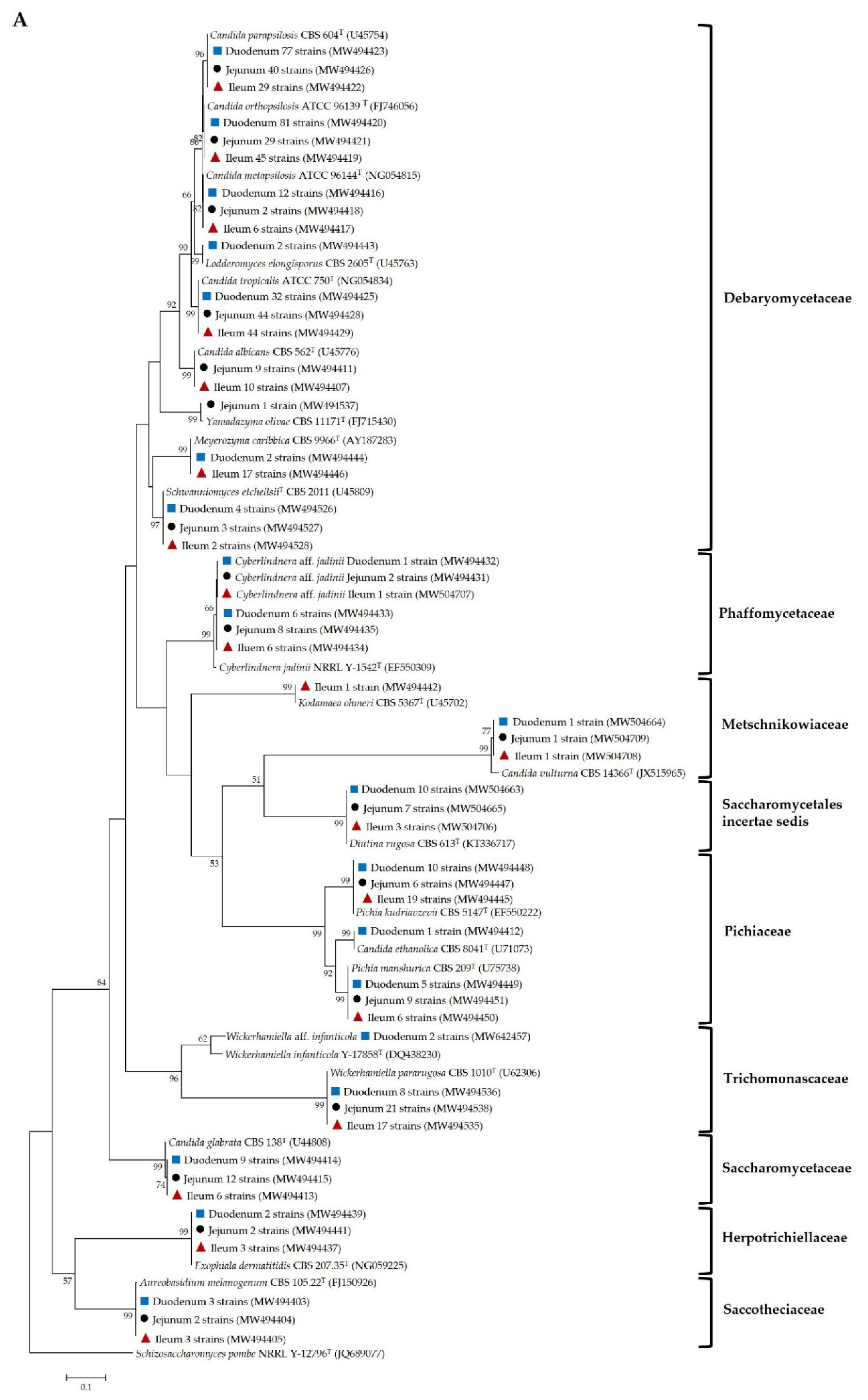
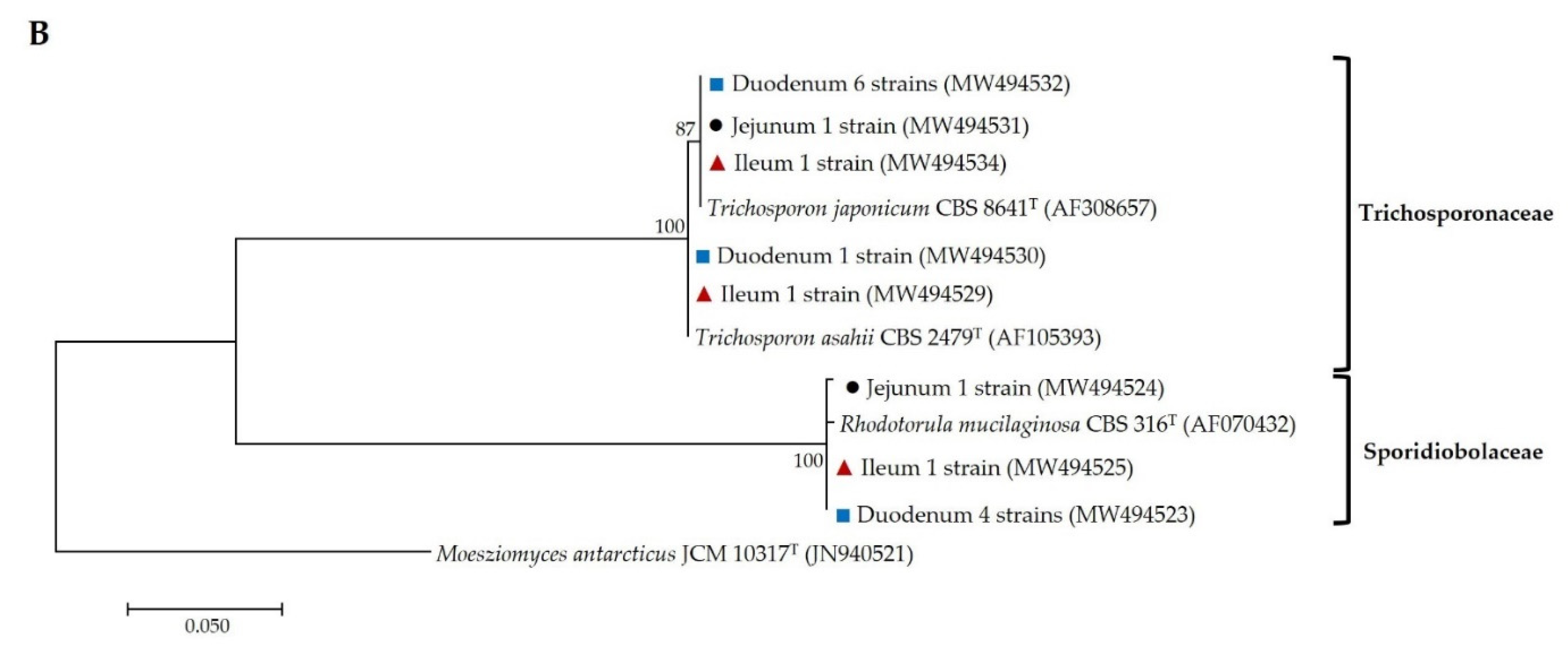
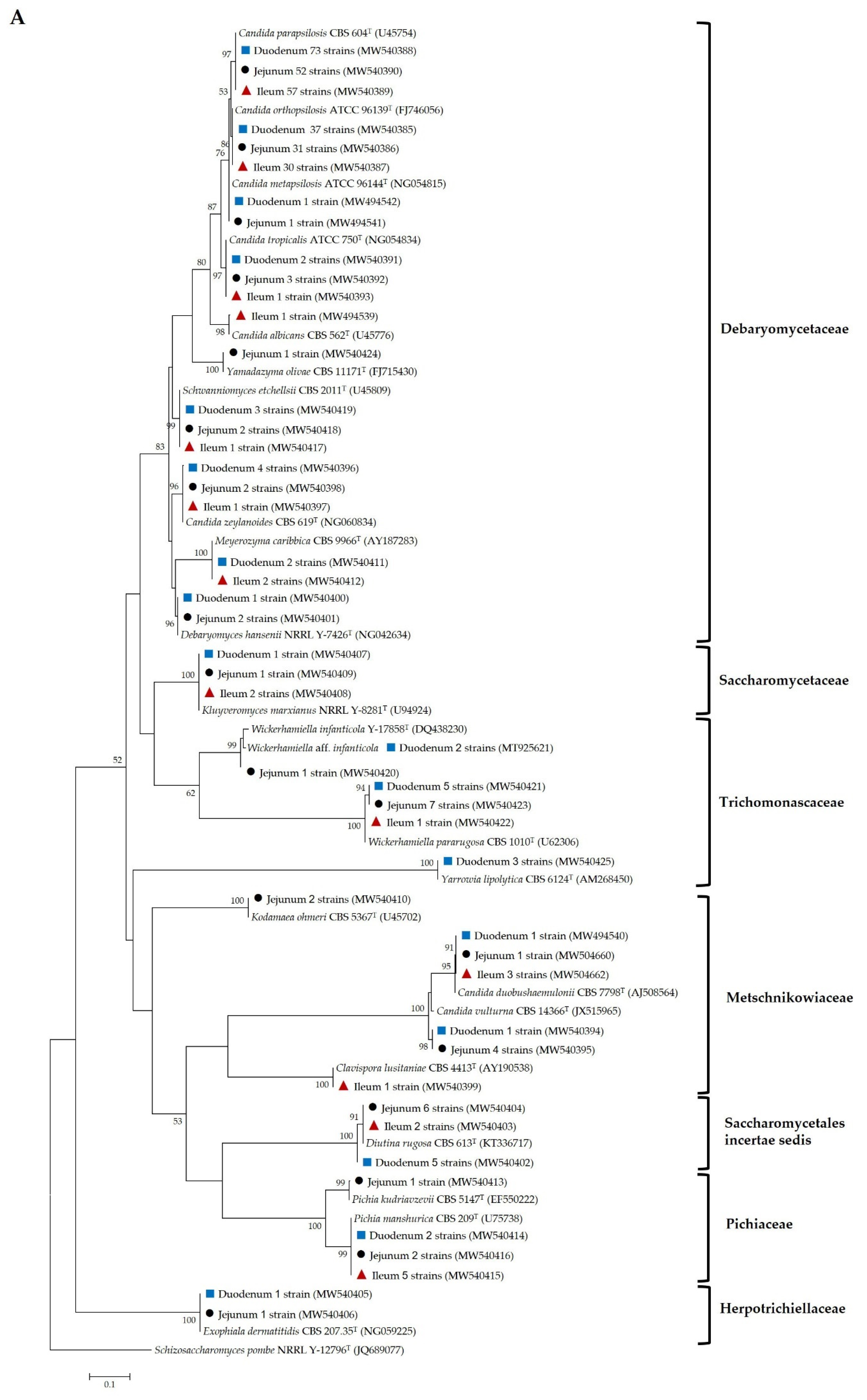
 ), jejunal pia (●) and ileal pia (
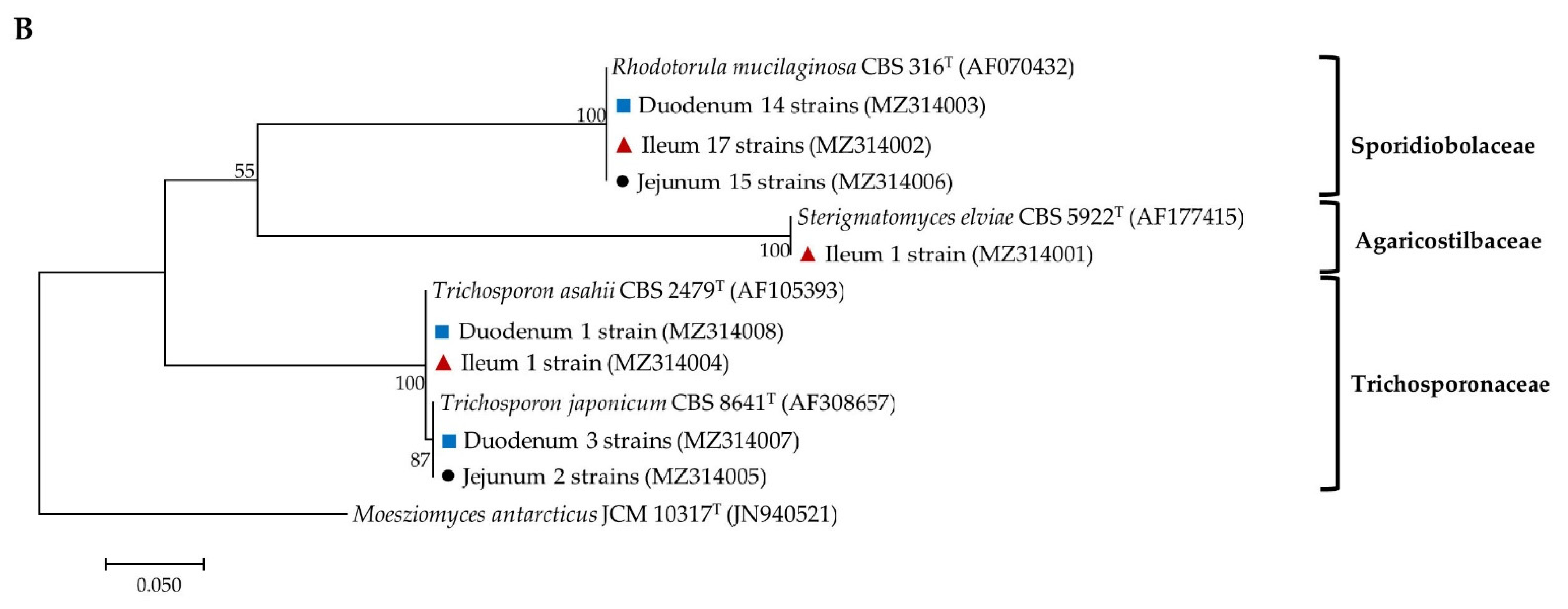
), jejunal pia (●) and ileal pia (  ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) Phylum Basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.
), jejunal pia (●) and ileal pia ( ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) Phylum Basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.
) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) Phylum Basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.
), jejunal pia (●) and ileal pia ( ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) Phylum Basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.


 ), jejuna (●) and ilea ( ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) phylum basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.
), jejuna (●) and ilea ( ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) phylum basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.
), jejuna (●) and ilea ( ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) phylum basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.
), jejuna (●) and ilea ( ) and their closely related type strain sequences received from the GenBank database. Numbers on branches demonstrate the bootstrap percentages (>50%), derived from 1000 replications. (A) Phylum Ascomycota; scale bar, 0.1 substitutions per nucleotide position; Schizosaccharomyces pombe NRRL Y-12796T (JQ689077) was used as the outgroup; (B) phylum basidiomycota; scale bar, 0.05 substitutions per nucleotide position; Moesziomyces antarcticus JCM 10317T (JN940521) was used as the outgroup.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Sample Site | Average pH Value | Number of Samples | No. of Yeast Isolates |
|---|---|---|---|---|
| Pia | Duodenum | 5.9 | 18 | 279 |
| Jejunum | 7.2 | 18 | 200 | |
| Ileum | 7.8 | 18 | 222 | |
| Total | 54 | 701 | ||
| Epithelium | Duodenum | - | 17 | 162 |
| Jejunum | - | 16 | 139 | |
| Ileum | - | 17 | 124 | |
| Total | 50 | 425 |
| Diversity Indices | Duodenum | Jejunum | Ileum | |||
|---|---|---|---|---|---|---|
| Pia | Epithelium | Pia | Epithelium | Pia | Epithelium | |
| Total number of collected samples | 18 | 17 | 18 | 16 | 18 | 17 |
| Total number of samples obtained yeasts | 18 | 17 | 18 | 14 | 18 | 15 |
| Total number of yeast isolates | 279 | 162 | 200 | 139 | 222 | 124 |
| Total number of yeast species (S) | 22 | 20 | 19 | 20 | 21 | 16 |
| Shannon-Weiner index (H’) | 2.2 | 1.8 | 2.3 | 2.0 | 2.4 | 1.7 |
| Equitability index (EH) | 0.7 | 0.6 | 0.8 | 0.7 | 0.8 | 0.6 |
| The most prevalent known yeast species | C. parapsilosis | C. parapsilosis | C. parapsilosis | C. parapsilosis | C. parapsilosis, C. tropicalis | C. parapsilosis |
| Yeast Taxa | Relative Frequency (%) 1/Frequency of Occurrence (%) 2 | ||
|---|---|---|---|
| Duodenum | Jejunum | Ileum | |
| Ascomycota | |||
| Pezizomycotina | |||
| Herpotrichiellaceae | |||
| Exophiala dermatitidis | 0.7/11.1 | 1.0/5.6 | 1.4/11.1 |
| Saccotheciaceae | |||
| Aureobasidium melanogenum | 1.1/5.6 | 1.0/5.6 | 1.4/5.6 |
| Saccharomycotina | |||
| Debaryomycetaceae | |||
| Candida albicans | nd | 4.5/11.1 | 4.5/11.1 |
| Candida metapsilosis | 4.3/38.9 | 1.0/11.1 | 2.7/22.2 |
| Candida orthopsilosis | 29.0/66.7 | 14.5/50.0 | 20.3/55.6 |
| Candida parapsilosis | 27.6/83.3 | 20.0/77.8 | 13.1/61.1 |
| Candida tropicalis | 11.5/38.9 | 22.0/50.0 | 19.8/61.1 |
| Lodderomyces elongisporus | 0.7/11.1 | nd | nd |
| Meyerozyma caribbica | 0.7/11.1 | nd | 7.7/5.6 |
| Schwanniomyces etchellsii | 1.4/16.7 | 1.5/5.6 | 0.9/11.1 |
| Yamadazyma olivae | nd | 0.5/5.6 | nd |
| Metschnikowiaceae | |||
| Candida vulturna | 0.4/5.6 | 0.5/5.6 | 0.5/5.6 |
| Kodamaea ohmeri | nd | nd | 0.5/5.6 |
| Phaffomycetaceae | |||
| Cyberlindnera aff. jadinii | 0.4/5.6 | 1.0/5.6 | 0.5/5.6 |
| Cyberlindnera jadinii | 2.2/11.1 | 4.0/22.2 | 2.7/22.2 |
| Pichiaceae | |||
| Candida ethanolica | 0.4/5.6 | nd | nd |
| Pichia kudriavzevii | 3.6/16.7 | 3.0/11.1 | 8.6/22.2 |
| Pichia manshurica | 1.8/16.7 | 4.5/27.8 | 2.7/27.8 |
| Saccharomycetaceae | |||
| Candida glabrata | 3.2/16.7 | 6.0/16.7 | 2.7/16.7 |
| Saccharomycetales incertae sedis | |||
| Diutina rugosa | 3.6/33.3 | 3.5/16.7 | 1.4/16.7 |
| Trichomonascaceae | |||
| Wickerhamiella aff. infanticola | 0.7/5.6 | nd | nd |
| Wickerhamiella pararugosa | 2.9/27.8 | 10.5/38.9 | 7.7/44.4 |
| Basidiomycota | |||
| Agaricomycotina | |||
| Trichosporonaceae | |||
| Trichosporon asahii | 0.4/5.6 | nd | 0.5/5.6 |
| Trichosporon japonicum | 2.2/16.7 | 0.5/5.6 | 0.5/5.6 |
| Pucciniomycotina | |||
| Sporidiobolaceae | |||
| Rhodotorula mucilaginosa | 1.4/16.7 | 0.5/5.6 | 0.5/5.6 |
| Yeast Taxa | Relative Frequency (%) 1/Frequency of Occurrence (%) 2 | ||
|---|---|---|---|
| Duodenum | Jejunum | Ileum | |
| Ascomycota | |||
| Pezizomycotina | |||
| Herpotrichiellaceae | |||
| Exophiala dermatitidis | 0.6/5.9 | 0.7/6.3 | nd |
| Saccharomycotina | |||
| Debaryomycetaceae | |||
| Candida albicans | Nd | nd | 0.8/5.9 |
| Candida metapsilosis | 0.6/5.9 | 0.7/6.3 | nd |
| Candida orthopsilosis | 22.8/58.8 | 22.3/56.3 | 24.2/58.8 |
| Candida parapsilosis | 45.1/88.2 | 37.4/87.5 | 46.0/88.2 |
| Candida tropicalis | 1.2/11.8 | 2.2/18.8 | 0.8/5.9 |
| Candida zeylanoides | 2.5/5.9 | 1.4/6.3 | 0.8/5.9 |
| Debaryomyces hansenii | 0.6/5.9 | 1.4/12.5 | nd |
| Meyerozyma caribbica | 1.2/5.9 | nd | 1.6/5.9 |
| Schwanniomyces etchellsii | 1.9/17.6 | 1.4/12.5 | 0.8/5.9 |
| Yamadazyma olivae | nd | 0.7/6.3 | nd |
| Metschnikowiaceae | |||
| Candida duobushaemulonii | 0.6/5.9 | 0.7/6.3 | 2.4/5.9 |
| Candida vulturna | 0.6/5.9 | 2.9/18.8 | nd |
| Clavispora lusitaniae | nd | nd | 0.8/5.9 |
| Kodamaea ohmeri | nd | 1.4/12.5 | nd |
| Pichiaceae | |||
| Pichia kudriavzevii | nd | 0.7/6.3 | nd |
| Pichia manshurica | 1.2/5.9 | 1.4/12.5 | 4.0/5.9 |
| Saccharomycetaceae | |||
| Kluyveromyces marxianus | 0.6/5.9 | 0.7/6.3 | 1.6/5.9 |
| Saccharomycetales incertae sedis | |||
| Diutina rugosa | 3.1/11.8 | 4.3/25 | 1.6/11.8 |
| Trichomonascaceae | |||
| Wickerhamiella aff. infanticola | 1.2/11.8 | nd | nd |
| Wickerhamiella infanticola | nd | 0.7/6.3 | nd |
| Wickerhamiella pararugosa | 3.1/23.5 | 5.0/18.8 | 0.8/5.9 |
| Yarrowia lipolytica | 1.9/11.8 | nd | nd |
| Basidiomycota | |||
| Agaricomycotina | |||
| Trichosporonaceae | |||
| Trichosporon asahii | 0.6/5.9 | nd | 0.8/5.9 |
| Trichosporon japonicum | 1.9/5.9 | 1.4/12.5 | nd |
| Pucciniomycotina | |||
| Agaricostilbaceae | |||
| Sterigmatomyces elviae | nd | nd | 0.8/5.9 |
| Sporidiobolaceae | |||
| Rhodotorula mucilaginosa | 8.6/52.9 | 12.2/43.8 | 12.1/52.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angchuan, J.; Khunnamwong, P.; Wongpanit, K.; Limtong, S.; Srisuk, N. Yeasts Associated with the Small-Intestinal Contents and Epithelium of Pon Yang Kham (Charolais Crossbred) Fattening Beef Cattle. Microorganisms 2021, 9, 1444. https://doi.org/10.3390/microorganisms9071444
Angchuan J, Khunnamwong P, Wongpanit K, Limtong S, Srisuk N. Yeasts Associated with the Small-Intestinal Contents and Epithelium of Pon Yang Kham (Charolais Crossbred) Fattening Beef Cattle. Microorganisms. 2021; 9(7):1444. https://doi.org/10.3390/microorganisms9071444
Chicago/Turabian StyleAngchuan, Jirameth, Pannida Khunnamwong, Kannika Wongpanit, Savitree Limtong, and Nantana Srisuk. 2021. "Yeasts Associated with the Small-Intestinal Contents and Epithelium of Pon Yang Kham (Charolais Crossbred) Fattening Beef Cattle" Microorganisms 9, no. 7: 1444. https://doi.org/10.3390/microorganisms9071444