
Modulation of IGF2 Expression in the Murine Thymus and Thymic Epithelial Cells Following Coxsackievirus-B4 Infection

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Mice Inoculation with CV-B4
2.3. TEC Isolation, Enrichment, and Immunostaining
2.4. Infection of MTE4-14 Cells with CV-B4
2.5. Modified TCID50 Titration Assay
2.6. Flow Cytometry
2.7. One-Step and Two-Step CV-B4 RNA Detection Using PCR
2.8. Reverse Transcription, Endpoint, and Real-Time Quantitative PCR
2.9. Igf2 P3 Nluc Plasmid Construction and Site-Directed Mutagenesis
2.10. Dual-Luciferase Reporter Assay
2.11. mRNA Stability Assay
2.12. SDS-PAGE and Western Blot
2.13. Statistical Analyses
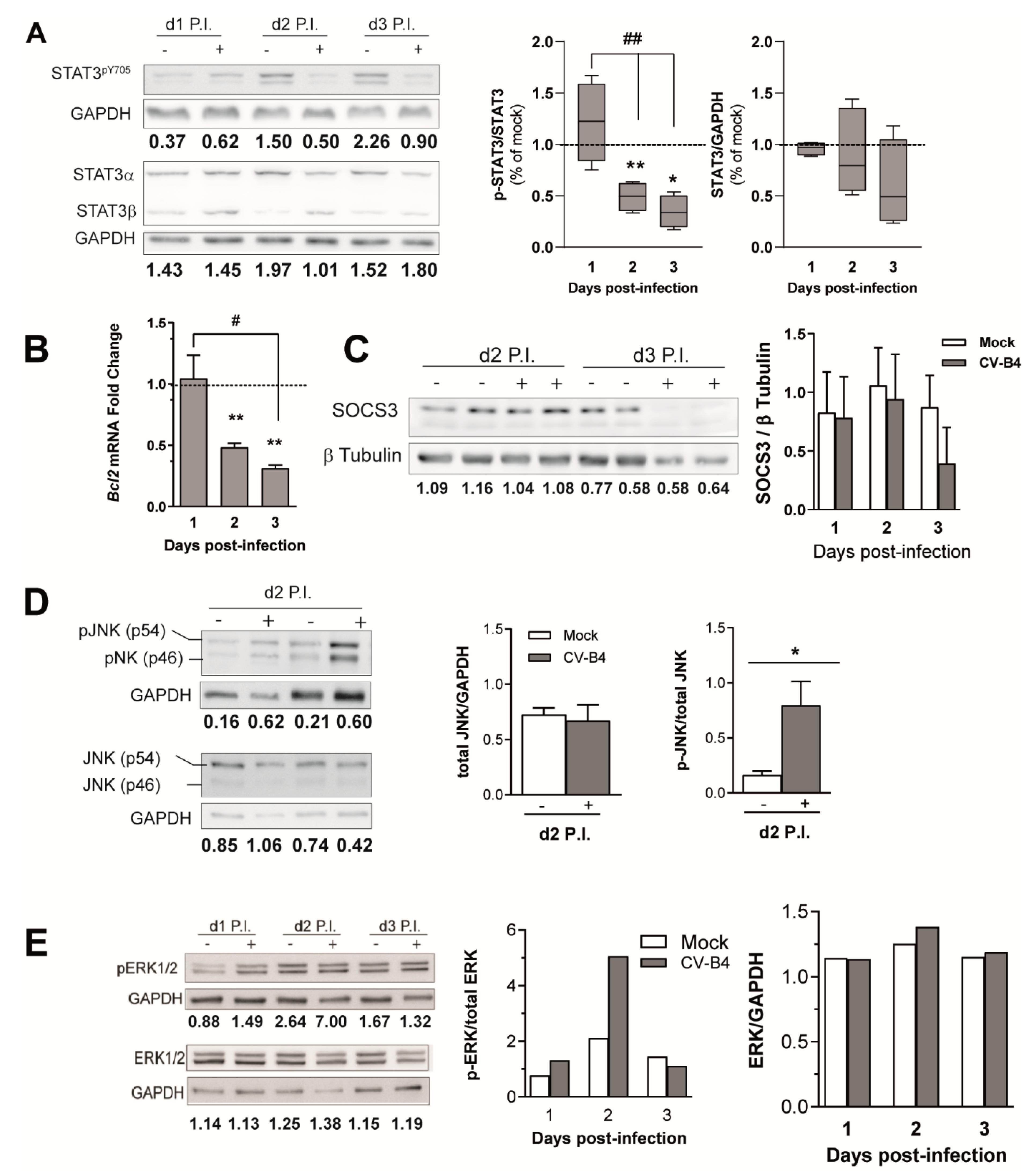
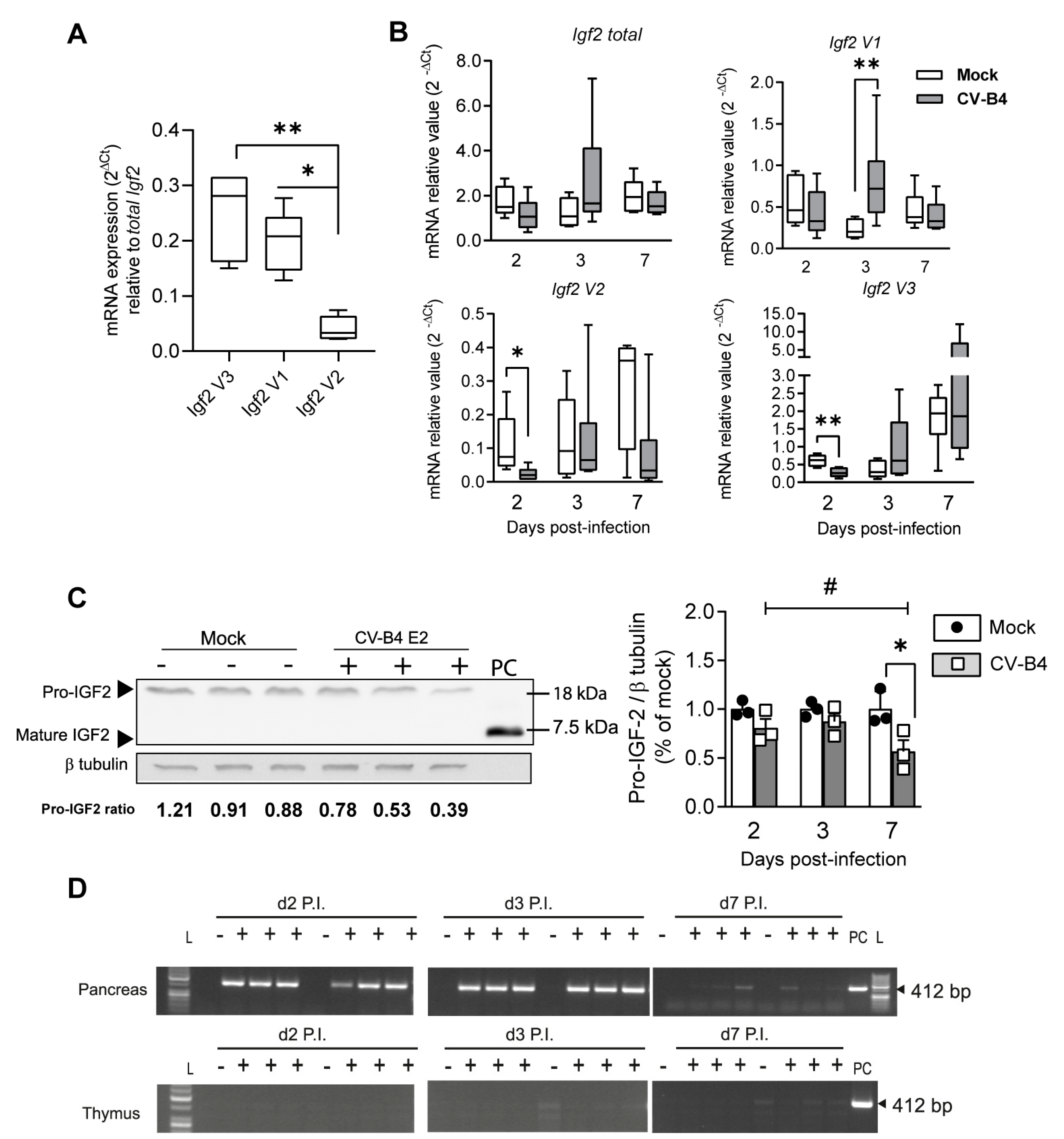
3. Results
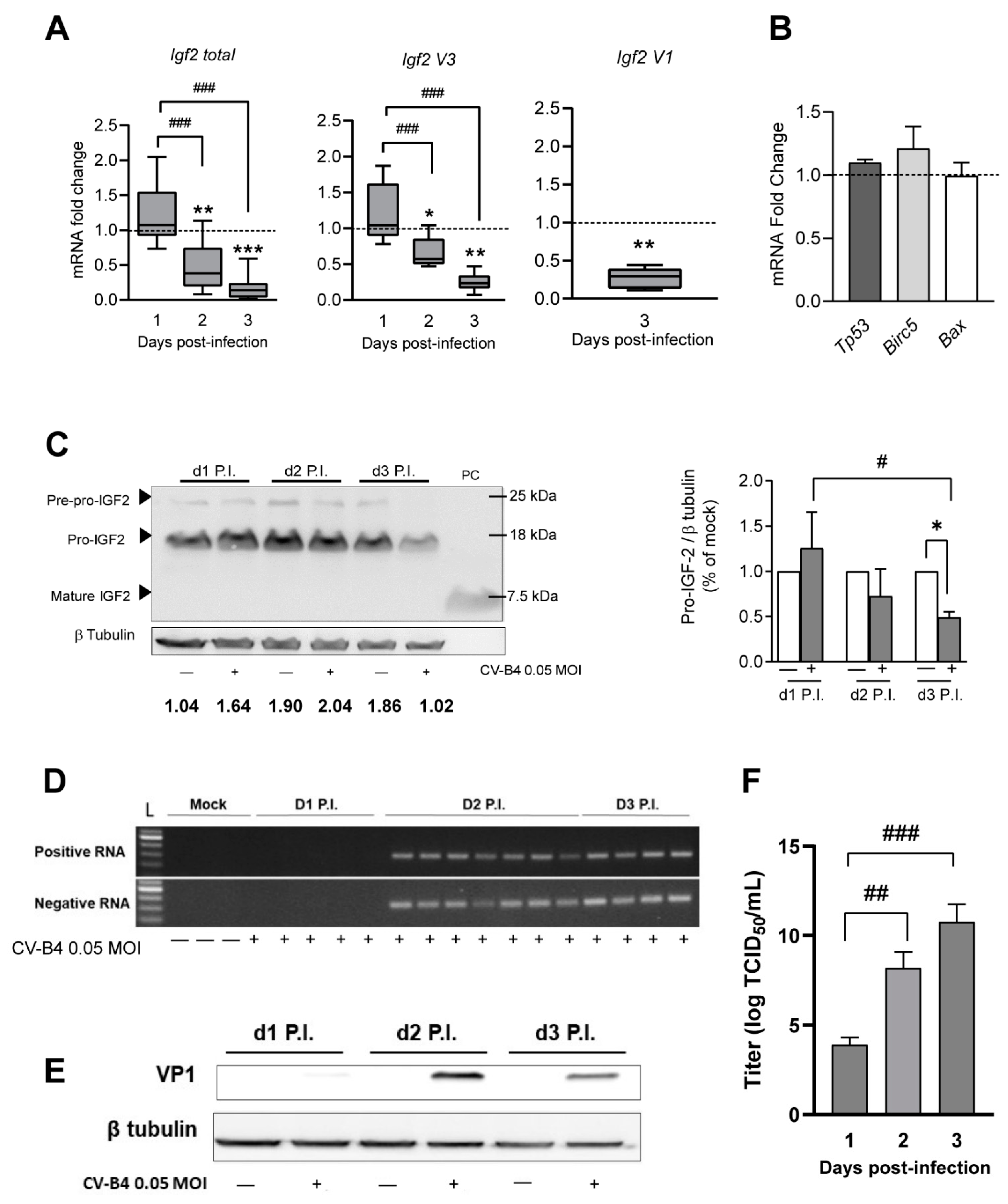
3.1. Decrease in Igf2 Transcripts and Pro-IGF2 Expression Following CV-B4 Infection of MTE4-14 Cells
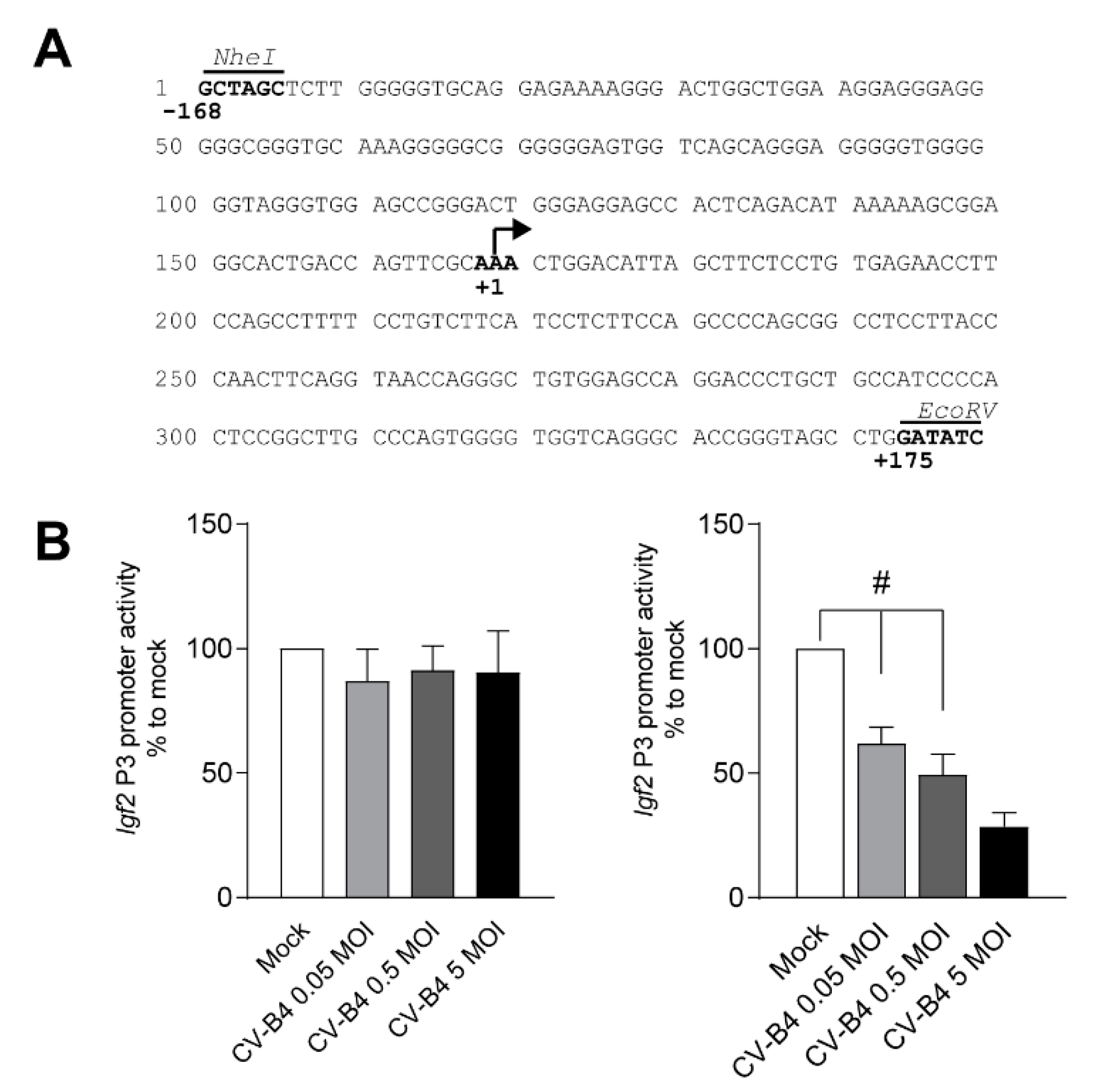
3.2. CV-B4 Infection Decreases Igf2 P3 Promoter Activity
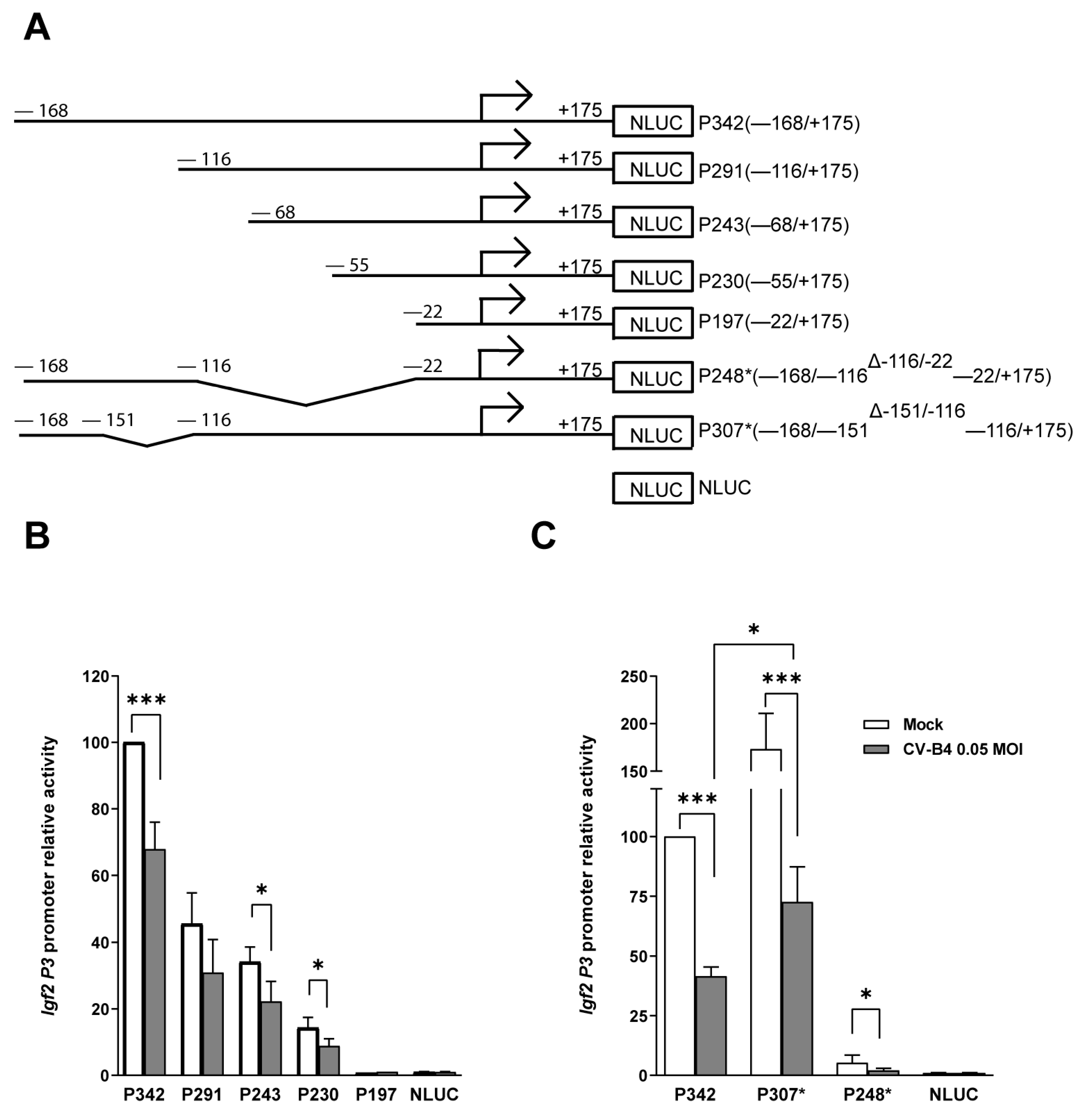
3.3. Impact of CV-B4 Infection on Specific Regions of the Igf2 P3 Promoter
3.4. CV-B4 Infection Decreases STAT3 Phosphorylation
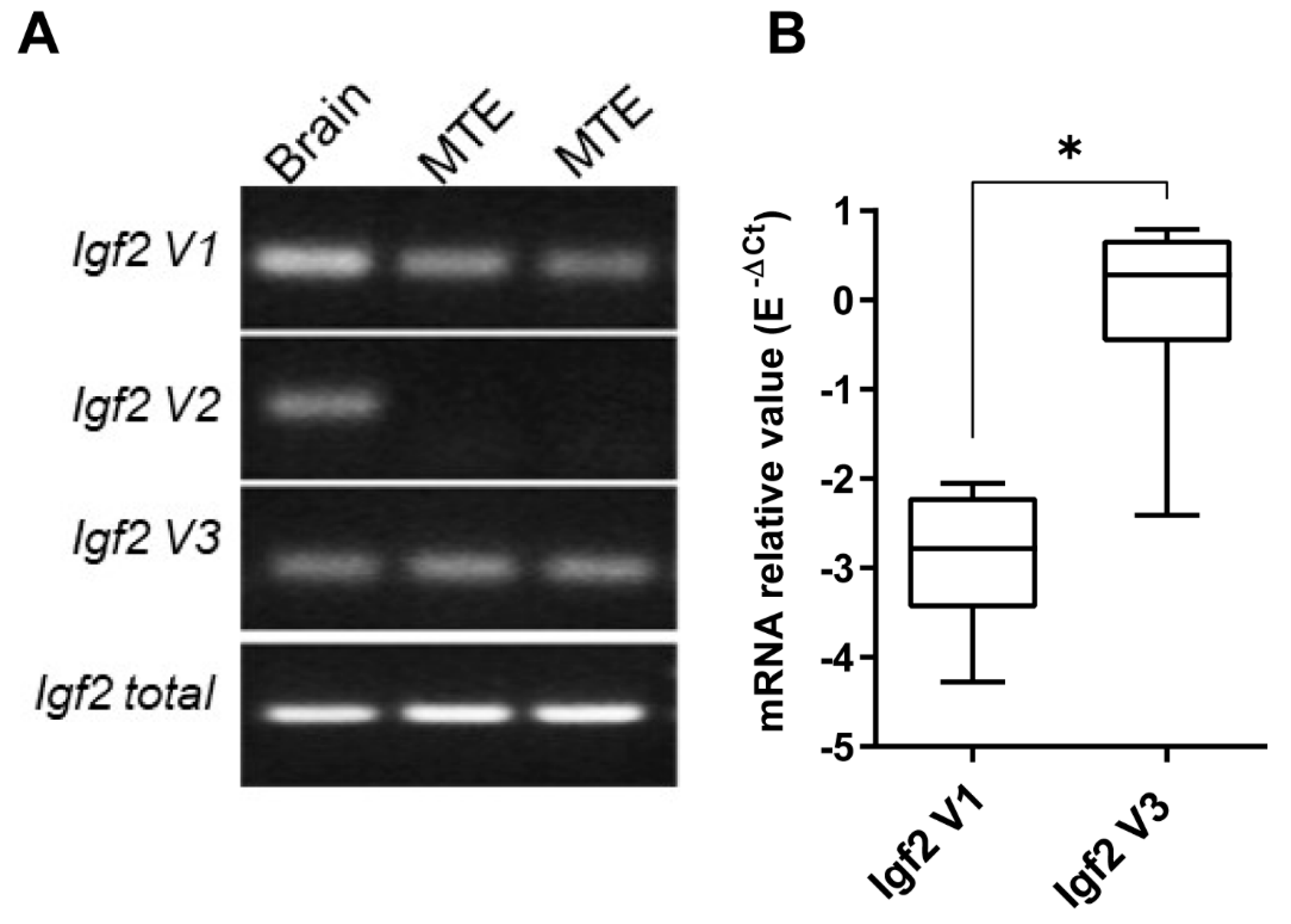
3.5. Decrease in Igf2 Transcripts and Downregulation of Pro-IGF2 Expression in TECs Following CV-B4 Inoculation In Vivo
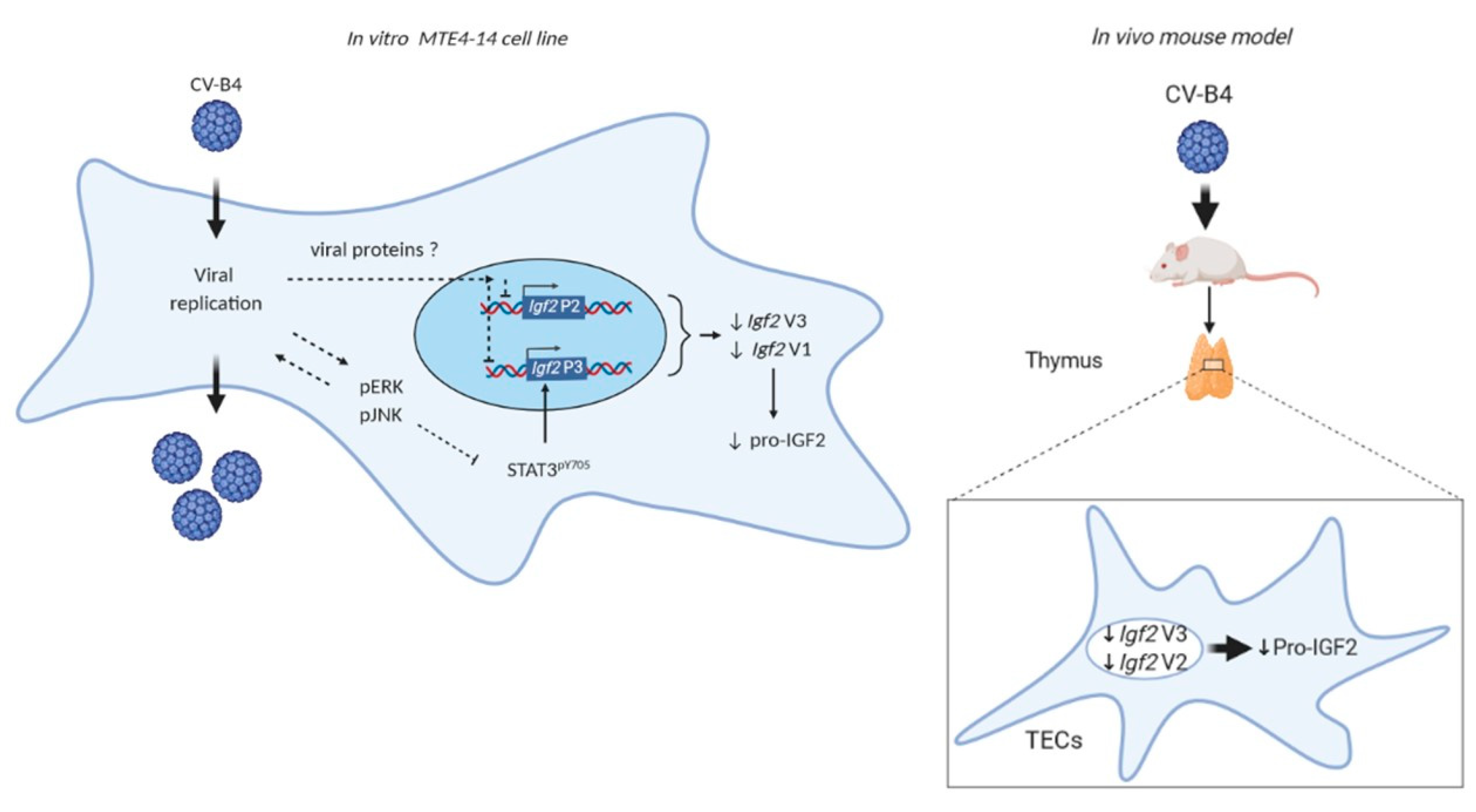
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geenen, V.; Trussart, C.; Michaux, H.; Halouani, A.; Jaïdane, H.; Collée, C.; Renard, C.; Daukandt, M.; Ledent, P.; Martens, H. The presentation of neuroendocrine self-peptides in the thymus: An essential event for individual life and vertebrate survival. Ann. N. Y. Acad. Sci. 2019, 1455, 113–125. [Google Scholar] [CrossRef]
- Geenen, V.; Louis, C.; Martens, H.; Registry, T.B.D. An Insulin-like Growth Factor 2-Derived Self-Antigen Inducing a Regulatory Cytokine Profile after Presentation to Peripheral Blood Mononuclear Cells from DQ8+Type 1 Diabetic Adolescents: Preliminary Design of a Thymus-Based Tolerogenic Self-Vaccination. Ann. N. Y. Acad. Sci. 2004, 1037, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Hansenne, I.; Renard-Charlet, C.; Greimers, R.; Geenen, V. Dendritic Cell Differentiation and Immune Tolerance to Insulin-Related Peptides in Igf2-Deficient Mice. J. Immunol. 2006, 176, 4651–4657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, M.R.; Wasserfall, C.H.; McGrail, S.M.; Posgai, A.L.; Bacher, R.; Muir, A.; Haller, M.J.; Schatz, D.A.; Wesley, J.D.; Von Herrath, M.; et al. Insulin-Like Growth Factor Dysregulation Both Preceding and Following Type 1 Diabetes Diagnosis. Diabetes 2020, 69, 413–423. [Google Scholar] [CrossRef]
- Yang, G.; Geng, X.-R.; Song, J.-P.; Wu, Y.; Yan, H.; Zhan, Z.; Yang, L.; He, W.; Liu, Z.-Q.; Qiu, S.; et al. Insulin-like growth factor 2 enhances regulatory T-cell functions and suppresses food allergy in an experimental model. J. Allergy Clin. Immunol. 2014, 133, 1702–1708.e5. [Google Scholar] [CrossRef] [PubMed]
- Geng, X.-R.; Yang, G.; Li, M.; Song, J.-P.; Liu, Z.-Q.; Qiu, S.; Liu, Z.; Yang, P.-C. Insulin-like Growth Factor-2 Enhances Functions of Antigen (Ag)-specific Regulatory B Cells. J. Biol. Chem. 2014, 289, 17941–17950. [Google Scholar] [CrossRef] [Green Version]
- Kecha-Kamoun, O.; Achour, I.; Martens, H. Thymic expression of insulin-related genes in an animal model of autoimmune type 1 diabetes. Diabetes Metab. Res. Rev. 2001, 17, 146–152. [Google Scholar] [CrossRef]
- Hober, D.; Sauter, P. Pathogenesis of type 1 diabetes mellitus: Interplay between enterovirus and host. Nat. Rev. Endocrinol. 2010, 6, 279–289. [Google Scholar] [CrossRef]
- Yoon, J.W.; Austin, M.; Onodera, T.; Notkins, A.L. Virus-Induced Diabetes Mellitus: Isolation of a Virus from the Pancreas of a Child with Diabetic Ketoacidosis. N. Engl. J. Med. 1979, 300, 1173–1179. [Google Scholar] [CrossRef]
- Vargová, A.; Bopegamage, S.; Borsanyiová, M.; Petrovičová, A.; Benkovičová, M. Coxsackievirus infection of mice. II. Viral kinetics and histopathological changes in mice experimentally infected with Coxsackievirus B3 by intraperitoneal route. Acta Virol. 2003, 47, 253–257. [Google Scholar]
- Bopegamage, S.; Kovacova, J.; Vargova, A.; Motusova, J.; Petrovicova, A.; Benkovicova, M.; Gomolcak, P.; Bakkers, J.; Van Kuppeveld, F.; Melchers, W.J.G.; et al. Coxsackie B virus infection of mice: Inoculation by the oral route protects the pancreas from damage, but not from infection. J. Gen. Virol. 2005, 86, 3271–3280. [Google Scholar] [CrossRef]
- Chatterjee, N.K.; Hou, J.; Dockstader, P.; Charbonneau, T. Coxsackievirus B4 infection alters thymic, splenic, and peripheral lymphocyte repertoire preceding onset of hyperglycemia in mice. J. Med Virol. 1992, 38, 124–131. [Google Scholar] [CrossRef]
- Jaïdane, H.; Gharbi, J.; Lobert, P.-E.; Lucas, B.; Hiar, R.; Ben M’Hadheb, M.; Brilot, F.; Geenen, V.; Aouni, M.; Hober, D. Prolonged Viral RNA Detection in Blood and Lymphoid Tissues fromCoxsackievirus B4 E2Orally-InoculatedSwissMice. Microbiol. Immunol. 2006, 50, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Brilot, F.; Geenen, V.; Hober, D.; Stoddart, C.A. Coxsackievirus B4 Infection of Human Fetal Thymus Cells. J. Virol. 2004, 78, 9854–9861. [Google Scholar] [CrossRef] [Green Version]
- Brilot, F.; Jaïdane, H.; Geenen, V.; Hober, D. Coxsackievirus B4 infection of murine foetal thymus organ cultures. J. Med. Virol. 2008, 80, 659–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brilot, F.; Chehadeh, W.; Charlet-Renard, C.; Martens, H.; Geenen, V.; Hober, D. Persistent Infection of Human Thymic Epithelial Cells by Coxsackievirus B4. J. Virol. 2002, 76, 5260–5265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepesant, H.; Pierres, M.; Naquet, P. Deficient Antigen Presentation by Thymic Epithelial Cells Reveals Differential Induction of T Cell Clone Effector Functions by CD28-Mediated Costimulation. Cell. Immunol. 1995, 161, 279–287. [Google Scholar] [CrossRef]
- Jaïdane, H.; Caloone, D.; Lobert, P.-E.; Sane, F.; Dardenne, O.; Naquet, P.; Gharbi, J.; Aouni, M.; Geenen, V.; Hober, D. Persistent Infection of Thymic Epithelial Cells with Coxsackievirus B4 Results in Decreased Expression of Type 2 Insulin-Like Growth Factor. J. Virol. 2012, 86, 11151–11162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergman, D.; Halje, M.; Nordin, M.; Engström, W. Insulin-Like Growth Factor 2 in Development and Disease: A Mini-Review. Gerontology 2013, 59, 240–249. [Google Scholar] [CrossRef]
- Panda, A.C.; Grammatikakis, I.; Yoon, J.-H.; Abdelmohsen, K. Posttranscriptional Regulation of Insulin Family Ligands and Receptors. Int. J. Mol. Sci. 2013, 14, 19202–19229. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Roth, A.; Yu, M.; Morris, R.; Bersani, F.; Rivera, M.N.; Lu, J.; Shioda, T.; Vasudevan, S.; Ramaswamy, S.; et al. The IGF2 intronic miR-483 selectively enhances transcription from IGF2 fetal promoters and enhances tumorigenesis. Genes Dev. 2013, 27, 2543–2548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, J.C.; Baron, J. Evidence that Igf2 down-regulation in postnatal tissues and up-regulation in malignancies is driven by transcription factor E2f3. Proc. Natl. Acad. Sci. USA 2013, 110, 6181–6186. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Wang, C.; Xiao, F.; Wang, H.; Wu, Z. JAK2/STAT2/STAT3 are required for myogenic differentiation. J. Biol. Chem. 2008, 283, 34029–34036. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-C.; Min, H.-Y.; Jung, H.J.; Park, K.H.; Hyun, S.Y.; Cho, J.; Woo, J.K.; Kwon, S.J.; Lee, H.-J.; Johnson, F.M. Essential role of insulin-like growth factor 2 in resistance to histone deacetylase inhibitors. Oncogene 2016, 35, 5515–5526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himes, B.E.; Obraztsova, K.; Lian, L.; Shumyatcher, M.; Rue, R.; Atochina-Vasserman, E.N.; Hur, S.K.; Bartolomei, M.S.; Evans, J.F.; Krymskaya, V.P. Rapamycin-independent IGF2 expression in Tsc2-null mouse embryo fibroblasts and human lymphangioleiomyomatosis cells. PLoS ONE 2018, 13, e0197105. [Google Scholar] [CrossRef]
- Satoh, R.; Kakugawa, K.; Yasuda, T.; Yoshìda, H.; Sibilia, M.; Katsura, Y.; Levi, B.; Abramson, J.; Koseki, Y.; Koseki, H.; et al. Requirement of Stat3 Signaling in the Postnatal Development of Thymic Medullary Epithelial Cells. PLoS Genet. 2016, 12, e1005776. [Google Scholar] [CrossRef] [Green Version]
- Lomada, D.; Jain, M.; Bolner, M.; Reeh, K.A.G.; Kang, R.; Reddy, M.C.; DiGiovanni, J.; Richie, E.R. Stat3 Signaling Promotes Survival And Maintenance Of Medullary Thymic Epithelial Cells. PLoS Genet. 2016, 12, e1005777. [Google Scholar] [CrossRef] [Green Version]
- Sano, S.; Takahama, Y.; Sugawara, T.; Kosaka, H.; Itami, S.; Yoshikawa, K.; Miyazaki, J.-I.; Van Ewijk, W.; Takeda, J. Stat3 in Thymic Epithelial Cells Is Essential for Postnatal Maintenance of Thymic Architecture and Thymocyte Survival. Immunity 2001, 15, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Wang, H.-W.; Cuenca, A.; Huang, M.; Ghansah, T.; Brayer, J.; Kerr, W.G.; Takeda, K.; Akira, S.; Schoenberger, S.P.; et al. A Critical Role for Stat3 Signaling in Immune Tolerance. Immunity 2003, 19, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.; Hogquist, K.A. Isolation, identification, and purification of murine thymic epithelial cells. J. Vis. Exp. 2014, 8, e51780. [Google Scholar] [CrossRef] [Green Version]
- Roberts, N.A.; Adams, B.D.; McCarthy, N.I.; Tooze, R.M.; Parnell, S.M.; Anderson, G.; Kaech, S.M.; Horsley, V. Prdm1 Regulates Thymic Epithelial Function To Prevent Autoimmunity. J. Immunol. 2017, 199, 1250–1260. [Google Scholar] [CrossRef] [PubMed]
- Pourianfar, H.R.; Javadi, A.; Grollo, L. A Colorimetric-Based Accurate Method for the Determination of Enterovirus 71 Titer. Indian J. Virol. 2012, 23, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotwein, P.; Hall, L.J. Evolution of Insulin-Like Growth Factor II: Characterization of the Mouse IGF-II Gene and Identification of Two Pseudo-Exons. DNA Cell Biol. 1990, 9, 725–735. [Google Scholar] [CrossRef]
- Caricasole, A.; Ward, A. Transactivation of mouse insulin-like growth factor II (IGF-II) gene promoters by the AP-1 complex. Nucleic Acids Res. 1993, 21, 1873–1879. [Google Scholar] [CrossRef] [Green Version]
- Dreos, R.; Ambrosini, G.; Périer, R.C.; Bucher, P. EPD and EPDnew, high-quality promoter resources in the next-generation sequencing era. Nucleic Acids Res. 2012, 41, D157–D164. [Google Scholar] [CrossRef]
- Bok, K.; Prikhodko, V.G.; Green, K.Y.; Sosnovtsev, S.V. Apoptosis in Murine Norovirus-Infected RAW264.7 Cells Is Associated with Downregulation of Survivin. J. Virol. 2009, 83, 3647–3656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chau, D.H.W.; Yuan, J.; Zhang, H.; Cheung, P.; Lim, T.; Liu, Z.; Sall, A.; Yang, D. Coxsackievirus B3 proteases 2A and 3C induce apoptotic cell death through mitochondrial injury and cleavage of eIF4GI but not DAP5/p97/NAT1. Apoptosis 2007, 12, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, S.; Chen, Y.; Wang, T.; Dong, C.; Wo, X.; Zhang, J.; Dong, Y.; Xu, W.; Feng, X.; et al. The Capsid Protein VP1 of Coxsackievirus B Induces Cell Cycle Arrest by Up-Regulating Heat Shock Protein 70. Front. Microbiol. 2019, 10, 1633. [Google Scholar] [CrossRef] [Green Version]
- Durant, L.; Watford, W.T.; Ramos, H.L.; Laurence, A.; Vahedi, G.; Wei, L.; Takahashi, H.; Sun, H.-W.; Kanno, Y.; Powrie, F.; et al. Diverse Targets of the Transcription Factor STAT3 Contribute to T Cell Pathogenicity and Homeostasis. Immunity 2010, 32, 605–615. [Google Scholar] [CrossRef] [Green Version]
- Weirauch, M.T.; Yang, A.; Albu, M.; Cote, A.G.; Montenegro-Montero, A.; Drewe, P.; Najafabadi, H.S.; Lambert, S.A.; Mann, I.; Cook, K.; et al. Determination and Inference of Eukaryotic Transcription Factor Sequence Specificity. Cell 2014, 158, 1431–1443. [Google Scholar] [CrossRef] [Green Version]
- Kiuchi, N.; Nakajima, K.; Ichiba, M.; Fukada, T.; Narimatsu, M.; Mizuno, K.; Hibi, M.; Hirano, T. STAT3 Is Required for the gp130-mediated Full Activation of the c-myc Gene. J. Exp. Med. 1999, 189, 63–73. [Google Scholar] [CrossRef]
- Wakahara, R.; Kunimoto, H.; Tanino, K.; Kojima, H.; Inoue, A.; Shintaku, H.; Nakajima, K. Phospho-Ser727 of STAT3 regulates STAT3 activity by enhancing dephosphorylation of phospho-Tyr705 largely through TC45. Genes Cells 2012, 17, 132–145. [Google Scholar] [CrossRef]
- Gkouveris, I.; Nikitakis, N.; Karanikou, M.; Rassidakis, G.; Sklavounou, A. JNK1/2 expression and modulation of STAT3 signaling in oral cancer. Oncol. Lett. 2016, 12, 699–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkouveris, I.; Nikitakis, N.; Karanikou, M.; Rassidakis, G.; Sklavounou, A. Erk1/2 activation and modulation of STAT3 signaling in oral cancer. Oncol. Rep. 2014, 32, 2175–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asfari, M.; De, W.; Nöel, M.; Holthuizen, P.E.; Czernichow, P. Insulin-like growth factor-II gene expression in a rat insulin-producing beta-cell line (INS-1) is regulated by glucose. Diabetologia 1995, 38, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Erbay, E.; Park, I.-H.; Nuzzi, P.D.; Schoenherr, C.J.; Chen, J. IGF-II transcription in skeletal myogenesis is controlled by mTOR and nutrients. J. Cell Biol. 2003, 163, 931–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaleb, A.M.; Yang, V.W. Krüppel-like factor 4 (KLF4): What we currently know. Gene 2017, 611, 27–37. [Google Scholar] [CrossRef]
- Lee, Y.I.; Kim, S.-J.; Kim, Y.I.L.-J. Transcriptional Repression of Human Insulin-Like Growth Factor-II P4 Promoter by Wilms’ Tumor Suppressor WT1. DNA Cell Biol. 1996, 15, 99–104. [Google Scholar] [CrossRef]
- Sun, D.; Chen, S.; Cheng, A.; Wang, M. Roles of the Picornaviral 3C Proteinase in the Viral Life Cycle and Host Cells. Viruses 2016, 8, 82. [Google Scholar] [CrossRef] [Green Version]
- Dimova, D.K.; Dyson, N.J. The E2F transcriptional network: Old acquaintances with new faces. Oncogene 2005, 24, 2810–2826. [Google Scholar] [CrossRef] [Green Version]
- Babon, J.J.; Varghese, L.N.; Nicola, N.A. Inhibition of IL-6 family cytokines by SOCS3. Semin. Immunol. 2014, 26, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, M.; Duan, H.; Gao, M.; Zhang, H.; Peng, Y. Both ERK1 and ERK2 Are Required for Enterovirus 71 (EV71) Efficient Replication. Viruses 2015, 7, 1344–1356. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Yanagawa, B.; Zhang, J.; Luo, Z.; Zhang, M.; Esfandiarei, M.; Carthy, C.; Wilson, J.E.; Yang, D.; McManus, B.M. Coxsackievirus B3 Replication Is Reduced by Inhibition of the Extracellular Signal-Regulated Kinase (ERK) Signaling Pathway. J. Virol. 2002, 76, 3365–3373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, X.; Luo, H.; Morgan, A.; Zhang, J.; Wong, J.; Yuan, J.; Esfandiarei, M.; Gao, G.; Cheung, C.; McManus, B.M. Stress-Activated Protein Kinases Are Involved in Coxsackievirus B3 Viral Progeny Release. J. Virol. 2005, 79, 13875–13881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguech-Oueslati, L.; Jaidane, H.; Sane, F.; Jrad-Battikh, N.; Ben Hamed, S.; Hober, D.; Gharbi, J. Evaluation of Contamination Risks with Coxsackievirus B4 E2 in Swiss Albino Mice Stools. Curr. Microbiol. 2017, 75, 32–39. [Google Scholar] [CrossRef]
- Kecha, O.; Martens, H.; Franchimont, N.; Achour, I.; Hazee-Hagelstein, M.-T.; Charlet-Renard, C.; Geenen, V.; Winkler, R. Characterization of the Insulin-Like Growth Factor Axis in the Human Thymus. J. Neuroendocr. 1999, 11, 435–440. [Google Scholar] [CrossRef]
- Kecha, O. Involvement of Insulin-Like Growth Factors in Early T Cell Development: A Study Using Fetal Thymic Organ Cultures. Endocrinology 2000. [Google Scholar] [CrossRef]
- Steinmetz, A.B.; Johnson, S.A.; Iannitelli, D.E.; Pollonini, G.; Alberini, C.M. Insulin-like growth factor 2 rescues aging-related memory loss in rats. Neurobiol. Aging 2016, 44, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.; Basak, A.; Mbikay, M.; Tsang, B.K.; Gruslin, A. Role of pro-IGF-II processing by proprotein convertase 4 in human placental development. Proc. Natl. Acad. Sci. USA 2005, 102, 11047–11052. [Google Scholar] [CrossRef] [Green Version]
- Van Koetsveld, P.M.; Vitale, G.; Feelders, R.A.; Waaijers, M.; Sprij-Mooij, D.M.; De Krijger, R.R.; Hofland, L.J.; de Herder, W.W.; Lamberts, S.W.J.; de Krijger, R.R.; et al. Interferon-β is a potent inhibitor of cell growth and cortisol production in vitro and sensitizes human adrenocortical carcinoma cells to mitotane. Endocr. Relat. Cancer 2013, 20, 443–454. [Google Scholar] [CrossRef] [Green Version]
- Booy, S.; van Eijck, C.H.J.; Janssen, J.A.M.J.L.; Dogan, F.; van Koetsveld, P.M.; Hofland, L.J. IFN-β is a potent inhibitor of insulin and insulin like growth factor stimulated proliferation and migration in human pancreatic cancer cells. Am. J. Cancer Res. 2015, 5, 2035. [Google Scholar]
- Henke, A.; Mohr, C.; Sprenger, H.; Graebner, C.; Stelzner, A.; Nain, M.; Gemsa, D. Coxsackievirus B3-induced production of tumor necrosis factor-alpha, IL-1 beta, and IL-6 in human monocytes. J. Immunol. 1992, 148, 2270–2277. [Google Scholar] [PubMed]
- Yang, K.; Puel, A.; Zhang, S.; Eidenschenk, C.; Ku, C.-L.; Casrouge, A.; Picard, C.; Von Bernuth, H.; Senechal, B.; Plancoulaine, S.; et al. Human TLR-7-, -8-, and -9-Mediated Induction of IFN-α/β and -λ Is IRAK-4 Dependent and Redundant for Protective Immunity to Viruses. Immunity 2005, 23, 465–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hober, D.; Chehadeh, W.; Bouzidi, A.; Wattré, P. Antibody-Dependent Enhancement of Coxsackievirus B4 Infectivity of Human Peripheral Blood Mononuclear Cells Results in Increased Interferon-α Synthesis. J. Infect. Dis. 2001, 184, 1098–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaïdane, H.; Gharbi, J.; Lobert, P.-E.; Caloone, D.; Lucas, B.; Sane, F.; Idziorek, T.; Romond, M.-B.; Aouni, M.; Hober, D. Infection of primary cultures of murine splenic and thymic cells with coxsackievirus B4. Microbiol. Immunol. 2008, 52, 40–46. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT-qPCR Primers | ||
|---|---|---|
| Gene | Sequence 5′-3′ | PCR Product Size (bp) |
| Hprt | Forward: TTATCAGACTGAAGAGCTACTGTAATG Reverse: CTTCAACAATCAAGACATTCTTTCC | 108 |
| Total Igf2 | Forward: GGGAGCTTGTGGACACGC Reverse: GCACTCTTCCACGATGCCA | 107 |
| Igf2 V1 | Forward: CCGGCTTCCAGGTACCAAT Reverse: GCAGCGATGCAGCACAAG | 91 |
| Igf2 V2 | Forward: GCCCTTCTCCTCCGATCCT Reverse: ATGAGAAGCACCAACATCGACTT | 99 |
| Igf2 V3 | Forward: CCAGCCTTTTCCTGTCTTCATC reverse: CCATTGGTACCTGAAGTTGGGTAA | 69 |
| Birc5 | Forward: TCTGGCAGCTGTACCTCAAGAACT Reverse: AAACACTGGGCCAAATCAGGCT | 148 |
| Tp53 | Forward: TTCATTGGGACCATCCTGGC Reverse: TGGCAGTCATCCAGTCTTCG | 121 |
| Bcl2 | Forward: GTGAACTGGGGGAGGATTGT Reverse: GGAGAAATCAAACAGAGGCC | 216 |
| Il6 | Forward: GTTCTCTGGGAAATCGTGGA Reverse: TGTACTCCAGGTAGCTATGG | 208 |
| Cxadr | Forward: GTCTAGTCGCAGCATACAC | 287 |
| Reverse: TTCCTGCTGACCGTTCTTG | ||
| Aire | Forward: GGGACTGGTTTAGGTCCACA | 326 |
| Reverse: AGGTGGGGATGGAATGCTAC | ||
| Beta actin | Forward: ATGCTCCCCGGGCTGTAT | 87 |
| Reverse: CATAGGAGTCCTTCTGACCCATTC | ||
| Site-directed mutagenesis primers | ||
| Igf2 P3 Plasmid (Sequence Relative to TSS) | Primer | Annealing (°C) |
| P243 (−68/+175) | Forward: GGTAGGGTGGAGCCGGGA Reverse: GAGCTCAGGTACCGGCCA | 68 |
| P149 (+26/+175) | Forward: AACCTTCCAGCCTTTTCCTGT Reverse: GAGCTCAGGTACCGGCCA | 66 |
| P98 (+77/+175) | Forward: TTACCCAACTTCAGGTAACCAGG Reverse: GAGCTCAGGTACCGGCCA | 70 |
| P197 (−22/+175) | Forward: GGAGGCACTGACCAGTTCG Reverse: GAGCTCAGGTACCGGCCA | 67 |
| P169 (+6/+175) | Forward: ACATTAGCTTCTCCTGTGAGAACC Reverse: GAGCTCAGGTACCGGCCA | 65 |
| P291 (−116/+175) | Forward: GCGGGTGCAAAGGGGGCG Reverse: GAGCTCAGGTACCGGCCAGTTAG | 70 |
| P230 (−55/+175) | Forward: GGGACTGGGAGGAGCCAC Reverse: GAGCTCAGGTACCGGCCA | 71 |
| P248 * (−167/−116Δ−116/−22−22/+175) | Forward: CGGAGGCACTGACCAGTTC Reverse: CCCCTCCCTCCTTCCAGC | 67 |
| P307 * (−167/−151Δ−151/−116−116/+175) | Forward: GGCGGGTGCAAAGGGGGC Reverse: CACCCCCAAGAGCTAGCGAGC | 69 |
| E2FX3 | Forward: GGAAGTGGCTGGAAGTGGCTGGAAGGAGGGAGG | 66 |
| Sequencing primer | Forward: CTAGCAAAATAGGCTGTCCC Reverse: ACTGCATTCTAGTTGTGGTTTGC |
| Primary Antibody | Dilution | Supplier | Clone |
|---|---|---|---|
| Rabbit anti-IGF2 | 1:500 | AVIVA SYSTEMS BIOLOGY | OAAB07463 |
| Rabbit anti-STAT3 | 1:1000 | Cell Signaling Technology | D3Z2G |
| Rabbit anti-STAT3 (p705 Tyr) | 1:1000 | Cell Signaling Technology | D3A7 |
| Rabbit anti-JNK/SAPK | 1:1000 | Cell Signaling Technology | |
| Rabbit anti-pJNK | 1:1000 | Cell Signaling Technology | 81E11 |
| Rabbit anti-ERK (1/2) | 1:1000 | Cell Signaling Technology | 137F5 |
| Rabbit anti-pERK (1/2) | 1:1000 | Cell Signaling Technology | Antirabbit IgG HRP |
| Rabbit anti-SOCS3 | 1:1000 | Cell Signaling Technology | Antirabbit IgG HRP |
| Mouse anti-GAPDH | 1:1000 | Pierce | GA1R |
| Mouse anti-β tubulin | 1:1000 | Pierce | BT7R |
| Mouse anti-VP1 | 1:1000 | Dako | 5-D8/1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michaux, H.; Halouani, A.; Trussart, C.; Renard, C.; Jaïdane, H.; Martens, H.; Geenen, V.; Hober, D. Modulation of IGF2 Expression in the Murine Thymus and Thymic Epithelial Cells Following Coxsackievirus-B4 Infection. Microorganisms 2021, 9, 402. https://doi.org/10.3390/microorganisms9020402
Michaux H, Halouani A, Trussart C, Renard C, Jaïdane H, Martens H, Geenen V, Hober D. Modulation of IGF2 Expression in the Murine Thymus and Thymic Epithelial Cells Following Coxsackievirus-B4 Infection. Microorganisms. 2021; 9(2):402. https://doi.org/10.3390/microorganisms9020402
Chicago/Turabian StyleMichaux, Hélène, Aymen Halouani, Charlotte Trussart, Chantal Renard, Hela Jaïdane, Henri Martens, Vincent Geenen, and Didier Hober. 2021. "Modulation of IGF2 Expression in the Murine Thymus and Thymic Epithelial Cells Following Coxsackievirus-B4 Infection" Microorganisms 9, no. 2: 402. https://doi.org/10.3390/microorganisms9020402