
A New Thermophilic Ene-Reductase from the Filamentous Anoxygenic Phototrophic Bacterium Chloroflexus aggregans

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sequence Analysis and Cloning
2.2. Expression, Analysis and Purification of Recombinant Protein
2.3. Native Gel and NBT Staining
2.4. Analytical Gel Filtration
2.5. Thermofluor Measurements
2.6. Activity Assay and Kinetics
2.7. Determination of pH Optimum
2.8. Crystallization and Data Collection
2.9. Model Building and Refinement
3. Results
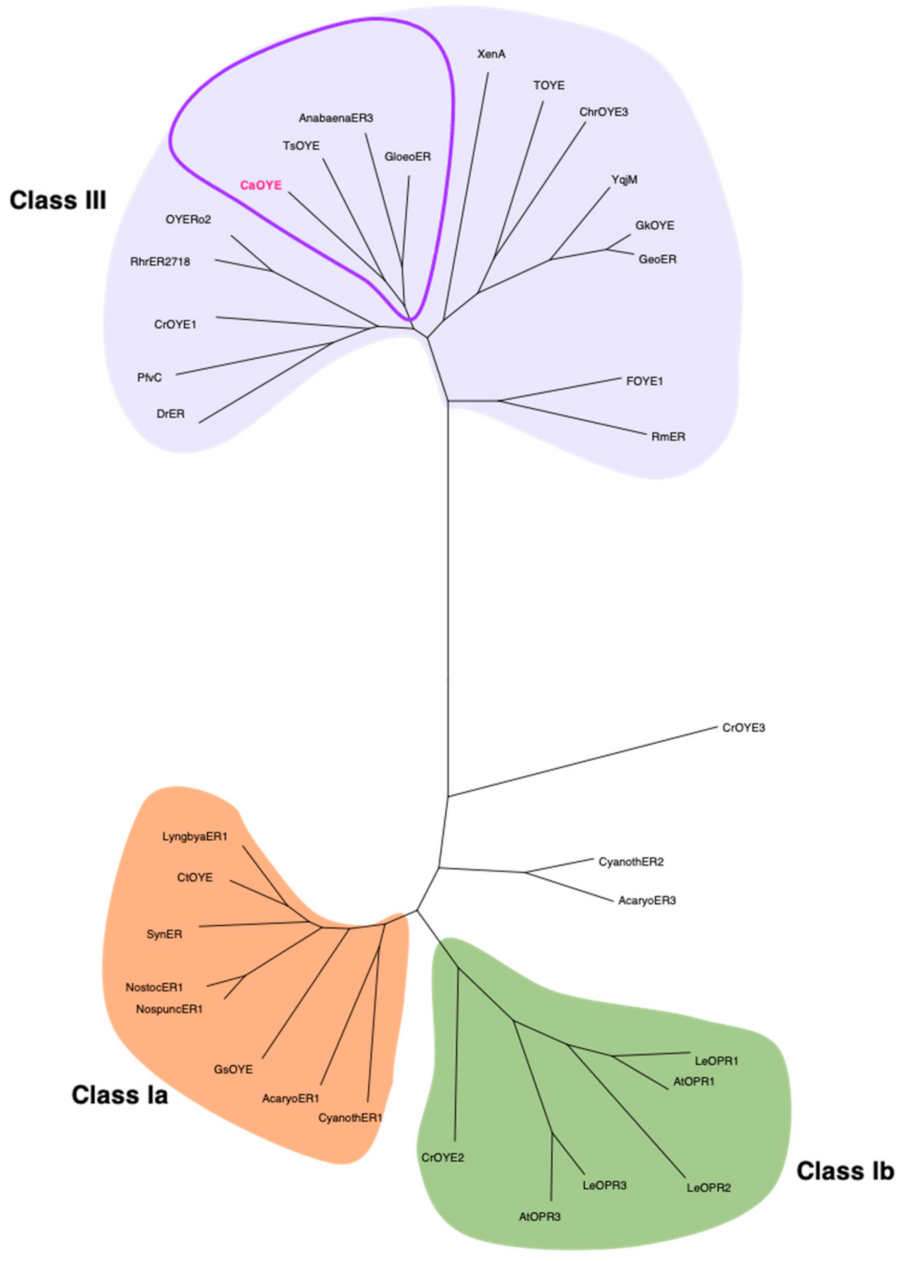
3.1. Identification of New Putative CaOYE and Sequence Analysis
3.2. Purification of Recombinant CaOYE and Evaluation of Its Oligomeric State
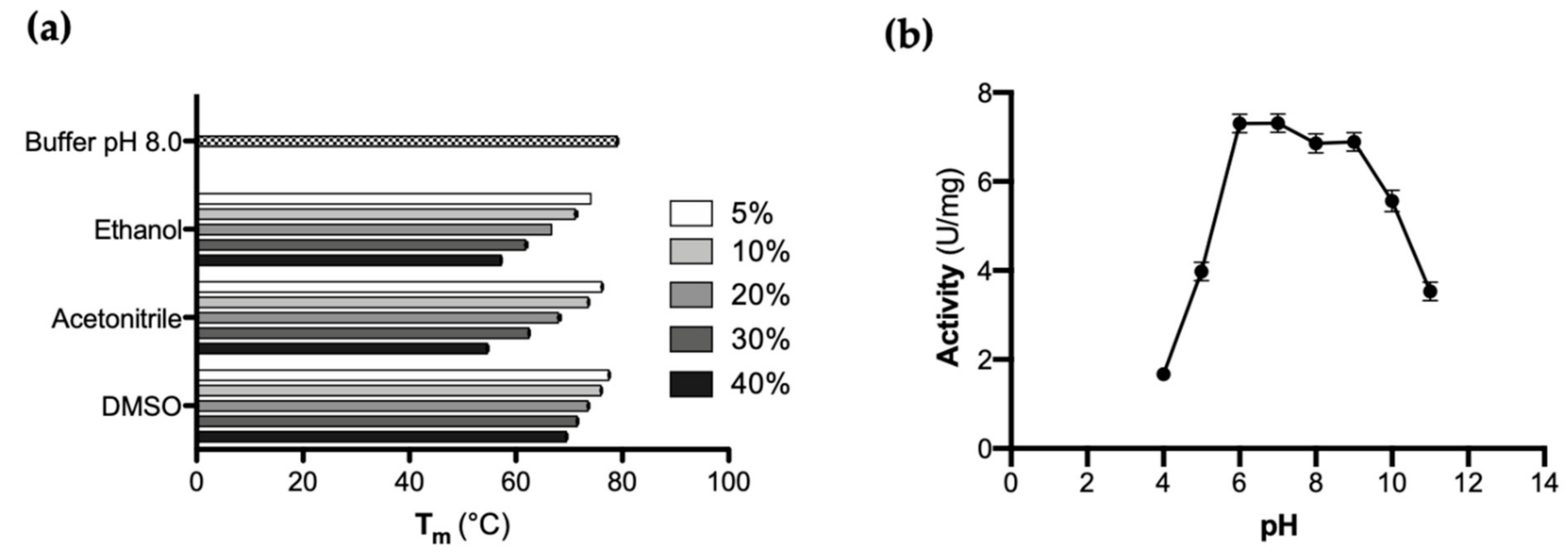
3.3. Evaluation of CaOYE Thermal Stability and pH Optimum
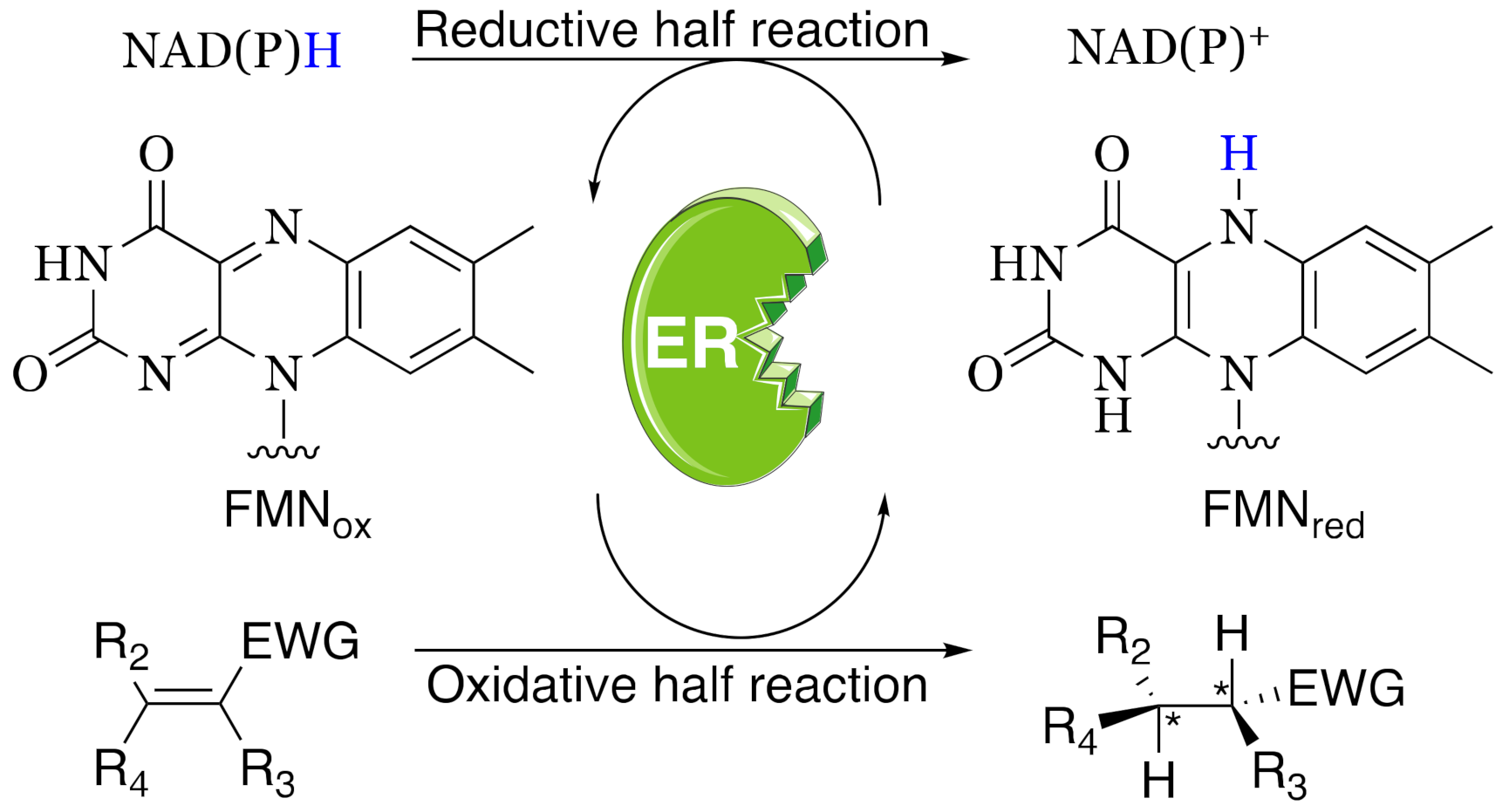
3.4. Evaluation of CaOYE Activity with Standard Substrates and Steady-State Kinetic Parameters
3.5. CaOYE Crystal Structure
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biotechnol. 2015, 99, 7907–7913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholtissek, A.; Tischler, D.; Westphal, A.H.; van Berkel, W.J.H.; Paul, C.E. Old Yellow Enzyme-Catalysed asymmetric hydrogenation: Linking family roots with improved catalysis. Catalysts 2017, 7, 130. [Google Scholar] [CrossRef]
- Williams, R.E.; Bruce, N.C. ‘New uses for an old enzyme’—The Old Yellow Enzyme family of flavoenzymes. Microbiology 2002, 148, 1607–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaller, F.; Biesgen, C.; Mussig, C.; Altmann, T.; Weiler, E.W. 12-Oxophytodienoate reductase 3 (OPR3) is the isoenzyme involved in jasmonate biosynthesis. Planta 2000, 210, 979–984. [Google Scholar] [CrossRef]
- Cheng, J.Z.; Coyle, C.M.; Panaccione, D.G.; O’Connor, S.E. A role for Old Yellow Enzyme in ergot alkaloid biosynthesis. J. Am. Chem. Soc. 2010, 132, 1776–1777. [Google Scholar] [CrossRef]
- Diaz-Viraque, F.; Chiribao, M.L.; Tronchine, A.; Gonzalez-Herrera, F.; Castillo, C.; Liempi, A.; Kemmerling, U.; Maya, J.D.; Robello, C. Old Yellow Enzyme from Trypanosoma cruzi exhibits in vivo prostaglandin F2α synthase activity and has a key role in parasite infection and drug susceptibility. Front. Immunol. 2018, 9, 456. [Google Scholar] [CrossRef] [Green Version]
- Toogood, H.S.; Scrutton, N.S. Discovery, characterization, engineering, and applications of ene-reductases for industrial biocatalysis. ACS Catal. 2018, 8, 3532–3549. [Google Scholar] [CrossRef]
- Hall, M.; Yanto, Y.; Bommarius, A.S. Old Yellow Enzyme family and Enoate Reductases. In The Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology; Flickinger, M.C., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2010; pp. 2234–2247. [Google Scholar]
- Durchschein, K.; Hall, M.; Faber, K. Unusual reactions mediated by FMN-dependent ene- and nitro-reductases. Green Chem. 2013, 15, 1764. [Google Scholar] [CrossRef] [Green Version]
- Turrini, N.G.; Eger, E.; Reiter, T.C.; Faber, K.; Hall, M. Sequential enzymatic conversion of alpha-angelica lactone to gamma-valerolactone through hydride-independent C=C bond isomerization. ChemSusChem 2016, 9, 3393–3396. [Google Scholar] [CrossRef]
- Sandoval, B.A.; Meichan, A.J.; Hyster, T.K. Enantioselective hydrogen atom transfer: Discovery of catalytic promiscuity in flavin-dependent ‘ene’-reductases. J. Am. Chem. Soc. 2017, 139, 11313–11316. [Google Scholar] [CrossRef]
- Sandoval, B.A.; Kurtoic, S.I.; Chung, M.M.; Biegasiewicz, K.F.; Hyster, T.K. Photoenzymatic catalysis enables radical-mediated ketone reduction in ene-reductases. Angew. Chem. Int. Ed. 2019, 58, 8714–8718. [Google Scholar] [CrossRef]
- Biegasiewicz, K.F.; Cooper, S.J.; Gao, X.; Oblinsky, D.G.; Kim, J.H.; Garfinkle, S.E.; Joyce, L.A.; Sandoval, B.A.; Scholes, G.D.; Hyster, T.K. Photoexcitation of flavoenzymes enables a stereoselective radical cyclization. Science 2019, 364, 1166–1169. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, B.; Wang, Y.; Jiang, G.; Feng, J.; Zhao, H. Photoenzymatic enantioselective intermolecular radical hydroalkylation. Nature 2020, 584, 69–74. [Google Scholar] [CrossRef]
- Adalbjornsson, B.V.; Toogood, H.S.; Fryszkowska, A.; Pudney, C.R.; Jowitt, T.A.; Leys, D.; Scrutton, N.S. Biocatalysis with thermostable enzymes: Structure and properties of a thermophilic ‘ene’-reductase related to Old Yellow Enzyme. ChemBioChem 2010, 11, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Opperman, D.J.; Piater, L.A.; van Heerden, E. A novel chromate reductase from Thermus scotoductus SA-01 related to Old Yellow Enzyme. J. Bacteriol. 2008, 190, 3076–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schittmayer, M.; Glieder, A.; Uhl, M.K.; Winkler, A.; Zach, S.; Schrittwieser, J.H.; Kroutil, W.; Macheroux, P.; Gruber, K.; Kambourakis, S.; et al. Old Yellow Enzyme-Catalyzed dehydrogenation of saturated ketones. Adv. Synth. Catal. 2011, 353, 268–274. [Google Scholar] [CrossRef]
- Tsuji, N.; Honda, K.; Wada, M.; Okano, K.; Ohtake, H. Isolation and characterization of a thermotolerant ene-reductase from Geobacillus sp. 30 and its heterologous expression in Rhodococcus opacus. Appl. Microbiol. Biotechnol. 2014, 98, 5925–5935. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.-Y.; Pei, X.-Q.; Wu, Z.-L. Identification and characterization of a novel “thermophilic-like” Old Yellow Enzyme from the genome of Chryseobacterium sp. CA49. J. Mol. Catal. B Enzym. 2014, 108, 64–71. [Google Scholar] [CrossRef]
- Scholtissek, A.; Ullrich, S.R.; Muhling, M.; Schlomann, M.; Paul, C.E.; Tischler, D. A thermophilic-like ene-reductase originating from an acidophilic iron oxidizer. Appl. Microbiol. Biotechnol. 2017, 101, 609–619. [Google Scholar] [CrossRef]
- Robescu, M.S.; Niero, M.; Hall, M.; Cendron, L.; Bergantino, E. Two new ene-reductases from photosynthetic extremophiles enlarge the panel of Old Yellow Enzymes: CtOYE and GsOYE. Appl. Microbiol. Biotechnol. 2020, 104, 2051–2066. [Google Scholar] [CrossRef]
- Peters, C.; Frasson, D.; Sievers, M.; Buller, R. Novel Old Yellow Enzyme subclasses. ChemBioChem 2019, 20, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Böhmer, S.; Marx, C.; Gomez-Baraibar, A.; Nowaczyk, M.M.; Tischler, D.; Hemschemeier, A.; Happe, T. Evolutionary diverse Chlamydomonas reinhardtii Old Yellow Enzymes reveal distinctive catalytic properties and potential for whole-cell biotransformations. Algal Res. 2020, 50, 101970. [Google Scholar] [CrossRef]
- Magallanes-Noguera, C.; Cecati, F.M.; Mascotti, M.L.; Reta, G.F.; Agostini, E.; Orden, A.A.; Kurina-Sanz, M. Plant tissue cultures as sources of new ene- and ketoreductase activities. J. Biotechnol. 2017, 251, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strassner, J.; Furholz, A.; Macheroux, P.; Amrhein, N.; Schaller, A. A homolog of Old Yellow Enzyme in tomato: Spectral properties and substrate specificity of the recombinant protein. J. Biol. Chem. 1999, 274, 35067–35073. [Google Scholar] [CrossRef] [Green Version]
- Strassner, J.; Schaller, F.; Frick, U.B.; Howe, G.A.; Weiler, E.W.; Amrhein, N.; Macheroux, P.; Schaller, A. Characterization and cDNA-microarray expression analysis of 12-oxophytodienoate reductases reveals differential roles for octadecanoid biosynthesis in the local versus the systemic wound response. Plant J. 2002, 32, 17. [Google Scholar] [CrossRef]
- Fu, Y.; Castiglione, K.; Weuster-Botz, D. Comparative characterization of novel ene-reductases from Cyanobacteria. Biotechnol. Bioeng. 2013, 110, 1293–1301. [Google Scholar] [CrossRef]
- Fu, Y.; Hoelsch, K.; Weuster-Botz, D. A novel ene-reductase from Synechococcus sp. PCC 7942 for the asymmetric reduction of alkenes. Process Biochem. 2012, 47, 1988–1997. [Google Scholar] [CrossRef]
- Hanada, S.; Hiraishi, A.; Shimada, K.; Matsuura, K. Chloroflexus aggregans sp. nov., a filamentous phototrophic bacterium which forms dense cell aggregates by active gliding movement. Int. J. Syst. Bacteriol. 1995, 45, 676–681. [Google Scholar] [CrossRef] [Green Version]
- Kanno, N.; Haruta, S.; Hanada, S. Sulfide-dependent photoautotrophy in the filamentous anoxygenic phototrophic bacterium, Chloroflexus aggregans. Microbes Environ. 2019, 34, 304–309. [Google Scholar] [CrossRef] [Green Version]
- Kawai, S.; Nishihara, A.; Matsuura, K.; Haruta, S. Hydrogen-dependent autotrophic growth in phototrophic and chemolithotrophic cultures of thermophilic bacteria, Chloroflexus aggregans and Chloroflexus aurantiacus, isolated from Nakabusa hot springs. FEMS Microbiol. Lett. 2019, 366, fnz122. [Google Scholar] [CrossRef] [Green Version]
- Kabsch, W. XDS. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.W.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Murshudov, G.N.; Skubák, P.; Lebedev, A.A.; Pannu, N.S.; Steiner, R.A.; Nicholls, R.A.; Winn, M.D.; Long, F.; Vagin, A.A. REFMAC 5 for the refinement of macromolecular crystal structures. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 355–367. [Google Scholar] [CrossRef] [Green Version]
- Afonine, P.V.; Grosse-Kunstleve, R.W.; Echols, N.; Headd, J.J.; Moriarty, N.W.; Mustyakimov, M.; Terwilliger, T.C.; Urzhumtsev, A.; Zwart, P.H.; Adams, P.D. Towards automated crystallographic structure refinement with phenix.refine. Acta Crystallogr. D Biol. Crystallogr. 2012, 68, 352–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberdorfer, G.; Steinkellner, G.; Stueckler, C.; Faber, K.; Gruber, K. Stereopreferences of Old Yellow Enzymes: Structure correlations and sequence patterns in enoate reductases. ChemCatChem 2011, 3, 1562–1566. [Google Scholar] [CrossRef]
- Gaisin, V.A.; Kooger, R.; Grouzdev, D.S.; Gorlenko, V.M.; Pilhofer, M. Cryo-electron tomography reveals the complex ultrastructural organization of multicellular filamentous Chloroflexota (Chloroflexi) bacteria. Front. Microbiol. 2020, 11, 1373. [Google Scholar] [CrossRef] [PubMed]
- Kawai, S.; Martinez, J.N.; Lichtenberg, M.; Trampe, E.; Kühl, M.; Tank, M.; Haruta, S.; Nishihara, A.; Hanada, S.; Thiel, V. In-situ metatranscriptomic analyses reveal the metabolic flexibility of the thermophilic anoxygenic photosynthetic bacterium Chloroflexus aggregans in a hot spring cyanobacteria-dominated microbial mat. Microorganisms 2021, 9, 652. [Google Scholar] [CrossRef]
- Mimuro, M.; Tomo, T.; Tsuchiya, T. Two unique cyanobacteria lead to a traceable approach of the first appearance of oxygenic photosynthesis. Photosynth. Res. 2008, 97, 167–176. [Google Scholar] [CrossRef]
- Thiel, T.; Pratte, B.S.; Zhong, J.; Goodwin, L.; Copeland, A.; Lucas, S.; Han, C.; Pitluck, S.; Land, M.L.; Kyrpides, N.C.; et al. Complete genome sequence of Anabaena variabilis ATCC 29413. Stand. Genom. Sci. 2014, 9, 562–573. [Google Scholar] [CrossRef] [Green Version]
- Gounder, K.; Brzuszkiewicz, E.; Liesegang, H.; Wollherr, A.; Daniel, R.; Gottschalk, G.; Reva, O.; Kumwenda, B.; Srivastava, M.; Bricio, C.; et al. Sequence of the hyperplastic genome of the naturally competent Thermus scotoductus SA-01. BMC Genom. 2011, 12, 577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Litthauer, S.; Gargiulo, S.; van Heerden, E.; Hollmann, F.; Opperman, D.J. Heterologous expression and characterization of the ene-reductases from Deinococcus radiodurans and Ralstonia metallidurans. J. Mol. Catal. B Enzym. 2014, 99, 89–95. [Google Scholar] [CrossRef]
- Opperman, D.J. Structural investigation into the C-terminal extension of the ene-reductase from Ralstonia (Cupriavidus) metallidurans. Proteins 2017, 85, 2252–2257. [Google Scholar] [CrossRef] [PubMed]
- Riedel, A.; Tischler, D. Functional characterization and stability improvement of a ‘thermophilic-like’ ene-reductase from Rhodococcus opacus 1CP. Front. Microbiol. 2015, 6, 1073. [Google Scholar] [CrossRef] [Green Version]
- Opperman, D.J.; Sewell, B.T.; Litthauer, D.; Isupov, M.N.; Littlechild, J.A. Crystal structure of a thermostable Old Yellow Enzyme from Thermus scotoductus SA-01. Biochem. Biophys. Res. Commun. 2010, 393, 426–431. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Specific Activity | Kinetic Parameters | |||
|---|---|---|---|---|---|
| Structure | Name | U/mg | Km (mM) | kcat (s−1) | kcat/Km (mM−1 s−1) |
 | 2-Cyclopenten-1-one | 1.1 ± 0.09 | N.D. | N.D. | N.D. |
 | 2-Cyclohexen-1-one | 5.25 ± 0.22 | 5.520 ± 0.627 | 3.82 ± 0.176 | 0.69 |
 | 2-Methyl-cyclohexen-1-one | 1.19 ± 0.07 | N.D. | N.D. | N.D. |
 | 4-Ketoisophorone | 8.3 ± 0.50 | 0.013 ± 0.002 | 2.27 ± 0.055 | 174.6 |
 | Maleimide | 2.2 ± 0.37 | N.D. | N.D. | N.D. |
 | 2-Methyl-pentenal | 2.0 ± 0.15 | N.D. | N.D. | N.D. |
 | trans-2-Hexen-1-al | 1.1 ± 0.09 | N.D. | N.D. | N.D. |
 | 1-Octen-3-one | 9.4 ± 0.40 | 10.32 ± 1.438 | 12.13 ± 0.661 | 1.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robescu, M.S.; Niero, M.; Loprete, G.; Cendron, L.; Bergantino, E. A New Thermophilic Ene-Reductase from the Filamentous Anoxygenic Phototrophic Bacterium Chloroflexus aggregans. Microorganisms 2021, 9, 953. https://doi.org/10.3390/microorganisms9050953
Robescu MS, Niero M, Loprete G, Cendron L, Bergantino E. A New Thermophilic Ene-Reductase from the Filamentous Anoxygenic Phototrophic Bacterium Chloroflexus aggregans. Microorganisms. 2021; 9(5):953. https://doi.org/10.3390/microorganisms9050953
Chicago/Turabian StyleRobescu, Marina Simona, Mattia Niero, Giovanni Loprete, Laura Cendron, and Elisabetta Bergantino. 2021. "A New Thermophilic Ene-Reductase from the Filamentous Anoxygenic Phototrophic Bacterium Chloroflexus aggregans" Microorganisms 9, no. 5: 953. https://doi.org/10.3390/microorganisms9050953