
The RNA Chaperone Hfq Participates in Persistence to Multiple Antibiotics in the Fish Pathogen Yersinia ruckeri

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
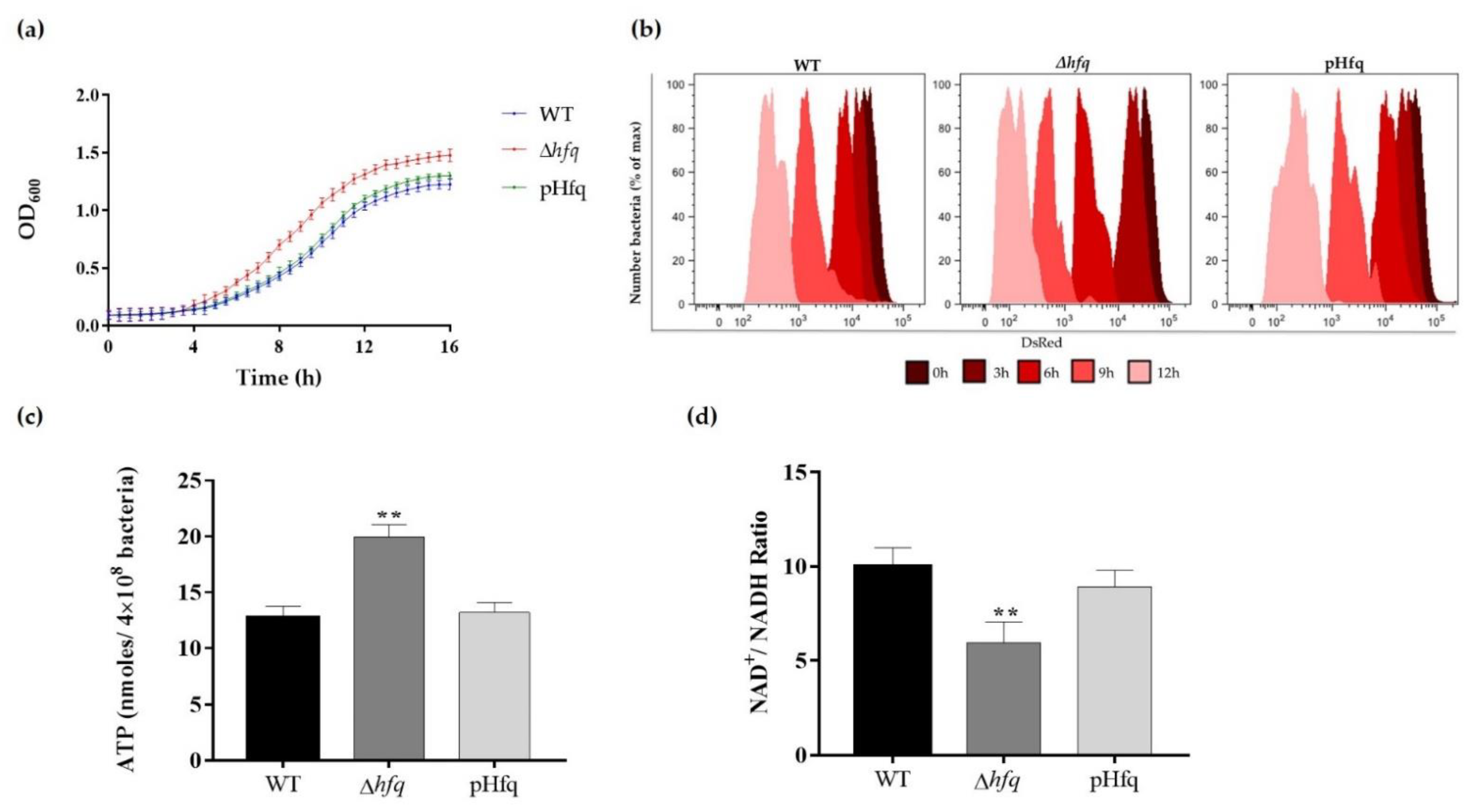
3.1. Hfq Deletion Results in Bacteria with Faster Growth and More Active Metabolism
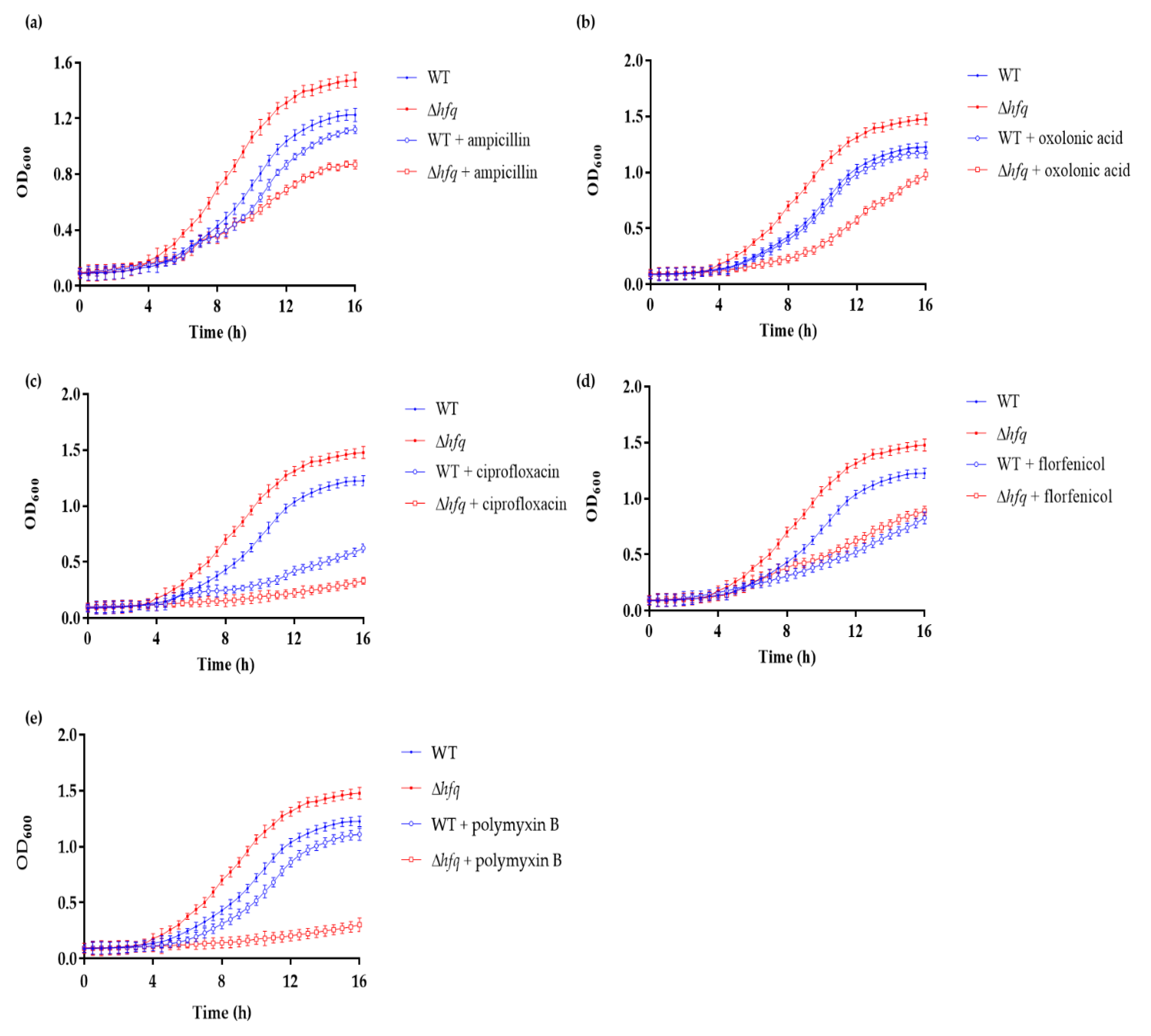
3.2. Y. ruckeri Δhfq Exhibits Increased Susceptibility to Sublethal Concentrations of Different Antibiotics
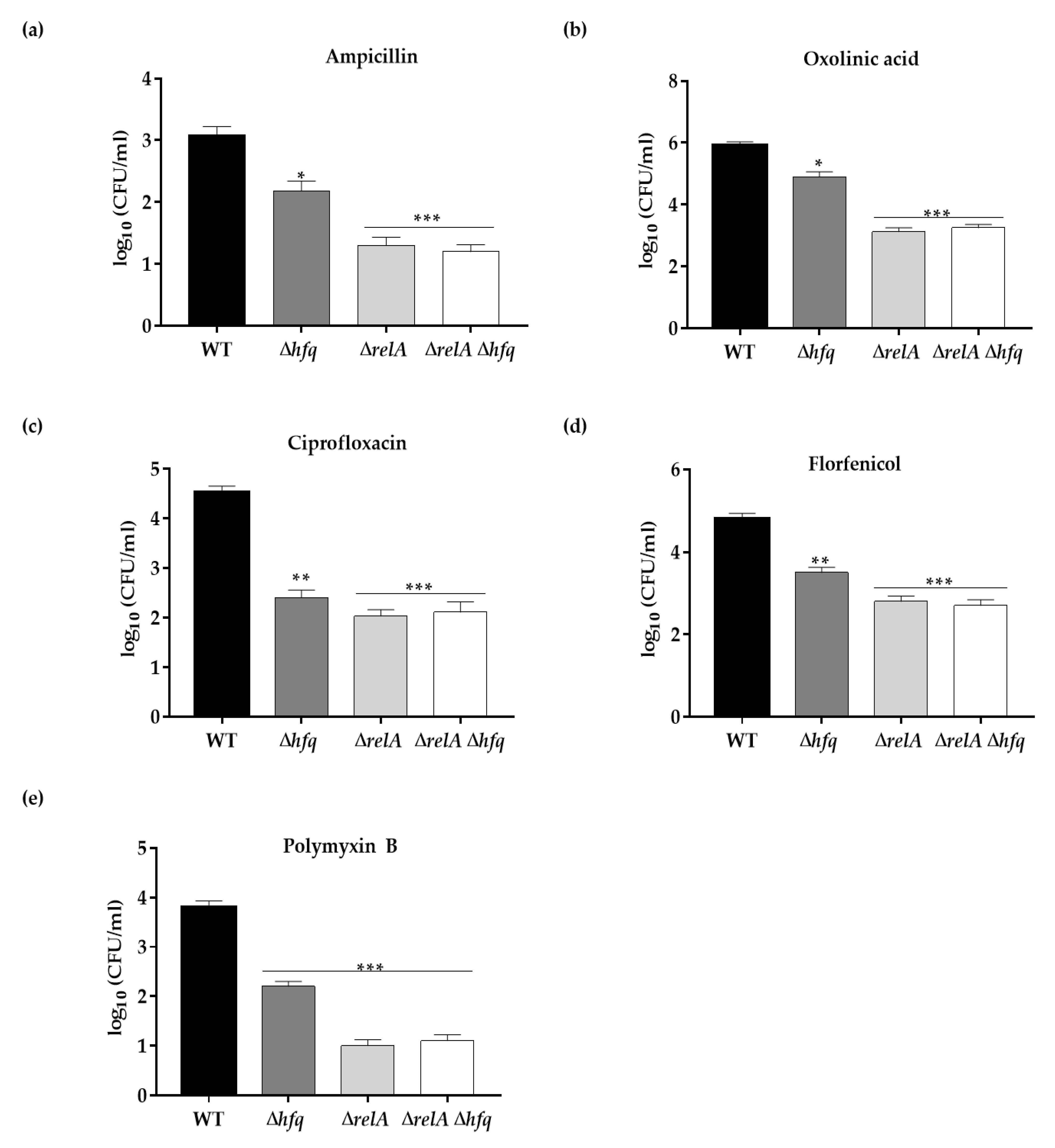
3.3. Hfq Is Required for Antibiotics Persistence in Y. ruckeri
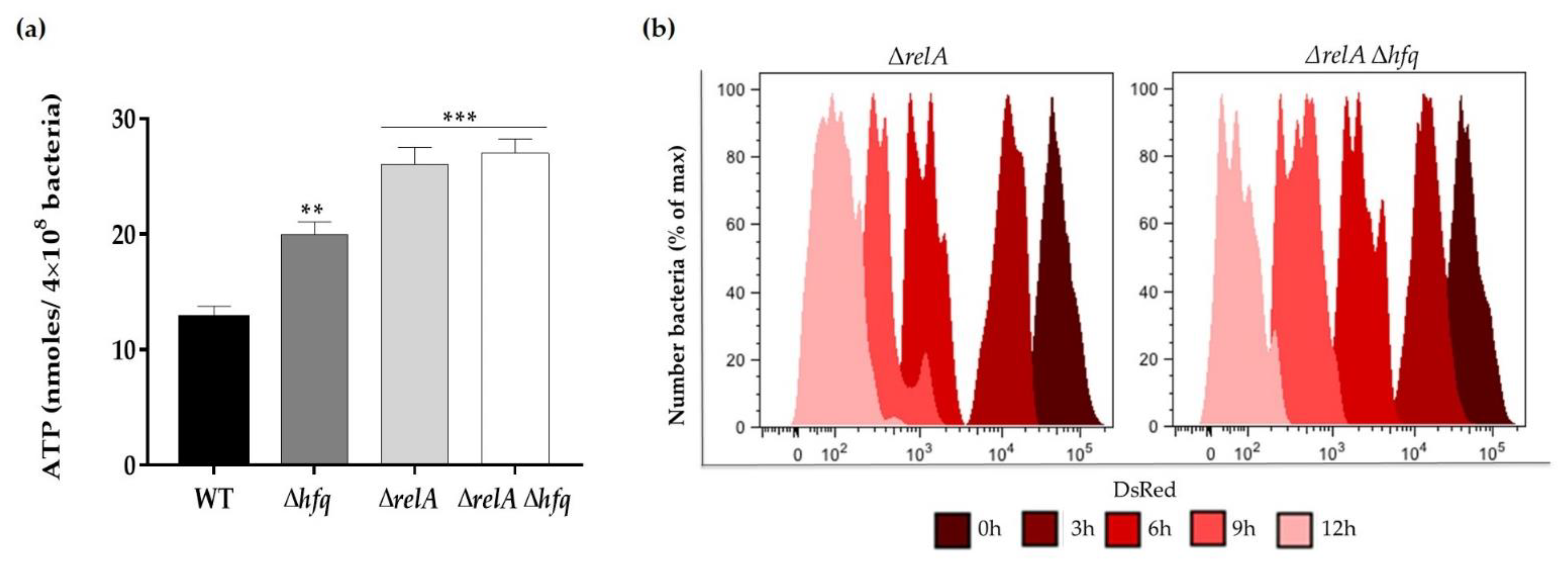
3.4. The Effect of Hfq on Persister Cells Production Is Further Aggravated in the ΔrelA ((p)ppGpp synthetase) Genetic Background
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef]
- Moldoveanu, A.L.; Rycroft, J.A.; Helaine, S. Impact of bacterial persisters on their host. Curr. Opin. Microbiol. 2021, 59, 55–71. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent bacterial infections and persister cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef]
- Salcedo-Sora, J.E.; Kell, D.B. A Quantitative Survey of Bacterial Persistence in the Presence of Antibiotics: Towards Antipersister Antimicrobial Discovery. Antibiotics 2020, 9, 508. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, W.; Tang, Y.; Li, H.; Ma, X.; Liu, Z. RNA chaperone Hfq mediates persistence to multiple antibiotics in Aeromonas veronii. Microb. Pathog. 2019, 132, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Persisters, persistent infections and the Yin-Yang model. Emerg. Microbes Infect. 2014, 3, e3. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, S.; Wu, N.; Yuan, Y.; Zhang, W.; Zhang, Y. Small Non-coding RNA RyhB Mediates Persistence to Multiple Antibiotics and Stresses in Uropathogenic Escherichia coli by Reducing Cellular Metabolism. Front. Microbiol. 2018, 9, 136. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Wood, T.K. Toxins Hha and CspD and small RNA regulator Hfq are involved in persister cell formation through MqsR in Escherichia coli. Biochem. Biophys. Res. Commun. 2010, 391, 209–213. [Google Scholar] [CrossRef] [Green Version]
- Chao, Y.; Vogel, J. The role of Hfq in bacterial pathogens. Curr. Opin. Microbiol. 2010, 13, 24–33. [Google Scholar] [CrossRef]
- Wrobel, A.; Leo, J.C.; Linke, D. Overcoming Fish Defences: The Virulence Factors of Yersinia ruckeri. Genes 2019, 10, 700. [Google Scholar] [CrossRef] [Green Version]
- Guijarro, J.A.; García-Torrico, A.I.; Cascales, D.; Méndez, J. The Infection Process of Yersinia ruckeri: Reviewing the Pieces of the Jigsaw Puzzle. Front. Cell Infect. Microbiol. 2018, 8, 218. [Google Scholar] [CrossRef]
- Bystritskaya, E.; Stenkova, A.; Chistuylin, D.; Chernysheva, N.; Khomenko, V.; Anastyuk, S.; Novikova, O.; Rakin, A.; Isaeva, M. Adaptive responses of outer membrane porin balance of Yersinia ruckeri under different incubation temperature, osmolarity, and oxygen availability. Microbiol. Open 2016, 5, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Michel, C.; Kerouault, B.; Martin, C. Chloramphenicol and florfenicol susceptibility of fish-pathogenic bacteria isolated in France: Comparison of minimum inhibitory concentration, using recommended provisory standards for fish bacteria. J. Appl. Microbiol. 2003, 95, 1008–1015. [Google Scholar] [CrossRef] [PubMed]
- Duman, M.; Altun, S.; Cengiz, M.; Saticioglu, I.B.; Buyukekiz, A.G.; Sahinturk, P. Genotyping and antimicrobial resistance genes of Yersinia ruckeri isolates from rainbow trout farms. Dis. Aquat. Organ. 2017, 125, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Menanteau-Ledouble, S.; Saleh, M.; El-Matbouli, M. Yersinia ruckeri, the causative agent of enteric redmouth disease in fish. Vet. Res. 2015, 46, 103. [Google Scholar] [CrossRef] [Green Version]
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current Status of the Use of Antibiotics and the Antimicrobial Resistance in the Chilean Salmon Farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.C. Resistance of Yersinia ruckeri to antimicrobial agents in vitro. Aquaculture 2001, 196, 325–345. [Google Scholar] [CrossRef]
- Acuña, L.G.; Barros, M.J.; Montt, F.; Peñaloza, D.; Núñez, P.; Valdés, I.; Gil, F.; Fuentes, J.A.; Calderón, I.L. Participation of two sRNA RyhB homologs from the fish pathogen Yersinia ruckeri in bacterial physiology. Microbiol. Res. 2021, 242, 126629. [Google Scholar] [CrossRef]
- Marx, C.I. Development of a broad-host-range sacB-based vector for unmarked allelic exchange. BMC Res. Notes 2008, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Bolivar, F.; Rodriguez, R.L.; Greene, P.J.; Betlach, M.C.; Heyneker, H.L.; Boyer, H.W.; Crosa, J.H.; Falkow, S. Construction and characterization of new cloning vehicles. II. A multipurpose cloning system. Gene 1977, 2, 95–113. [Google Scholar] [CrossRef]
- Helaine, S.; Thompson, J.A.; Watson, K.G.; Liu, M.; Boyle, C.; Holden, D.W. Dynamics of intracellular bacterial replication at the single cell level. Proc. Natl. Acad. Sci. USA 2010, 107, 3746–3751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakoschke, T.; Kakoschke, S.; Magistro, G.; Schubert, S.; Borath, M.; Heesemann, J.; Rossier, O. The RNA chaperone Hfq impacts growth, metabolism and production of virulence factors in Yersinia enterocolitica. PLoS ONE 2014, 9, e86113. [Google Scholar] [CrossRef] [Green Version]
- Vo, P.N.L.; Lee, H.M.; Ren, J.; Na, D. Optimized expression of Hfq protein increases Escherichia coli growth. J. Biol. Eng. 2021, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, Y. PhoU is a persistence switch involved in persister formation and tolerance to multiple antibiotics and stresses in Escherichia coli. Antimicrob. Agents Chemother. 2007, 51, 2092–2099. [Google Scholar] [CrossRef] [Green Version]
- Luidalepp, H.; Jõers, A.; Kaldalu, N.; Tenson, T. Age of inoculum strongly influences persister frequency and can mask effects of mutations implicated in altered persistence. J. Bacteriol. 2011, 193, 3598–3605. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K. Persister cells. Annu. Rev. Microbiol. 2010, 64, 357–372. [Google Scholar] [CrossRef]
- Miyaue, S.; Suzuki, E.; Komiyama, Y.; Kondo, Y.; Morikawa, M.; Maeda, S. Bacterial Memory of Persisters: Bacterial Persister Cells Can Retain Their Phenotype for Days or Weeks After Withdrawal from Colony-Biofilm Culture. Front. Microbiol. 2018, 9, 1396. [Google Scholar] [CrossRef] [Green Version]
- Hauryliuk, V.; Atkinson, G.C.; Murakami, K.S.; Tenson, T.; Gerdes, K. Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 2015, 13, 298–309. [Google Scholar] [CrossRef] [Green Version]
- Pacios, O.; Blasco, L.; Bleriot, I.; Fernandez-Garcia, L.; Ambroa, A.; López, M.; Bou, G.; Cantón, R.; Garcia-Contreras, R.; Wood, T.K.; et al. (p)ppGpp and Its Role in Bacterial Persistence: New Challenges. Antimicrob. Agents Chemother. 2020, 64, e01283-20. [Google Scholar] [CrossRef]
- Calvez, S.; Gantelet, H.; Blanc, G.; Douet, D.G.; Daniel, P. Yersinia ruckeri Biotypes 1 and 2 in France: Presence and antibiotic susceptibility. Dis. Aquat. Organ. 2014, 109, 117–126. [Google Scholar] [CrossRef]
- Huang, Y.; Michael, G.B.; Becker, R.; Kaspar, H.; Mankertz, J.; Schwarz, S.; Runge, M.; Steinhagen, D. Pheno- and genotypic analysis of antimicrobial resistance properties of Yersinia ruckeri from fish. Vet. Microbiol. 2014, 171, 406–412. [Google Scholar] [CrossRef]
- Subpesca. Instituto de Fomento Pesquero (IFOP). Informe Final: Vigilancia de la Resistencia Bacteriana en la Salmonicultura. FIP N°2008-65. 2011. Available online: https://www.subpesca.cl/fipa/613/articles-89266_informe_final.pdf (accessed on 3 May 2021).
- Bai, G.; Golubov, A.; Smith, E.A.; McDonough, K.A. The importance of the small RNA chaperone Hfq for growth of epidemic Yersinia pestis, but not Yersinia pseudotuberculosis, with implications for plague biology. J. Bacteriol. 2010, 192, 4239–4245. [Google Scholar] [CrossRef] [Green Version]
- Padalon-Brauch, G.; Hershberg, R.; Elgrably-Weiss, M.; Baruch, K.; Rosenshine, I.; Margalit, H.; Altuvia, S. Small RNAs encoded within genetic islands of Salmonella typhimurium show host-induced expression and role in virulence. Nucleic Acids Res. 2008, 36, 1913–1927. [Google Scholar] [CrossRef] [PubMed]
- Peñaloza, D.; Acuña, L.G.; Barros, M.J.; Núñez, P.; Montt, F.; Gil, F.; Fuentes, J.A.; Calderón, I.L. The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages. Microorganisms 2021, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Tsui, H.C.; Leung, H.C.; Winkler, M.E. Characterization of broadly pleiotropic phenotypes caused by an hfq insertion mutation in Escherichia coli K-12. Mol. Microbiol. 1994, 13, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Sittka, A.; Pfeiffer, V.; Tedin, K.; Vogel, J. The RNA chaperone Hfq is essential for the virulence of Salmonella typhimurium. Mol. Microbiol. 2007, 63, 193–217. [Google Scholar] [CrossRef] [Green Version]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 2016, 1, 16051. [Google Scholar] [CrossRef] [Green Version]
- Shan, Y.; Brown Gandt, A.; Rowe, S.E.; Deisinger, J.P.; Conlon, B.P.; Lewis, K. ATP-Dependent Persister Formation in Escherichia coli. mBio 2017, 8, e02267-16. [Google Scholar] [CrossRef] [Green Version]
- Pusic, P.; Sonnleitner, E.; Krennmayr, B.; Heitzinger, D.A.; Wolfinger, M.T.; Resch, A.; Bläsi, U. Harnessing Metabolic Regulation to Increase Hfq-Dependent Antibiotic Susceptibility in Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 2709. [Google Scholar] [CrossRef] [Green Version]
- Yamada, J.; Yamasaki, S.; Hirakawa, H.; Hayashi-Nishino, M.; Yamaguchi, A.; Nishino, K. Impact of the RNA chaperone Hfq on multidrug resistance in Escherichia coli. J. Antimicrob. Chemother. 2010, 65, 853–858. [Google Scholar] [CrossRef]
- Hayashi-Nishino, M.; Fukushima, A.; Nishino, K. Impact of Hfq on the intrinsic drug resistance of Salmonella enterica serovar typhimurium. Front. Microbiol. 2012, 3, 205. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.C.; Chien, H.F.; Tsai, Y.L.; Liu, M.C.; Liaw, S.J. The RNA chaperone Hfq is involved in stress tolerance and virulence in uropathogenic Proteus mirabilis. PLoS ONE 2014, 9, e85626. [Google Scholar] [CrossRef] [PubMed]
- Dersch, P.; Khan, M.A.; Mühlen, S.; Görke, B. Roles of Regulatory RNAs for Antibiotic Resistance in Bacteria and Their Potential Value as Novel Drug Targets. Front. Microbiol. 2017, 8, 803. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.F.; Han, K.; Chandler, C.E.; Tjaden, B.; Ernst, R.K.; Lory, S. Probing the sRNA regulatory landscape of P. aeruginosa: Post-transcriptional control of determinants of pathogenicity and antibiotic susceptibility. Mol. Microbiol. 2017, 106, 919–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, B.W.; Valenta, J.A.; Benedik, M.J.; Wood, T.K. Arrested protein synthesis increases persister-like cell formation. Antimicrob. Agents Chemother. 2013, 57, 1468–1473. [Google Scholar] [CrossRef] [Green Version]
- Allison, K.R.; Brynildsen, M.P.; Collins, J.J. Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature 2011, 473, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Korch, S.B.; Henderson, T.A.; Hill, T.M. Characterization of the hipA7 allele of Escherichia coli and evidence that high persistence is governed by (p)ppGpp synthesis. Mol. Microbiol. 2003, 50, 1199–1213. [Google Scholar] [CrossRef]
- Svenningsen, M.S.; Veress, A.; Harms, A.; Mitarai, N.; Semsey, S. Birth and Resuscitation of (p)ppGpp Induced Antibiotic Tolerant Persister Cells. Sci. Rep. 2019, 9, 6056. [Google Scholar] [CrossRef]
- Argaman, L.; Elgrably-Weiss, M.; Hershko, T.; Vogel, J.; Altuvia, S. RelA protein stimulates the activity of RyhB small RNA by acting on RNA-binding protein Hfq. Proc. Natl. Acad. Sci. USA 2012, 109, 4621–4626. [Google Scholar] [CrossRef] [Green Version]
- Basu, P.; Elgrably-Weiss, M.; Hassouna, F.; Kumar, M.; Wiener, R.; Altuvia, S. RNA binding of Hfq monomers promotes RelA-mediated hexamerization in a limiting Hfq environment. Nat. Commun. 2021, 12, 2249. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Characteristics | Reference |
|---|---|---|
| Y. ruckeri CD2 | Wild-type strain of Y. ruckeri | [18] |
| Δhfq | Y. ruckeri CD2 lacking hfq gene | [18] |
| ΔrelA | Y. ruckeri CD2 lacking relA gene | This study |
| ΔrelA Δhfq | Y. ruckeri CD2 lacking realA and hfq genes | This study |
| Plasmid | Genotype | Reference |
| pCM433 | ApR, CmR, TcR, oriT, tet, cat, sacB and ColEI ori; broad-host-range sacB-based allelic exchange vector | [19] |
| pCM433-hfq | hfq region of Y. ruckeri CD2 cloned into pCM433 | This study |
| pCM433-relA | relA region of Y. ruckeri CD2 cloned into pCM433 | This study |
| pBR322 | ApR, TcR, ColEl Ori. | [20] |
| pHfq | hfq region of Y. ruckeri CD2 cloned into pBR322 | This study |
| pDiGc | bla GFP pBAD DsRed ori f1 AmpR | [21] |
| Antibiotic | MIC (mg mL−1) | MBC (mg mL−1) | ||||
|---|---|---|---|---|---|---|
| WT | Δhfq | pHfq | WT | Δhfq | pHfq | |
| Ampicillin | 1 | 0.1 | 1 | 1 | 0.1 | 1 |
| Oxolinic acid | 0.005 | 0.0005 | 0.005 | 0.05 | 0.5 | 0.05 |
| Ciprofloxacin | 0.003 | 0.003 | 0.003 | 0.003 | 0.3 | 0.003 |
| Florfenicol | 0.01 | 0.01 | 0.01 | 0.01 | 0.1 | 0.01 |
| Polymyxin B | 2 | 0.2 | 2 | 2 | 0.2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderón, I.L.; Barros, M.J.; Montt, F.; Gil, F.; Fuentes, J.A.; Acuña, L.G. The RNA Chaperone Hfq Participates in Persistence to Multiple Antibiotics in the Fish Pathogen Yersinia ruckeri. Microorganisms 2021, 9, 1404. https://doi.org/10.3390/microorganisms9071404
Calderón IL, Barros MJ, Montt F, Gil F, Fuentes JA, Acuña LG. The RNA Chaperone Hfq Participates in Persistence to Multiple Antibiotics in the Fish Pathogen Yersinia ruckeri. Microorganisms. 2021; 9(7):1404. https://doi.org/10.3390/microorganisms9071404
Chicago/Turabian StyleCalderón, Iván L., María José Barros, Fernanda Montt, Fernando Gil, Juan A. Fuentes, and Lillian G. Acuña. 2021. "The RNA Chaperone Hfq Participates in Persistence to Multiple Antibiotics in the Fish Pathogen Yersinia ruckeri" Microorganisms 9, no. 7: 1404. https://doi.org/10.3390/microorganisms9071404