
The Analysis of Live-Attenuated Piscirickettsia salmonis Vaccine Reveals the Short-Term Upregulation of Innate and Adaptive Immune Genes in Atlantic Salmon (Salmo salar): An In Situ Open-Sea Cages Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish
2.2. Vaccination Trial and Sampling
2.3. RNA Extraction and cDNA Synthesis
2.4. Gene Expression Analysis
2.5. Statistical Analysis
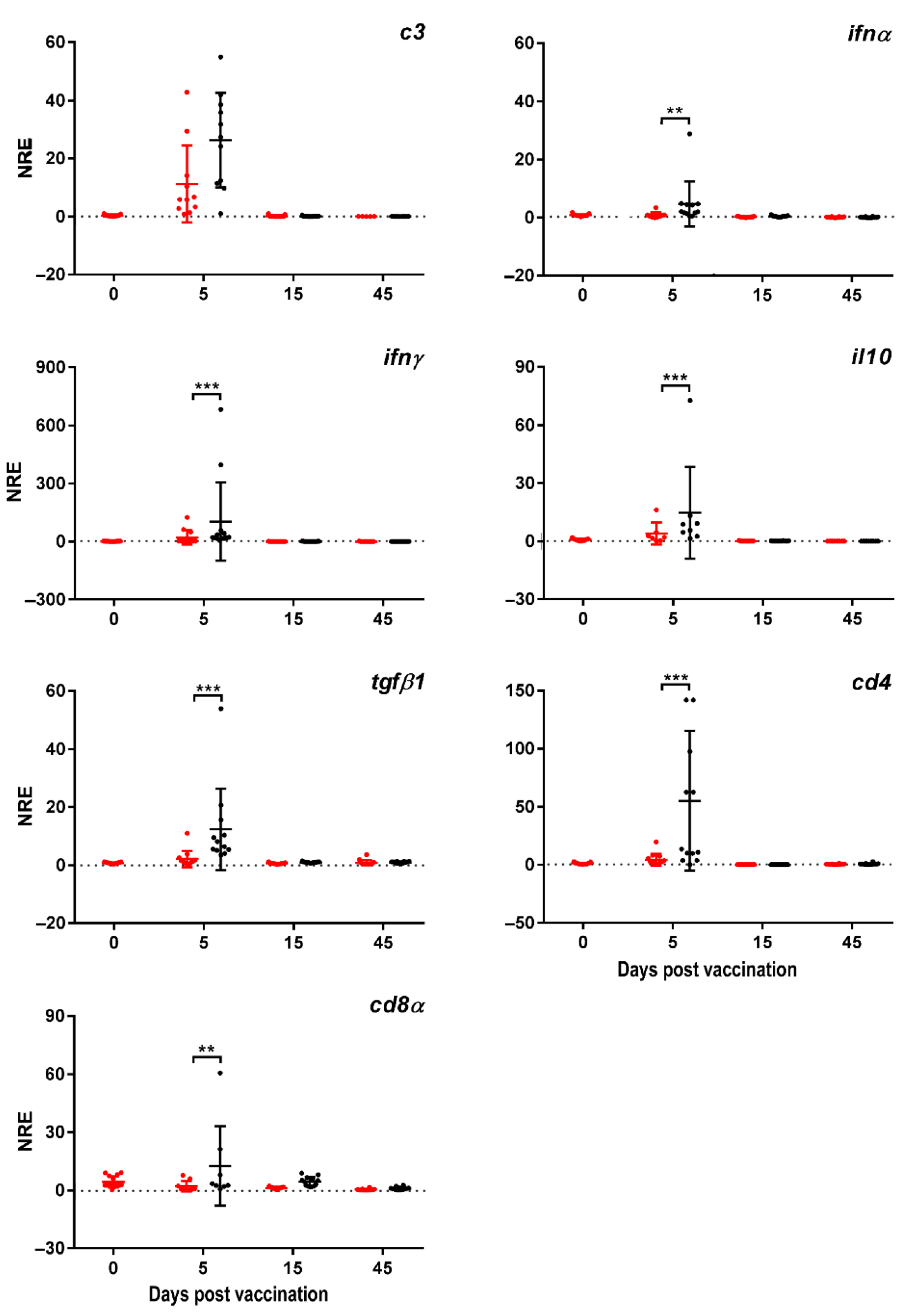
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Ethics Statement
References
- FAO. The State of World Fisheries and Aquaculture 2020; Sustainability in Action; FAO: Rome, Italy, 2020. [Google Scholar]
- FAO. Report of the FAO/MSU/WB First Multi-Stakeholder Consultation on a Progressive Management Pathway to Improve Aquaculture Biosecurity (PMP/AB). In Proceedings of the Stakeholder Consultation on PMP for Improving Aquaculture Biosecurity, Washington, DC, USA, 10–12 April 2018; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019; Volume 1254. [Google Scholar]
- Tort, L. Stress and immune modulation in fish. Dev. Comp. Immunol. 2011, 35, 1366–1375. [Google Scholar] [CrossRef]
- Figueroa, C.; Veloso, P.; Espin, L.; Dixon, B.; Torrealba, D.; Elalfy, I.S.; Afonso, J.M.; Soto, C.; Conejeros, P.; Gallardo, J.A. Host genetic variation explains reduced protection of commercial vaccines against Piscirickettsia salmonis in Atlantic salmon. Sci. Rep. 2020, 10, 18252. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Harvell, C.D.; Conrad, J.M.; Friedman, C.S.; Kent, M.L.; Kuris, A.M.; Powell, E.N.; Rondeau, D.; Saksida, S.M. Infectious diseases affect marine fisheries and aquaculture economics. Ann. Rev. Mar. Sci. 2015, 7, 471–496. [Google Scholar] [CrossRef] [Green Version]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Vet. Med. Int. 2018, 2018, 5432497. [Google Scholar] [CrossRef] [PubMed]
- Brudeseth, B.E.; Wiulsrod, R.; Fredriksen, B.N.; Lindmo, K.; Lokling, K.E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol. 2013, 35, 1759–1768. [Google Scholar] [CrossRef] [PubMed]
- Aqua Sernapesca: “El SRS Sigue Siendo el Mayor Problema Sanitario que Enfrenta la Salmoniculttura”. Available online: http://www.aqua.cl/2012/11/23/sernapesca-el-srs-sigue-siendo-el-mayor-problema-sanitario-que-enfrenta-la-salmoniculttura/# (accessed on 10 January 2021).
- SERNAPESCA. Informe Sanitario Acuícola año 2017; Departamento de Salud Animal, Subdirección de Acuicultura, Servicio Nacional de Pesca y Acuicultura: Valparaíso, Chile, 2018. [Google Scholar]
- SERNAPESCA. Informe Sanitario Acuícola año 2018; Departamento de Salud Animal, Subdirección de Acuicultura, Servicio Nacional de Pesca y Acuicultura: Valparaíso, Chile, 2019. [Google Scholar]
- SERNAPESCA. Informe Sanitario Acuícola año 2012; Unidad de Salud Animal, Subdirección de Acuicultura, Servicio Nacional de Pesca y Acuicultura, Servicio Nacional de Pesca y Acuicultura: Valparaíso, Chile, 2013. [Google Scholar]
- Marshall, S.H.; Conejeros, P.; Zahr, M.; Olivares, J.; Gomez, F.; Cataldo, P.; Henriquez, V. Immunological characterization of a bacterial protein isolated from salmonid fish naturally infected with Piscirickettsia salmonis. Vaccine 2007, 25, 2095–2102. [Google Scholar] [CrossRef]
- Maisey, K.; Montero, R.; Christodoulides, M. Vaccines for piscirickettsiosis (salmonid rickettsial septicaemia, SRS): The Chile perspective. Expert Rev. Vaccines 2017, 16, 215–228. [Google Scholar] [CrossRef]
- SERNAPESCA. Informe Sobre uso de Antimicrobianos en la Salmonicultura Nacional; Departamento de Salud Animal: Valparaíso, Chile, 2019. [Google Scholar]
- Flores-Kossack, C.; Montero, R.; Köllner, B.; Maisey, K. Chilean aquaculture and the new challenges: Pathogens, immune response, vaccination and fish diversification. Fish Shellfish Immunol. 2020, 98, 52–67. [Google Scholar] [CrossRef]
- Rozas, M.; Enriquez, R. Piscirickettsiosis and Piscirickettsia salmonis in fish: a review. J Fish Dis 2014, 37, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Evensen, O. Immunization Strategies against Piscirickettsia salmonis Infections: Review of Vaccination Approaches and Modalities and Their Associated Immune Response Profiles. Front. Immunol. 2016, 7, 482. [Google Scholar] [CrossRef] [Green Version]
- Sunyer, J.O. Fishing for mammalian paradigms in the teleost immune system. Nat. Immunol. 2013, 14, 320–326. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486–503. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Pellitero, P. Fish immunity and parasite infections: From innate immunity to immunoprophylactic prospects. Vet. Immunol. Immunopathol. 2008, 126, 171–198. [Google Scholar] [CrossRef] [PubMed]
- Tobar, I.; Arancibia, S.; Torres, C.; Vera, V.; Soto, P.; Carrasco, C.; Alvarado, M.; Neira, E.; Arcos, S.; Tobar, J.A. Successive Oral Immunizations Against Piscirickettsia Salmonis and Infectious Salmon Anemia Virus are Required to Maintain a Long-Term Protection in Farmed Salmonids. Front. Immunol. 2015, 6, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inohara, N.; Koseki, T.; Chen, S.; Wu, X.; Nunez, G. CIDE, a novel family of cell death activators with homology to the 45 kDa subunit of the DNA fragmentation factor. EMBO J. 1998, 17, 2526–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Stuardo, D.; Espinoza, A.; Tapia, S.; Morales-Reyes, J.; Barrientos, C.; Vallejos-Vidal, E.; Sandino, A.M.; Spencer, E.; Toro-Ascuy, D.; Rivas-Pardo, J.A.; et al. Non-Specific Antibodies Induce Lysosomal Activation in Atlantic Salmon Macrophages Infected by Piscirickettsia salmonis. Front. Immunol. 2020, 11, 544718. [Google Scholar] [CrossRef]
- Reyes-López, F.E.; Romeo, J.S.; Vallejos-Vidal, E.; Reyes-Cerpa, S.; Sandino, A.M.; Tort, L.; Mackenzie, S.; Imarai, M. Differential immune gene expression profiles in susceptible and resistant full-sibling families of Atlantic salmon (Salmo salar) challenged with infectious pancreatic necrosis virus (IPNV). Dev. Comp. Immunol. 2015, 53, 210–221. [Google Scholar] [CrossRef]
- Peña, A.A.; Bols, N.C.; Marshall, S.H. An evaluation of potential reference genes for stability of expression in two salmonid cell lines after infection with either Piscirickettsia salmonis or IPNV. BMC Res. Notes 2010, 3, 101. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ÄÄCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Bravo, S.; Midtlyng, P. The use of fish vaccines in the Chilean salmon industry 1999–2003. Aquaculture 2007, 270, 36–42. [Google Scholar] [CrossRef]
- Tobar, J.A.; Jerez, S.; Caruffo, M.; Bravo, C.; Contreras, F.; Bucarey, S.A.; Harel, M. Oral vaccination of Atlantic salmon (Salmo salar) against salmonid rickettsial septicaemia. Vaccine 2011, 29, 2336–2340. [Google Scholar] [CrossRef] [PubMed]
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current Status of the Use of Antibiotics and the Antimicrobial Resistance in the Chilean Salmon Farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef]
- Millanao, B.A.; Barrientos, H.M.; Gómez, C.C.; Tomova, A.; Buschmann, A.; Dölz, H.; Cabello, F.C. Injudicious and excessive use of antibiotics: Public health and salmon aquaculture in Chile. Rev. Med. Chile 2011, 139, 107–118. [Google Scholar]
- Shoemaker, C.A.; Klesius, P.H.; Evans, J.J.; Arias, C.R. Use of modified live vaccines in aquaculture. J. World Aquac. Soc. 2009, 40, 573–585. [Google Scholar] [CrossRef]
- Meza, K.; Inami, M.; Dalum, A.S.; Lund, H.; Bjelland, A.M.; Sorum, H.; Lovoll, M. Comparative evaluation of experimental challenge by intraperitoneal injection and cohabitation of Atlantic salmon (Salmo salar L.) after vaccination against Piscirickettsia salmonis (EM90-like). J. Fish Dis. 2019, 42, 1713–1730. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Stuardo, D.; Morales-Reyes, J.; Tapia, S.; Ahumada, D.E.; Espinoza, A.; Soto-Herrera, V.; Brianson, B.; Ibaceta, V.; Sandino, A.M.; Spencer, E.; et al. Non-lysosomal Activation in Macrophages of Atlantic Salmon (Salmo salar) After Infection With Piscirickettsia salmonis. Front. Immunol. 2019, 10, 434. [Google Scholar] [CrossRef]
- Beck, B.H.; Peatman, E. Mucosal Health in Aquaculture, 1st ed.; Academic Press: Oxford, UK, 2015; ISBN 9780124171862. [Google Scholar]
- Rozas-Serri, M.; Pena, A.; Arriagada, G.; Enriquez, R.; Maldonado, L. Comparison of gene expression in post-smolt Atlantic salmon challenged by LF-89-like and EM-90-like Piscirickettsia salmonis isolates reveals differences in the immune response associated with pathogenicity. J. Fish Dis. 2018, 41, 539–552. [Google Scholar] [CrossRef]
- Seder, R.A.; Hill, A. V Vaccines against intracellular infections requiring cellular immunity. Nature 2000, 406, 793–798. [Google Scholar] [CrossRef]
- Gan, Z.; Chen, S.N.; Huang, B.; Zou, J.; Nie, P. Fish type I and type II interferons: Composition, receptor usage, production and function. Rev. Aquac. 2020, 12, 773–804. [Google Scholar] [CrossRef] [Green Version]
- Green, D.S.; Young, H.A.; Valencia, J.C. Current prospects of type II interferon γ signaling & autoimmunity. J. Biol. Chem. 2017, 292, 13925–13933. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, C.A.; Gomez, F.A.; Mercado, L.; Ramírez, R.; Marshall, S.H. Piscirickettsia salmonis imbalances the innate immune response to succeed in a productive infection in a salmonid cell line model. PLoS ONE 2016, 11, e163943. [Google Scholar] [CrossRef] [Green Version]
- Constant, S.L.; Bottomly, K. Induction of Th1 and Th2 CD4+ T cell responses: The alternative approaches. Annu. Rev. Immunol. 1997, 15, 297–322. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Skjæveland, I.; Svingerud, T.; Zou, J.; Jørgensen, J.; Robertsen, B. Antiviral activity of salmonid gamma interferon against infectious pancreatic necrosis virus and salmonid alphavirus and its dependency on type I interferon. J. Virol. 2011, 85, 9188–9198. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Guo, T.-C.; Mutoloki, S.; Haugland, O.; Evensen, O. Gene expression studies of host response to Salmonid alphavirus subtype 3 experimental infections in Atlantic salmon. Vet. Res. 2012, 43, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertsen, B. The interferon system of teleost fish. Fish Shellfish Immunol. 2006, 20, 172–191. [Google Scholar] [CrossRef]
- Boehm, U.; Klamp, T.; Groot, M.; Howard, J.C. Cellular responses to interferon-gamma. Annu. Rev. Immunol. 1997, 15, 749–795. [Google Scholar] [CrossRef]
- Maisey, K.; Montero, R.; Corripio-Miyar, Y.; Toro-Ascuy, D.; Valenzuela, B.; Reyes-Cerpa, S.; Sandino, A.M.; Zou, J.; Wang, T.; Secombes, C.J.; et al. Isolation and Characterization of Salmonid CD4+ T Cells. J. Immunol. 2016, 196, 4150–4163. [Google Scholar] [CrossRef] [Green Version]
- Takizawa, F.; Magadan, S.; Parra, D.; Xu, Z.; Koryta, T.; Boudinot, P.; Sunyer, J.O. Novel Teleost CD4-Bearing Cell Populations Provide Insights into the Evolutionary Origins and Primordial Roles of CD4+ Lymphocytes and CD4+ Macrophages. J. Immunol. 2016, 196, 4522–4535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Bromage, E.S.; Kaattari, S.L. The strength of B cell interaction with antigen determines the degree of IgM polymerization. J. Immunol. 2010, 184, 844–850. [Google Scholar] [CrossRef] [Green Version]
- Kaattari, S.L.; Zhang, H.L.; Khor, I.W.; Kaattari, I.M.; Shapiro, D.A. Affinity maturation in trout: Clonal dominance of high affinity antibodies late in the immune response. Dev. Comp. Immunol. 2002, 26, 191–200. [Google Scholar] [CrossRef]
- Ye, J.; Kaattari, I.M.; Kaattari, S.L. The differential dynamics of antibody subpopulation expression during affinity maturation in a teleost. Fish Shellfish Immunol. 2011, 30, 372–377. [Google Scholar] [CrossRef]
- Wu, L.; Fu, S.; Yin, X.; Leng, W.; Guo, Z.; Wang, A.; Ye, J. Affinity maturation occurs in channel catfish (Ictalurus punctaus) following immunization with a T-cell dependent antigen. Fish Shellfish Immunol 2019, 84, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Fu, S.; Yin, X.; Guo, Z.; Wang, A.; Ye, J. Long-Lived Plasma Cells Secrete High-Affinity Antibodies Responding to a T-Dependent Immunization in a Teleost Fish. Front. Immunol. 2019, 10, 2324. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Takizawa, F.; Furihata, M.; Soto-Lampe, V.; Dijkstra, J.M.; Fischer, U. Teleost cytotoxic T cells. Fish Shellfish Immunol. 2019, 95, 422–439. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, M.; Araki, K.; Maruyoshi, K.; Matsumoto, M.; Nakayasu, C.; Moritomo, T.; Nakanishi, T.; Yamamoto, A. Comparative analysis of adaptive immune response after vaccine trials using live attenuated and formalin-killed cells of Edwardsiella tarda in ginbuna crucian carp (Carassius auratus langsdorfii). Fish Shellfish Immunol. 2015, 45, 437–442. [Google Scholar] [CrossRef]
- Nakanishi, T.; Shibasaki, Y.; Matsuura, Y. T Cells in Fish. Biology 2015, 4, 640–663. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Cerpa, S.; Maisey, K.; Reyes-Lpez, F.; Toro-Ascuy, D.; Mara, A.; Imarai, M. Fish Cytokines and Immune Response. In New Advances and Contributions to Fish Biology; InTech: London, UK, 2013; pp. 3–58. [Google Scholar]
- Li, F.; Zeng, B.; Chai, Y.; Cai, P.; Fan, C.; Cheng, T. The linker region of Smad2 mediates TGF- b -dependent ERK2-induced collagen synthesis. Biochem. Biophys. Res. Commun. 2009, 386, 289–293. [Google Scholar] [CrossRef]
- Reyes-Cerpa, S.; Reyes-López, F.; Toro-Ascuy, D.; Montero, R.; Maisey, K.; Acuña-Castillo, C.; Sunyer, J.O.; Parra, D.; Sandino, A.M.; Imarai, M. Induction of anti-inflammatory cytokine expression by IPNV in persistent infection. Fish Shellfish Immunol. 2014, 41, 172–182. [Google Scholar] [CrossRef]


{kind=link}
| Gene Acronym | Activation Step | Number of Cycles | Denaturation Step | Annealing Step | Extension Step | Tm Product (°C) |
|---|---|---|---|---|---|---|
| c3 | 95 °C × 2 min | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 85.6 |
| ifnα | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 83 | |
| ifnγ | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 81.8 | |
| il-10 | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 82.5 | |
| tgf-β | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 80.1 | |
| eIFα | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 85.5 | |
| 18S | 40 | 95 °C × 5 s | 60 °C × 10 s | 72 °C × 10 s | 82.7 | |
| cd4 | 95 °C × 2 min | 40 | 95 °C × 5 s | 57 °C × 10 s | 72 °C × 10 s | 83.8 |
| cd8a | 95 °C × 2 min | 40 | 95 °C × 5 s | 55 °C × 10 s | 72 °C × 10 s | 81.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas, D.; Vallejos-Vidal, E.; Reyes-Cerpa, S.; Oyarzún-Arrau, A.; Acuña-Castillo, C.; Imarai, M.; Reyes-López, F.E.; Sandino, A.M. The Analysis of Live-Attenuated Piscirickettsia salmonis Vaccine Reveals the Short-Term Upregulation of Innate and Adaptive Immune Genes in Atlantic Salmon (Salmo salar): An In Situ Open-Sea Cages Study. Microorganisms 2021, 9, 703. https://doi.org/10.3390/microorganisms9040703
Vargas D, Vallejos-Vidal E, Reyes-Cerpa S, Oyarzún-Arrau A, Acuña-Castillo C, Imarai M, Reyes-López FE, Sandino AM. The Analysis of Live-Attenuated Piscirickettsia salmonis Vaccine Reveals the Short-Term Upregulation of Innate and Adaptive Immune Genes in Atlantic Salmon (Salmo salar): An In Situ Open-Sea Cages Study. Microorganisms. 2021; 9(4):703. https://doi.org/10.3390/microorganisms9040703
Chicago/Turabian StyleVargas, Deborah, Eva Vallejos-Vidal, Sebastián Reyes-Cerpa, Aarón Oyarzún-Arrau, Claudio Acuña-Castillo, Mónica Imarai, Felipe E. Reyes-López, and Ana María Sandino. 2021. "The Analysis of Live-Attenuated Piscirickettsia salmonis Vaccine Reveals the Short-Term Upregulation of Innate and Adaptive Immune Genes in Atlantic Salmon (Salmo salar): An In Situ Open-Sea Cages Study" Microorganisms 9, no. 4: 703. https://doi.org/10.3390/microorganisms9040703