
Antibiotic Resistance and Phylogeny of Pseudomonas spp. Isolated over Three Decades from Chicken Meat in the Norwegian Food Chain


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample and Strain Collection
2.2. 16S rRNA Gene Analyses
2.3. 16 S rDNA Phylogenetic Analyses
2.4. Antimicrobial Susceptibility Testing
2.5. Integron-PCR
2.6. Whole-Genome Sequencing (WGS)
Analyses of WGS Data
2.7. Statistical Analyses
3. Results
3.1. Overall Identification and 16S rRNA Gene Phylogenetic Analyses of Bacteria from Chicken Samples
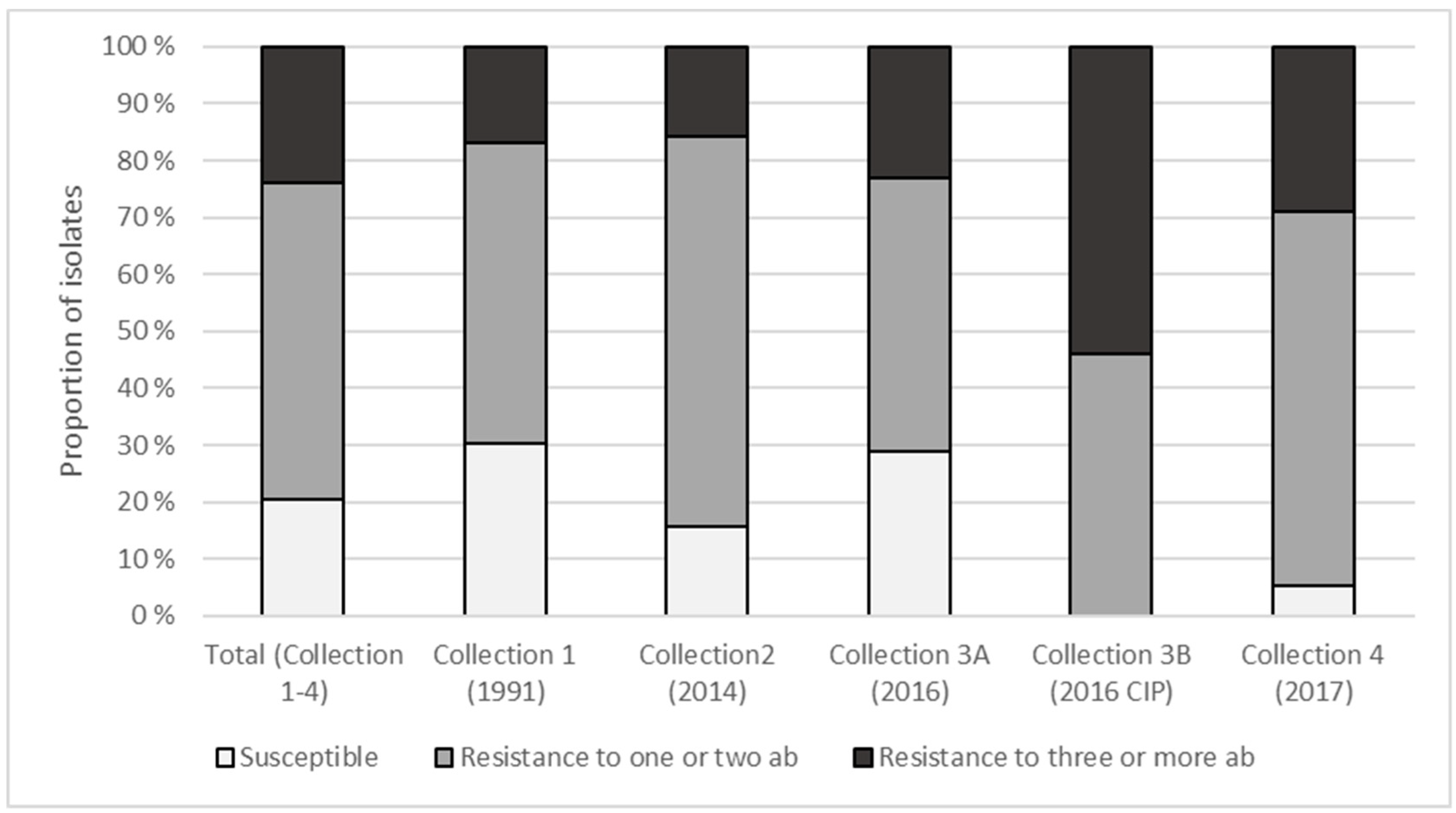
3.2. Phenotypic Antimicrobial Resistance and Presence of Integrons
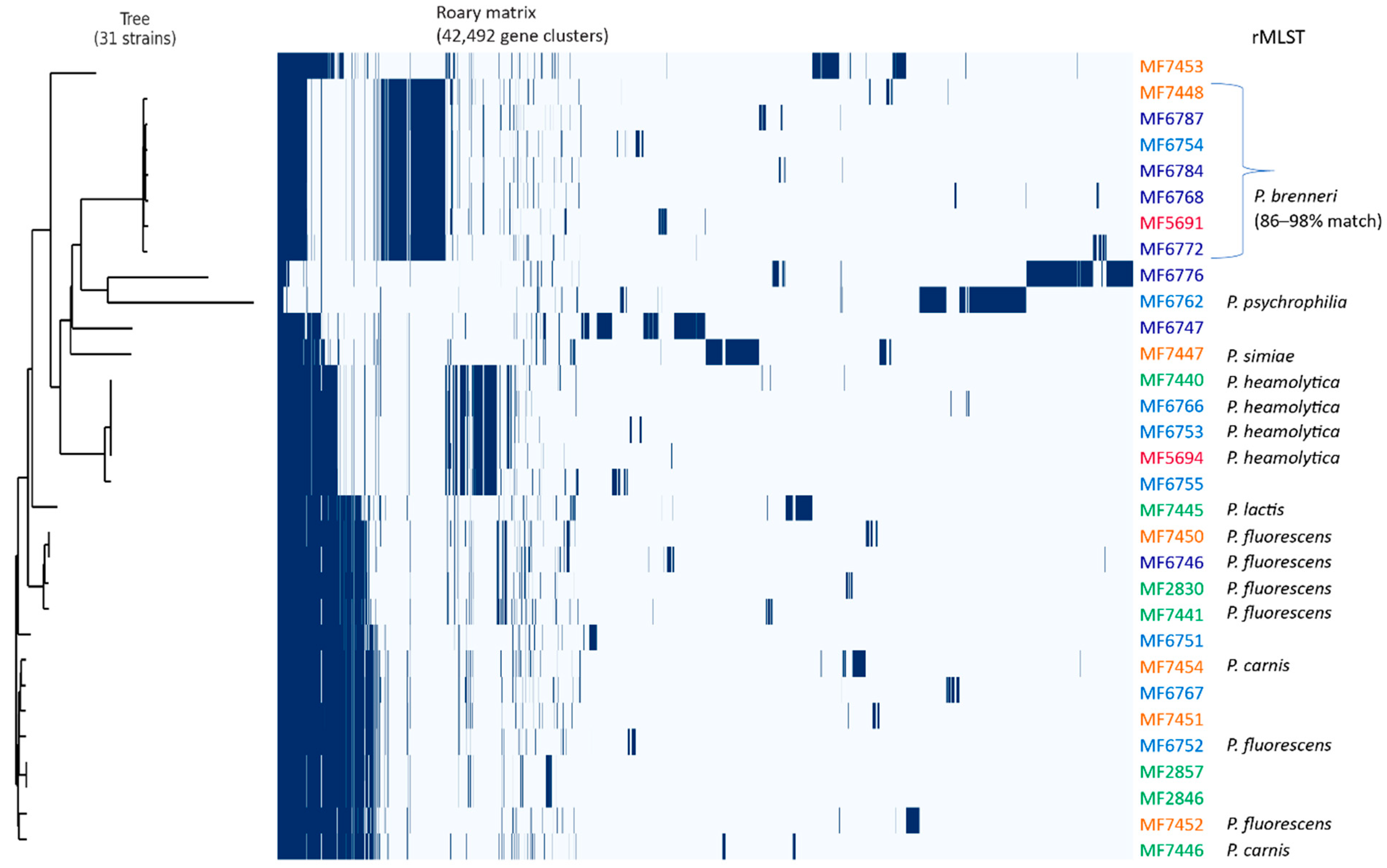
3.3. WGS of a Subset of Resistant Pseudomonas spp.
3.4. Antimicrobial Resistance Determinants in Pseudomonas spp.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peix, A.; Ramírez-Bahena, M.-H.; Velázquez, E. Historical Evolution and Current Status of the Taxonomy of Genus Pseudomonas. Infect. Genet. Evol. 2009, 9, 1132–1147. [Google Scholar] [CrossRef] [PubMed]
- Peix, A.; Ramírez-Bahena, M.H.; Velázquez, E. The Current Status on the Taxonomy of Pseudomonas Revisited: An Update. Infect. Genet. Evol. 2018, 57, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, S.; Anderson, G. The Genus Pseudomonas; Informa UK Limited: London, UK, 2015; pp. 321–344. [Google Scholar]
- Hilgarth, M.; Lehner, E.M.; Behr, J.; Vogel, R.F. Diversity and Anaerobic Growth of Pseudomonasspp. Isolated from Modified Atmosphere Packaged Minced Beef. J. Appl. Microbiol. 2019, 127, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Jeukens, J.; Freschi, L.; Kukavica-Ibrulj, I.; Emond-Rheault, J.-G.; Tucker, N.P.; Lévesque, R. Genomics of Antibiotic-Resistance Prediction in Pseudomonas Aeruginosa. Ann. N. Y. Acad. Sci. 2017, 1435, 5–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, A.; Cason, J.; Ingram, K.D. Tracking Spoilage Bacteria in Commercial Poultry Processing and Refrigerated Storage of Poultry Carcasses. Int. J. Food Microbiol. 2004, 91, 155–165. [Google Scholar] [CrossRef]
- Morales, P.A.; Aguirre, J.S.; Troncoso, M.R.; Figueroa, G.O. Phenotypic and Genotypic Characterization of Pseudomonas spp. Present in Spoiled Poultry Fillets Sold in Retail Settings. LWT 2016, 73, 609–614. [Google Scholar] [CrossRef]
- Lee, H.S.; Kwon, M.; Heo, S.; Kim, M.G.; Kim, G.-B. Characterization of the Biodiversity of the Spoilage Microbiota in Chicken Meat Using Next Generation Sequencing and Culture Dependent Approach. Food Sci. Anim. Resour. 2017, 37, 535–541. [Google Scholar] [CrossRef]
- Sundheim, G.; Sletten, A.; Dainty, R.H. Identification of Pseudomonads from Fresh and Chill-Stored Chicken Carcasses. Int. J. Food Microbiol. 1998, 39, 185–194. [Google Scholar] [CrossRef]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in Bacterial Taxonomy: Impact on the Genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Liu, H.; Lan, T.; Dong, L.; Hu, H.; Zhao, S.; Zhang, Y.; Zheng, N.; Wang, J. Antibiotic Resistance Patterns of Pseudomonas spp. Isolated from Raw Milk Revealed by Whole Genome Sequencing. Front. Microbiol. 2020, 11, 1005. [Google Scholar] [CrossRef]
- Quintieri, L.; Fanelli, F.; Caputo, L. Antibiotic Resistant Pseudomonas Spp. Spoilers in Fresh Dairy Products: An Underestimated Risk and the Control Strategies. Foods 2019, 8, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møretrø, T.; Langsrud, S. Residential Bacteria on Surfaces in the Food Industry and Their Implications for Food Safety and Quality. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1022–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amos, G.C.A.; Carter, A.; Hawkey, P.M.; Gaze, W.H.; Wellington, E. The Hidden Resistome of Retail Chicken Meat. J. Glob. Antimicrob. Resist. 2015, 3, 44–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.H.-Y.; Chan, E.W.C.; Chen, S. Isolation of Carbapenem-Resistant Pseudomonas spp. from Food. J. Glob. Antimicrob. Resist. 2015, 3, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Decimo, M.; Silvetti, T.; Brasca, M. Antibiotic Resistance Patterns of Gram-Negative Psychrotrophic Bacteria from Bulk Tank Milk. J. Food Sci. 2016, 81. [Google Scholar] [CrossRef] [PubMed]
- Estepa, V.; Rojo-Bezares, B.; Torres, C.; Sáenz, Y. Genetic Lineages and Antimicrobial Resistance in Pseudomonasspp. Isolates Recovered from Food Samples. Foodborne Pathog. Dis. 2015, 12, 486–491. [Google Scholar] [CrossRef]
- Breidenstein, E.B.; De La Fuente-Nunez, C.; Hancock, R.E. Pseudomonas Aeruginosa: All Roads Lead to Resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority) and ECDC (European Centre for Disease, Prevention and Control). The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2017/2018. EFSA J. 2020, 18. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency, European Surveillance of Veterinary Antimicrobial Consumption, 2020. Sales of Veterinary Antimicrobial agents in 31 European Countries in 2018. (EMA/24309/2020). Available online: https://www.ema.europa.eu/en/veterinary-regulatory/overview/antimicrobial-resistance/european-surveillance-veterinary-antimicrobial-consumption-esvac (accessed on 20 December 2020).
- Staff, T.P.O.; Staff, P.O. Correction: Occurrence and Characterization of Quinolone Resistant Escherichia Coli from Norwegian Turkey Meat and Complete Sequence of an IncX1 Plasmid Encoding qnrS1. PLoS ONE 2019, 14, e0217321. [Google Scholar] [CrossRef]
- NORM/NORM-VET. Usage of Antimicrobial Agents and Occurence of Antimicrobial Resistance in Norway; Norwegian Veterinary Institute/University Hospital of North Norway: Tromsø/Oslo, Norway, 2014. [Google Scholar]
- NORM/NORM-VET. Usage of Antimicrobial Agents and Occurence of Antimicrobial Resistance in Norway; Norwegian Veterinary Institute/University Hospital of North Norway: Tromsø/Oslo, Norway, 2015. [Google Scholar]
- NORM/NORM-VET. Usage of Antimicrobial Agents and Occurence of Antimicrobial Resistance in Norway; Norwegian Veterinary Institute/University Hospital of North Norway: Tromsø/Oslo, Norway, 2016. [Google Scholar]
- Mo, S.S.; Sunde, M.; Ilag, H.K.; Langsrud, S.; Heir, E. Transfer Potential of Plasmids Conferring Extended-Spectrum-Cephalosporin Resistance in Escherichia coli from Poultry. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [Green Version]
- Sunde, M. Prevalence and Characterization of Class 1 and Class 2 Integrons in Escherichia Coli Isolated from Meat and Meat Products of Norwegian Origin. J. Antimicrob. Chemother. 2005, 56, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Aronesty, E. Command-Line Tools for Processing Biological Sequencing Data. 2011. Available online: https://scholar.google.com/citations?user=RzuLHK5Or2QJ&hl=en (accessed on 20 December 2020).
- Bankevich, A.; Nurk, S.; Pyshkin, A.V.; Sirotkin, A.V.; Vyahhi, N.; Tesler, G.; Alekseyev, M.A.; Pevzner, P.A.; Antipov, D.; Gurevich, A.A.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid Large-Scale Prokaryote Pan Genome Analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.; Bliss, C.M.; Maiden, M.C.J.; Bennett, J.S.; Bratcher, H.B.; Brehony, C.; Colles, F.M.; Wimalarathna, H.; Harrison, O.B.; Sheppard, S.K.; et al. Ribosomal Multilocus Sequence Typing: Universal Characterization of Bacteria from Domain to Strain. Microbiology 2012, 158, 1005–1015. [Google Scholar] [CrossRef]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-Time Whole-Genome Sequencing for Routine Typing, Surveillance, and Outbreak Detection of Verotoxigenic Escherichia Coli. J. Clin. Microbiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the Problem of Comparing Whole Bacterial Genomes across Different Sequencing Platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- Leekitcharoenphon, P.; Nielsen, E.M.; Kaas, R.S.; Lund, O.; Aarestrup, F.M. Evaluation of Whole Genome Sequencing for Outbreak Detection of Salmonella enterica. PLoS ONE 2014, 9, e87991. [Google Scholar] [CrossRef] [Green Version]
- Handley, J.A.; Park, S.H.; Kim, S.A.; Ricke, S. Microbiome Profiles of Commercial Broilers Through Evisceration and Immersion Chilling during Poultry Slaughter and the Identification of Potential Indicator Microorganisms. Front. Microbiol. 2018, 9, 345. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Fegan, N.; Kocharunchitt, C.; Bowman, J.P.; Duffy, L.L. Changes of the Bacterial Community Diversity on Chicken Carcasses through an Australian Poultry Processing Line. Food Microbiol. 2020, 86, 103350. [Google Scholar] [CrossRef] [PubMed]
- Rouger, A.; Moriceau, N.; Prévost, H.; Remenant, B.; Zagorec, M. Diversity of Bacterial Communities in French Chicken Cuts Stored under Modified Atmosphere Packaging. Food Microbiol. 2018, 70, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Lerma, L.L.; Benomar, N.; Muñoz, M.D.C.C.; Gálvez, A.; Abriouel, H. Antibiotic Multiresistance Analysis of Mesophilic and Psychrotrophic Pseudomonas spp. Isolated from Goat and Lamb Slaughterhouse Surfaces throughout the Meat Production Process. Appl. Environ. Microbiol. 2014, 80, 6792–6806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, H.; Franzetti, L.; Kaushal, A.; Kumar, D. Pseudomonas Fluorescens: A Potential Food Spoiler and Challenges and Advances in its Detection. Ann. Microbiol. 2019, 69, 873–883. [Google Scholar] [CrossRef]
- Tran, Q.T.; Nawaz, M.S.; Deck, J.; Nguyen, K.T.; Cerniglia, C.E. Plasmid-Mediated Quinolone Resistance in Pseudomonas putidaIsolates from Imported Shrimp. Appl. Environ. Microbiol. 2010, 77, 1885–1887. [Google Scholar] [CrossRef] [Green Version]
- Hernández, A.; Sanchez, M.B.; Martínez, J.L. Quinolone Resistance: Much More than Predicted. Front. Microbiol. 2011, 2, 22. [Google Scholar] [CrossRef] [Green Version]
- Strand, L.; Jenkins, A.; Henriksen, I.H.; Allum, A.G.; Grude, N.; Kristiansen, B.-E. High Levels of Multiresistance in Quinolone Resistant Urinary Tract Isolates of Escherichia Coli from Norway; A Non Clonal Phenomen? BMC Res. Notes 2014, 7, 376. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Roldán, L.; Rojo-Bezares, B.; De Toro, M.; López, M.; Toledano, P.; Lozano, C.; Chichón, G.; Alvarez-Erviti, L.; Torres, C.; Sáenz, Y. Antimicrobial Resistance and Virulence of Pseudomonas spp. Among Healthy Animals: Concern about Exolysin ExlA Detection. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Jorth, P.; McLean, K.; Penewit, K.; Waalkes, A.; Singh, P.K.; Salipante, S.; Ratjen, A.; Secor, P.R.; Bautista, G.E.; Ravishankar, S.; et al. Evolved Aztreonam Resistance Is Multifactorial and Can Produce Hypervirulence in Pseudomonas Aeruginosa. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.T.; Sinha, U.; Hwang, D.M.; Zhang, Y.; Wang, P.W.; Donaldson, S.L.; Coburn, B.; Waters, V.J.; Yau, Y.C.; Tullis, D.E.; et al. Penicillin-Binding Protein 3 is a Common Adaptive Target Among Pseudomonas Aeruginosa Isolates from Adult Cystic Fibrosis Patients Treated with β-lactams. Int. J. Antimicrob. Agents 2019, 53, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Arslan, S.; Eyi, A.; Özdemir, F. Spoilage Potentials and Antimicrobial Resistance of Pseudomonas spp. Isolated from Cheeses. J. Dairy Sci. 2011, 94, 5851–5856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Palmieri, M.; Brilhante, M.; Masseron, A.; Perreten, V.; Nordmann, P. PFM-Like Enzymes Are a Novel Family of Subclass B2 Metallo-& β;-Lactamases from Pseudomonas synxantha Belonging to the Pseudomonas fluorescens Complex. Antimicrob. Agents Chemother. 2019, 64. [Google Scholar] [CrossRef]
- Nielsen, K.M.; Smalla, K.; Van Elsas, J.D. Natural Transformation of Acinetobactersp. Strain BD413 with Cell Lysates of Acinetobacter sp., Pseudomonas fluorescens, and Burkholderia Cepaciain Soil Microcosms. Appl. Environ. Microbiol. 2000, 66, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Peter, S.; Oberhettinger, P.; Willmann, M.; Schuele, L.; Dinkelacker, A.; Vogel, W.; Dörfel, D.; Bezdan, D.; Ossowski, S.; Marschal, M.; et al. Genomic Characterisation of Clinical and Environmental Pseudomonas Putida Group Strains and Determination of their Role in the Transfer of Antimicrobial Resistance Genes to Pseudomonas Aeruginosa. BMC Genom. 2017, 18, 1–11. [Google Scholar] [CrossRef]
- Oliver, A.; Torrens, G.; González-Nicolau, M.; Oliver, A. Diversity and Regulation of Intrinsic & β;-lactamases from Non-Fermenting and other Gram-Negative Opportunistic Pathogens. FEMS Microbiol. Rev. 2017, 41, 781–815. [Google Scholar] [CrossRef]
- Tacão, M.; Correia, A.; Henriques, I. Low Prevalence of Carbapenem-Resistant Bacteria in River Water: Resistance Is Mostly Related to Intrinsic Mechanisms. Microb. Drug Resist. 2015, 21, 497–506. [Google Scholar] [CrossRef]
- Fang, Z.-L.; Zhang, L.-Y.; Huang, Y.-M.; Qing, Y.; Cao, K.-Y.; Tian, G.-B.; Huang, X. OprD Mutations and Inactivation in Imipenem-Resistant Pseudomonas Aeruginosa Isolates from China. Infect. Genet. Evol. 2014, 21, 124–128. [Google Scholar] [CrossRef]
- Wolter, D.J.; Lister, P.D. Mechanisms of Beta-Lactam Resistance Among Pseudomonas Aeruginosa. Curr. Pharm. Des. 2013, 19, 209–222. [Google Scholar] [CrossRef]
- Chevalier, S.; Bodilis, J.; Jaouen, T.; Barray, S.; Feuilloley, M.; Orange, N. Sequence Diversity of the OprD Protein of Environmental Pseudomonas Strains. Environ. Microbiol. 2007, 9, 824–835. [Google Scholar] [CrossRef]
- Bonardi, S.; Pitino, R. Carbapenemase-Producing Bacteria in Food-Producing Animals, Wildlife and Environment: A Challenge for Human Health. Ital. J. Food Saf. 2019, 8, 7956. [Google Scholar] [CrossRef] [PubMed]
- Moubareck, C.A. Polymyxins and Bacterial Membranes: A Review of Antibacterial Activity and Mechanisms of Resistance. Membr. 2020, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.A.E.-G.E.-S.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and its Role in the Era of Antibiotic Resistance: An Extended Review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Weigand, M.R.; Oh, S.; Hatt, J.K.; Krishnan, R.; Tezel, U.; Pavlostathis, S.G.; Konstantinidis, K.T. Widely Used Benzalkonium Chloride Disinfectants Can Promote Antibiotic Resistance. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasr, A.M.; Mostafa, M.S.; Arnaout, H.H.; Elshimy, A.A.A. The Effect of Exposure to Sub-Inhibitory Concentrations of Hypochlorite and Quaternary Ammonium Compounds on Antimicrobial Susceptibility of Pseudomonas Aeruginosa. Am. J. Infect. Control. 2018, 46, e57–e63. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-Z.; Plésiat, P.; Nikaido, H. The Challenge of Efflux-Mediated Antibiotic Resistance in Gram-Negative Bacteria. Clin. Microbiol. Rev. 2015, 28, 337–418. [Google Scholar] [CrossRef] [Green Version]
- Dumas, J.-L.C.; Van Delden, C.; Perron, K.; Köhler, T. Analysis of Antibiotic Resistance Gene Expression in Pseudomonas Aeruginosaby Quantitative Real-Time-PCR. FEMS Microbiol. Lett. 2006, 254, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Jeannot, K.; Bolard, A.; Plésiat, P. Resistance to Polymyxins in Gram-negative Organisms. Int. J. Antimicrob. Agents 2017, 49, 526–535. [Google Scholar] [CrossRef]
- Devarajan, N.; Köhler, T.; Sivalingam, P.; Van Delden, C.; Mulaji, C.; Mpiana, P.T.; Ibelings, B.W.; Poté, J. Antibiotic Resistant Pseudomonas spp. in the Aquatic Environment: A Prevalence Study under Tropical and Temperate Climate Conditions. Water Res. 2017, 115, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Jové, T.; Da Re, S.; Denis, F.; Mazel, D.; Ploy, M.-C. Inverse Correlation between Promoter Strength and Excision Activity in Class 1 Integrons. PLoS Genet. 2010, 6, e1000793. [Google Scholar] [CrossRef] [Green Version]
- Heuer, H.; Binh, C.T.T.; Jechalke, S.; Kopmann, C.; Zimmerling, U.; Krögerrecklenfort, E.; Ledger, T.; González, B.; Top, E.M.; Smalla, K. IncP-1& ε Plasmids are Important Vectors of Antibiotic Resistance Genes in Agricultural Systems: Diversification Driven by Class 1 Integron Gene Cassettes. Front. Microbiol. 2012, 3, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.S.; Bruun, M.S.; Dalsgaard, I.; Larsen, J.L. Incidence, Distribution, and Spread of Tetracycline Resistance Determinants and Integron-Associated Antibiotic Resistance Genes among Motile Aeromonads from a Fish Farming Environment. Appl. Environ. Microbiol. 2001, 67, 5675–5682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillings, M.R. Integrons: Past, Present, and Future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, S.S.; Norström, M.; Slettemeås, J.S.; Løvland, A.; Urdahl, A.M.; Sunde, M. Emergence of AmpC-producing Escherichia Coli in the Broiler Production Chain in a Country with a Low Antimicrobial Usage Profile. Veter Microbiol. 2014, 171, 315–320. [Google Scholar] [CrossRef] [PubMed]
- NORM/NORM-VET 2018. Usage of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Norway; Norwegian Veterinary Institute/University Hospital of North Norway: Tromsø/Oslo, Norway, 2018. [Google Scholar]
- Grave, K.; Kaldhusdal, M.C.; Kruse, H.; Harr, L.M.; Flatlandsmo, K. What has Happened in Norway after the Ban of Avoparcin? Consumption of Antimicrobials by Poultry. Prev. Veter Med. 2004, 62, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Slettemeås, J.S.; Urdahl, A.M.; Mo, S.S.; Norström, M.; Steinbakk, M.; Johannessen, G.S.; Grave, K.; Sunde, M. Imported Food and Feed as Contributors to the Introduction of Plasmid-Mediated Colistin-Resistant Enterobacteriaceae to a ‘Low Prevalence’ Country. J. Antimicrob. Chemother. 2017, 72, 2675–2677. [Google Scholar] [CrossRef]
- Scales, B.S.; Dickson, R.P.; Lipuma, J.J.; Huffnagle, G.B. Microbiology, Genomics, and Clinical Significance of the Pseudomonas Fluorescens Species Complex, An Unappreciated Colonizer of Humans. Clin. Microbiol. Rev. 2014, 27, 927948. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Sample Collection (Reference) | Sample Type | Year | Chicken Samples | 16S rRNA Gene | MIC a | WGS b |
|---|---|---|---|---|---|---|
| 1 [9] | Cold stored Carcasses | 1991 | 20 | 97 | 53 | 7 |
| 2 (This study) | Raw fillet at retail | 2014 | 8 | 34 | 19 | 2 |
| 3A (This study) | Raw fillet at retail | 2016 | 60 | 113 | 52 | 7 |
| 3B (This study) | Raw fillet at retail | 2016 | 53 c | 36 | 13 | 8 |
| 4 (This study) | Raw carcass in slaughterhouse | 2017 | 18 | 45 | 38 | 7 |
| Total | 325 | 175 | 31 |
| Antibiotic | Resistance (%) | Distribution (%) of MIC Values (mg/L) a | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | ||
| Amikacin | 0.0 | 99.4 | 0.6 | |||||||||
| Gentamicin | I.E. b | 96.6 | 1.7 | 1.1 | 0.6 | |||||||
| Tobramycin | 0.0 | 1.7 | 98.3 | |||||||||
| Meropenem | 12.6 | 22.3 | 6.9 | 10.9 | 8.0 | 10.9 | 16.6 | 12.0 | 8.6 | 4.0 | ||
| Imipenem | 25.6 | 19.4 | 20.6 | 21.1 | 13.1 | 11.4 | 13.1 | 1.1 | ||||
| Ceftazidime | 10.2 | 1.1 | 5.7 | 29.7 | 33.7 | 19.4 | 5.1 | 5.1 | ||||
| Ceftazidime/Avibactam | 0.6 | 1.1 | 8.6 | 46.9 | 39.4 | 3.4 | 0.6 | |||||
| Ceftolozane/Tazobactam | 0.6 | 20.6 | 52.0 | 21.7 | 5.1 | 0.6 | ||||||
| Ciprofloxacin | 2.2 | 54.9 | 30.3 | 9.7 | 2.9 | 1.1 | 1.1 | |||||
| Aztreonam | 72.6 | 0.6 | 0.0 | 0.6 | 13.7 | 12.6 | 12.0 | 60.6 | ||||
| Piperacillin/Tazobactam | 0.6 | 1.1 | 12.0 | 42.9 | 34.9 | 8.6 | 0.6 | |||||
| Colistin | 30.2 | 1.1 | 23.4 | 36.6 | 8.6 | 7.4 | 5.1 | 17.7 | ||||
| Best Match (Species) 1 | Isolates (N) | MIC (N) | Proportion of Isolates (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ab = 0 | ab = 1 | ab = 2 | ab = 3 | ab = 4 | ab = 5 | ab = 7 | |||
| P. brassicacearum | 1 | 1 | 0.0 | 0.0 | 100 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. brenneri | 1 | 1 | 0.0 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. caeni2 | 1 | 0 | |||||||
| P. canadensis | 22 | 11 | 0.0 | 0.0 | 9.1 | 54.5 | 27.3 | 0.0 | 9.1 |
| P. cerasi | 16 | 10 | 50.0 | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. corrugata | 13 | 9 | 0.0 | 66.7 | 22.2 | 0.0 | 0.0 | 11.1 | 0.0 |
| P. deceptionensis | 2 | 2 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. flavescens | 2 | 2 | 50.0 | 50.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. fragi | 13 | 8 | 87.5 | 12.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. gessardii | 68 | 26 | 0.0 | 53.8 | 15.4 | 11.5 | 19.2 | 0.0 | 0.0 |
| P. helleri | 6 | 4 | 25.0 | 75.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. helmanticensis | 4 | 3 | 0.0 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. koreensis | 7 | 5 | 0.0 | 60.0 | 40.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. lactis | 57 | 26 | 0.0 | 0.0 | 15.4 | 61.5 | 23.1 | 0.0 | 0.0 |
| P. marginalis | 11 | 11 | 0.0 | 18.2 | 72.7 | 0.0 | 9.1 | 0.0 | 0.0 |
| P. migulae | 11 | 10 | 0.0 | 100.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. rhodesiae | 1 | 1 | 0.0 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. silesiensis | 5 | 4 | 25.0 | 75.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. tolaasii | 6 | 4 | 0.0 | 75.0 | 25.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. turukhanskensis | 3 | 3 | 0.0 | 33.3 | 66.7 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. veronii | 22 | 9 | 0.0 | 88.9 | 11.1 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. versuta | 1 | 1 | 0.0 | 100 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| P. weihenstephanensis | 52 | 24 | 79.2 | 20.8 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| SUM | 325 | 175 | 20.6 | 40.6 | 14.9 | 14.3 | 8.6 | 0.6 | 0.6 |
| Antibiotic Class/Type of Antimicrobial | Phenotypic Resistance a | Resistance Determinants (No. of Isolates) b |
|---|---|---|
| Antibiotic class | ||
| Aminocoumarine | Aminocoumarine | mdtA (31), mdtB (31), mdtC (30) |
| Aminoglycoside | NEO | neo (18) |
| β-lactam | IMI, MERO, β-lactam antibiotics | ampC (31), ampD (31), cphA (1), pbpC (10), mrcA (31), mrcB (9), oprD (31) |
| Bicyclomycin | BC | bcr (29) |
| Fluoroquionolones | CIP | Bmr3 (2), gyrA (31), gyrB (31), lfrA (6), mfd (31), parC (31), parE (31) |
| Fosfomycin | FOS | abaF (31), fosA (13), mdtD (30), mdtG (13) |
| Fosmidomycin | FOSM | fsr (22) |
| Lipopeptides | COL | arnA (30), emrA (28), emrB (27), lpxA (2), lpxD (4), pgsA (2), phoP (31), phoQ (9) |
| Macrolides | Macrolides, peptide antibiotics | macA (31), macB (31) |
| Tetracycline | tetC (7) | |
| Disinfectants and cationic agents | QAC, EBR, various cationic agents | emrE (2), qacA (1), qacC (1) |
| Multiple drugsc | Cationic peptides, fluoroquinolones, aminoglycosides | acrA (12), acrB (15), acrE (2), acrF (29) aphA (28), aphB (3) |
| Carbenicillin, chloramphenicol, erythromycin, novobiocin, streptomycin and tetracycline | arpC (26) | |
| Various multidrugs | bepC (29), bepE (15), bmr3 (2), emrA (31), emrB (30), emrD (5), emrE (2), emrY (24), mdfA (1), mdtE (16), mdtK (31), mdtL (28), mdtN (1), mepA (31), mexA (29), mexB (29), mtrA (19), norM (31), oprJ (5), oprM (29), oqxB7 (31), oqxB10 (27), oqxB13 (1), stp (30), tolC (30) | |
| Aromatic hydrocarbons, fluoroquinolones, beta-lactams except imipenem | ttgA (31), ttgB (31), ttgC (31), ttgI (31) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heir, E.; Moen, B.; Åsli, A.W.; Sunde, M.; Langsrud, S. Antibiotic Resistance and Phylogeny of Pseudomonas spp. Isolated over Three Decades from Chicken Meat in the Norwegian Food Chain. Microorganisms 2021, 9, 207. https://doi.org/10.3390/microorganisms9020207
Heir E, Moen B, Åsli AW, Sunde M, Langsrud S. Antibiotic Resistance and Phylogeny of Pseudomonas spp. Isolated over Three Decades from Chicken Meat in the Norwegian Food Chain. Microorganisms. 2021; 9(2):207. https://doi.org/10.3390/microorganisms9020207
Chicago/Turabian StyleHeir, Even, Birgitte Moen, Anette Wold Åsli, Marianne Sunde, and Solveig Langsrud. 2021. "Antibiotic Resistance and Phylogeny of Pseudomonas spp. Isolated over Three Decades from Chicken Meat in the Norwegian Food Chain" Microorganisms 9, no. 2: 207. https://doi.org/10.3390/microorganisms9020207