
Antifungal Activity of Propyl Disulfide from Neem (Azadirachta indica) in Vapor and Agar Diffusion Assays against Anthracnose Pathogens (Colletotrichum gloeosporioides and Colletotrichum acutatum) in Mango Fruit

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemicals
2.2. Isolation and Identification of Fungi
2.3. Pathogenicity Test
2.4. Antifungal Activity of Propyl Disulfide
2.5. Gompertz Model for Fungal Growth
2.6. Statistical Analysis
3. Results
3.1. Anthracnose Rot and Characteristics of Isolated Fungi
3.2. Antifungal Activity of Propyl Disulfide
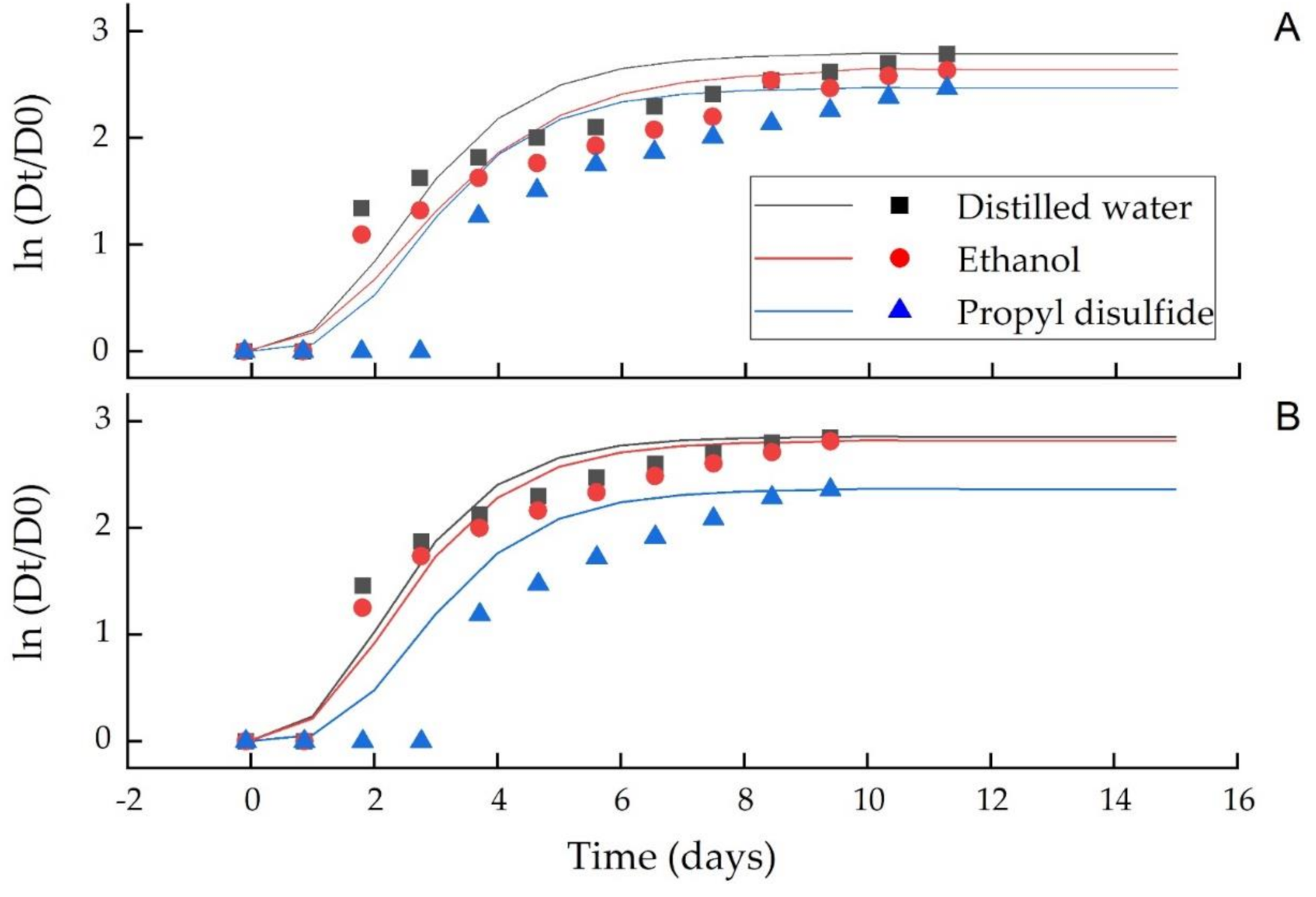
3.3. Gompertz Model for Fungal Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, X.; Lei, H.; Ma, X.; Lai, T.; Song, H.; Shi, X.; Li, J. Antifungal activity of 1-methylcyclopropene (1-MCP) against anthracnose (Colletotrichum gloeosporioides) in post-harvest mango fruit and its possible mechanisms of action. Int. J. Food Microbiol. 2017, 241, 1–6. [Google Scholar] [CrossRef]
- Konsue, W.; Dethoup, T.; Limtong, S. Biological control of fruit rot and anthracnose of postharvest mango by antagonistic yeasts from economic crops leaves. Microorganisms 2020, 8, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, G.; Zhang, X.S.; Zhang, Z.K.; Ye, H.C.; Liu, Y.Q.; Yang, G.Z.; Chen, C.; Chen, M.; Yan, C.; Wang, L.Y.; et al. Fungicidal activities of camptothecin semisynthetic derivatives against Colletotrichum gloeosporioides in vitro and in mango fruit. Postharvest Biol. Technol. 2019, 147, 139–147. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, D.; Yang, B.; Gao, Z.; Li, M.; Jiang, Y.; Hu, M. β-Aminobutyric acid induces resistance of mango fruit to post-harvest anthracnose caused by Colletotrichum gloeosporioides and enhances activity of fruit defense mechanisms. Sci. Hortic. 2013, 160, 78–84. [Google Scholar] [CrossRef]
- Hu, M.; Yang, D.; Huber, D.J.; Jiang, Y.; Li, M.; Gao, Z.; Zhang, Z. Reduction of post-harvest anthracnose and enhancement of disease resistance in ripening mango fruit by nitric oxide treatment. Postharvest Biol. Technol. 2014, 97, 115–122. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, L.; Liu, Y.; Xu, N.; Zhou, S.; Yang, Q.; Yang, Y.; Ai, X. Thymol protects channel catfish from Aeromonas hydrophila Infection by inhibiting aerolysin expression and biofilm formation. Microorganisms 2020, 8, 636. [Google Scholar] [CrossRef]
- Topa, S.H.; Palombo, E.A.; Kingshott, P.; Blackall, L.L. Activity of cinnamaldehyde on quorum sensing and biofilm susceptibility to antibiotics in pseudomonas aeruginosa. Microorganisms 2020, 8, 455. [Google Scholar] [CrossRef] [Green Version]
- Tzortzakis, N.G. Methyl jasmonate-induced suppression of anthracnose rot in tomato fruit. Crop Prot. 2007, 26, 1507–1513. [Google Scholar] [CrossRef]
- Abdelhamid, A.G.; Yousef, A.E. Natural antimicrobials suitable for combating desiccation-resistant Salmonella enterica in milk powder. Microorganisms 2021, 9, 421. [Google Scholar] [CrossRef]
- Munekata, P.E.S.; Pateiro, M.; Rodríguez-Lázaro, D.; Domínguez, R.; Zhong, J.; Lorenzo, J.M. The role of essential oils against pathogenic Escherichia coli in food products. Microorganisms 2020, 8, 924. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kudachikar, V. Efficacy of aroma compounds for post-harvest management of mango Anthracnose. J. Plant Dis. Prot. 2019. [Google Scholar] [CrossRef]
- De Rima Oliveira, K.A.; Berger, L.R.R.; De Amorim Araujo, S.; Camara, M.P.S.; De Leite Souza, E. Synergistic mixtures of chitosan and Mentha piperita L. essential oil to inhibit Colletotrichum species and anthracnose development in mango cultivar Tommy Atkins. Food Microbiol. 2017, 66, 96–103. [Google Scholar] [CrossRef]
- Oliveira, P.D.L.; De Rimá Oliveira, K.Á.; Vieira, W.A.S.; Câmara, M.P.S.; De Leite Souza, E. Control of anthracnose caused by Colletotrichum species in guava, mango and papaya using synergistic combinations of chitosan and Cymbopogon citratus (D.C. ex Nees) Stapf. essential oil. Int. J. Food Microbiol. 2018, 266, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Linde, J.H.; Combrinck, S.; Regnier, T.J.C.; Virijevic, S. Chemical composition and antifungal activity of the essential oils of Lippia rehmannii from South Africa. S. Afr. J. Bot. 2010, 76, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Dessalegn, Y.; Ayalew, A.; Woldetsadik, K. Integrating plant defense inducing chemical, inorganic salt and hot water treatments for the management of post-harvest mango anthracnose. Postharvest Biol. Technol. 2013, 85, 83–88. [Google Scholar] [CrossRef]
- Nicoletti, M.; Murugan, K. Neem the tree of 21st century. Pharmacologyonline 2013, 3, 115–121. [Google Scholar]
- Kumar, D.; Rahal, A.; Malik, K.J. Neem extract. In Nutraceuticals. Efficacy, Safety and Toxicity; Ramesh, C., Ed.; Gupta: Chennai, India, 2016; pp. 585–597. [Google Scholar] [CrossRef]
- Hossain, M.A.; Al-Toubi, W.A.S.; Weli, A.M.; Al-Riyami, Q.A.; Al-Sabahi, J.N. Identification and characterization of chemical compounds indifferent crude extracts from leaves of Omani neem. J. Taibah Uni. Sci. 2013, 7, 181–188. [Google Scholar] [CrossRef]
- Serrone, P.D.; Toniolo, C.; Nicoletti, M. Neem (Azadirachta indica A. Juss) oil: A natural preservative to control meat spoilage. Foods 2015, 4, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Zakawa, N.N.; Channya, K.F.; Magga, B.; Akesa, T.M. Antifungal effect of neem (Azadirachta indica) leaf extracts on mango fruit post-harvest rot agents in Yola, Adamawa state. J. Pharmacogn. Phytochem. 2018, 7, 23–26. [Google Scholar]
- Suleiman, M.N. Antifungal properties of leaf extract of neem and tobacco on three fungal pathogens of tomato (Lycopersicon Esculentum Mill). Adv. Appl. Sci. Res. 2011, 2, 217–220. [Google Scholar]
- Khan, M.R.; Suwanamornlert, P.; Sangchote, S.; Chonhenchob, V. Antifungal activity of propyl disulphide from neem against Lasiodiplodia theobromae and Neofusicoccum parvum causing stem end rot in mango. J. Appl. Microbiol. 2020, 129, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Wanjiku, E.K.; Waceke, J.W.; Wanjala, B.W.; Mbaka, J.N. Identification and pathogenicity of fungal pathogens associated with stem end rots of avocado fruits in kenya. Int. J. Microbiol. 2020. [Google Scholar] [CrossRef]
- Karabıçak, N.; Karatuna, O.; Akyar, I. Evaluation of the viabilities and stabilities of pathogenic mold and yeast species using three different preservation methods over a 12-year period along with a review of published reports. Mycopathologia 2016. [Google Scholar] [CrossRef] [PubMed]
- Suwanamornlert, P.; Sangchote, S.; Chinsirikul, W.; Sane, A.; Chonhenchob, V. Antifungal activity of plant-derived compounds and their synergism against major post-harvest pathogens of longan fruit in vitro. Int. J. Food Microbiol. 2018, 271, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Rosas-Gallo, A.; Ramírez-Corona, N.; Palou, E.; López-Malo, A. Modeling Penicillium expansum growth response to thyme essential oil at selected water activities and pH values using surface response methodology. Procedia Food Sci. 2016, 7, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Phoulivong, S.; Cai, L.; Chen, H.; McKenzie, E.H.C.; Abdelsalam, K.; Chukeatirote, E.; Hyde, K.D. Colletotrichum gloeosporioides is not a common pathogen on tropical fruits. Fungal Divers. 2010, 44, 33–43. [Google Scholar] [CrossRef]
- Wei, Y.; Pu, J.; Zhang, H.; Liu, Y.; Zhou, F.; Zhang, K.; Liu, X. The laccase gene (LAC1) is essential for Colletotrichum gloeosporioides development and virulence on mango leaves and fruits. Physiol. Mol. Plant Pathol. 2017, 1–10. [Google Scholar] [CrossRef]
- Ponzo, F.S.; Benato, E.A.; De-Silva, B.M.P.; Cia, P. Ethanol on the post-harvest control of anthracnose in ‘Kumagai’ guava. Bragantia 2018, 77, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, J.; Wan, C. Pinocembrin-7-Glucoside (P7G) reduced postharvest blue mold of navel orange by suppressing Penicillium italicum growth. Microorganisms 2020, 8, 536. [Google Scholar] [CrossRef] [Green Version]
- Koul, O. Biological activity of volatile di-n-propyl disulfide from seeds of neem, Azadirachta indica (Meliaceae), to two species of stored grain pests, Sitophilus oryzae (L.) and Tribolium castaneum (Herbst). J. Econ. Entomol. 2004, 97, 1142–1147. [Google Scholar] [CrossRef]
- Lyer, S.R.; Williamson, D. Efficacy of some plant extracts to inhibit the protease activity of Trichophyton spesies. Geobios 1991, 8, 3–6. [Google Scholar]
- Ramos De Resende, A.; Ludke Falcao, L.; Salviano Barbosa, G.; Helena Marcellino, L.; Silvano Gander, E. Neem (Azadirachta indica A. Juss) components: Candidates for control of Crinipellis perniciosa and Phytophthora spp. Microbiol. Res. 2007, 162, 238–243. [Google Scholar] [CrossRef]
- Zhong-hui, P.; Yu-qun, Z.; Zhong-qiong, Y.; Jiao, X.; Ren-yong, J.; Yang, L.; Fan, Y. Antibacterial activity of 9-octadecanoic acid-hexadecanoic acid-tetrahydrofuran-3, 4-diyl ester from neem oil. Agric. Sci. China 2010, 9, 1236–1240. [Google Scholar] [CrossRef]
- Okemo, P.O.; Mwatha, W.E.; Chhabra, S.C.; Fabry, W. The kill kinetics of Azadirachta indica A. juss. (Meliacae) extracts on Staphylococcus aureus, Escherichia coli, Pseudomonas aeruginosa and Candida albicans. Afr. J. Sci. Technol. 2001, 2, 113–118. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrations (g L−1) | C. acutatum (Day 14) | C. gloeosporioides (Day 10) |
|---|---|---|
| 800 | 29.23 aB (0.88) | 43.87 aA (1.70) |
| 400 | 23.41 bB (2.89) | 33.53 bA (1.75) |
| 200 | 19.34 cB (2.06) | 28.93 cA (1.93) |
| 100 | 17.96 cA (0.78) | 17.73 dA (0.55) |
| 50 | 11.74 dB (2.54) | 15.51 dA (1.70) |
| 25 | 5.26 eA (2.00) | 6.63 eA (2.39) |
| MIC * | ˃80 | ˃80 |
| Concentrations (g L−1) | C. acutatum (Day 14) | C. gloeosporioides (Day 10) |
|---|---|---|
| 800 | 35.19 aA (2.84) | 33.93 aA (1.10) |
| 400 | 25.15 bA (1.07) | 18.43 bB (2.13) |
| 200 | 19.93 cA (1.50) | 11.36 cB (2.18) |
| 100 | 9.33 dA (1.09) | 6.95 dB (1.73) |
| 50 | 8.64 dA (1.71) | 3.44 eB (1.15) |
| 25 | 7.15 dA (2.66) | 2.95 eB (0.82) |
| MIC * | ˃80 | ˃80 |
| Antifungal Agents | C. acutatum | C. gloeosporioides | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
(mm) | (1/d) | (d) | MSE | R2 | (mm) | (1/d) | (d) | MSE | R2 | |
| Distilled water | 2.79 a (0.01) | 0.81 a (0.01) | 0.96 b (0.01) | 0.091 | 0.905 | 2.86 a (0.01) | 0.94 a (0.00) | 0.91 b (0.00) | 0.041 | 0.949 |
| Ethanol | 2.64 b (0.03) | 0.66 c (0.01) | 0.99 b (0.00) | 0.077 | 0.911 | 2.82 a (0.02) | 0.87 b (0.02) | 0.94 b (0.01) | 0.044 | 0.938 |
| Propyl disulfide | 2.47 c (0.02) | 0.76 b (0.02) | 1.32 a (0.12) | 0.244 | 0.750 | 2.36 b (0.02) | 0.74 c (0.02) | 1.38 a (0.09) | 0.189 | 0.763 |
| Antifungal Agents | C. acutatum | C. gloeosporioides | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (mm) | (1/d) | (d) | MSE | R2 | (mm) | (1/d) | (d) | MSE | R2 | |
| Distilled water | 2.63 a (0.01) | 0.84 a (0.01) | 0.93 b (0.01) | 0.056 | 0.939 | 2.85 a (0.02) | 0.97 a (0.01) | 0.88 b (0.02) | 0.052 | 0.934 |
| Ethanol | 2.63 a (0.01) | 0.82 a (0.01) | 0.94 b (0.00) | 0.084 | 0.903 | 2.83 a (0.02) | 0.96 a (0.01) | 0.89 b (0.00) | 0.039 | 0.952 |
| Propyl disulfide | 2.40 b (0.01) | 0.69 b (0.01) | 1.53 a (0.17) | 0.064 | 0.924 | 2.49 b (0.01) | 0.72 b (0.01) | 1.33 a (0.00) | 0.225 | 0.722 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.R.; Chonhenchob, V.; Huang, C.; Suwanamornlert, P. Antifungal Activity of Propyl Disulfide from Neem (Azadirachta indica) in Vapor and Agar Diffusion Assays against Anthracnose Pathogens (Colletotrichum gloeosporioides and Colletotrichum acutatum) in Mango Fruit. Microorganisms 2021, 9, 839. https://doi.org/10.3390/microorganisms9040839
Khan MR, Chonhenchob V, Huang C, Suwanamornlert P. Antifungal Activity of Propyl Disulfide from Neem (Azadirachta indica) in Vapor and Agar Diffusion Assays against Anthracnose Pathogens (Colletotrichum gloeosporioides and Colletotrichum acutatum) in Mango Fruit. Microorganisms. 2021; 9(4):839. https://doi.org/10.3390/microorganisms9040839
Chicago/Turabian StyleKhan, Muhammad Rafiullah, Vanee Chonhenchob, Chongxing Huang, and Panitee Suwanamornlert. 2021. "Antifungal Activity of Propyl Disulfide from Neem (Azadirachta indica) in Vapor and Agar Diffusion Assays against Anthracnose Pathogens (Colletotrichum gloeosporioides and Colletotrichum acutatum) in Mango Fruit" Microorganisms 9, no. 4: 839. https://doi.org/10.3390/microorganisms9040839