Lactobacillus iners and gasseri, Prevotella bivia and HPV Belong to the Microbiological Signature Negatively Affecting Human Reproduction

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Enrolment
2.2. Nucleic Acid Extraction and Library Preparation
2.3. HPV Detection
2.4. Next Generation Sequencing Data Processing
2.5. Statistical Analysis
2.6. Data Availability
3. Results
3.1. Features of the Study Cohort
3.2. HPV Testing
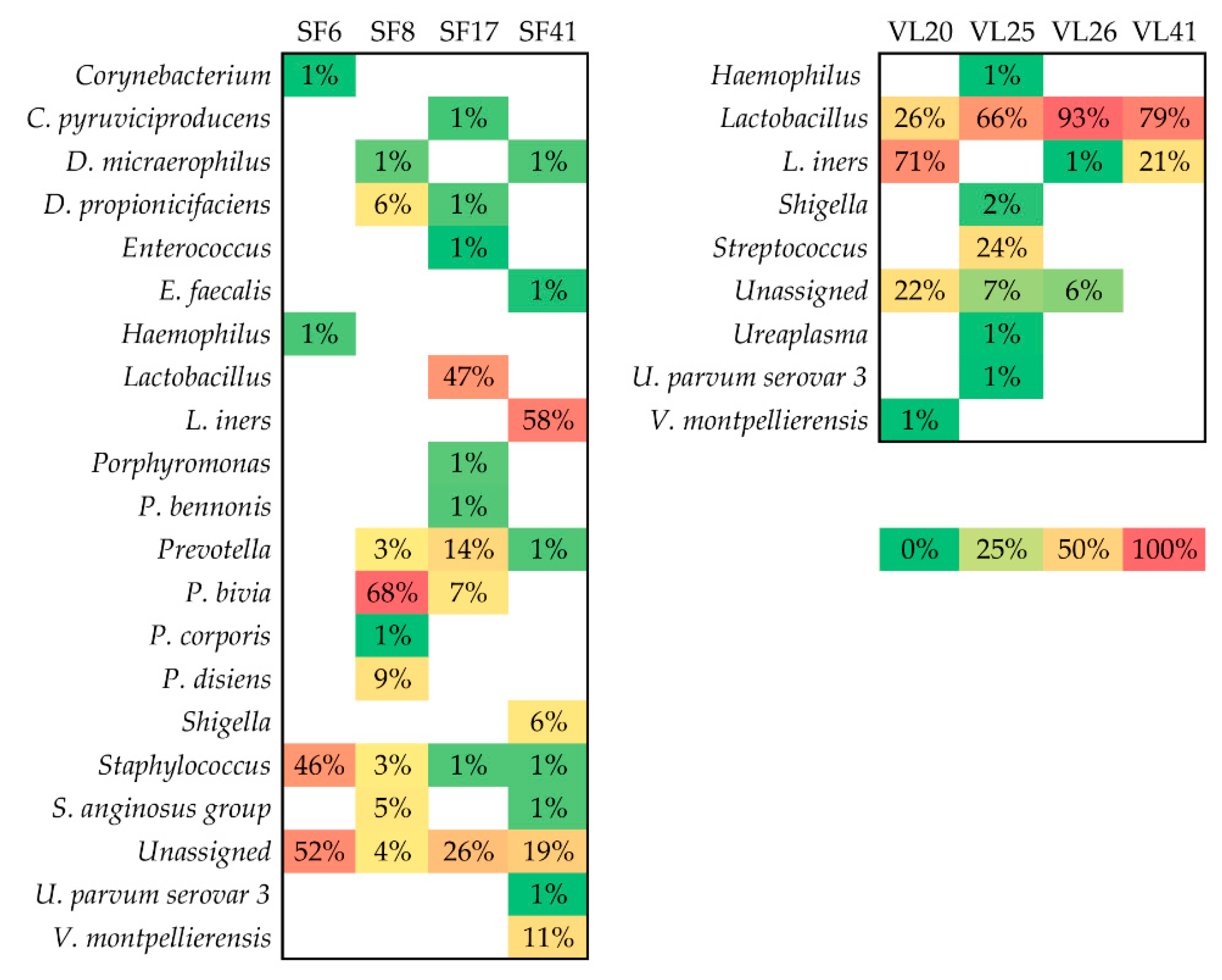
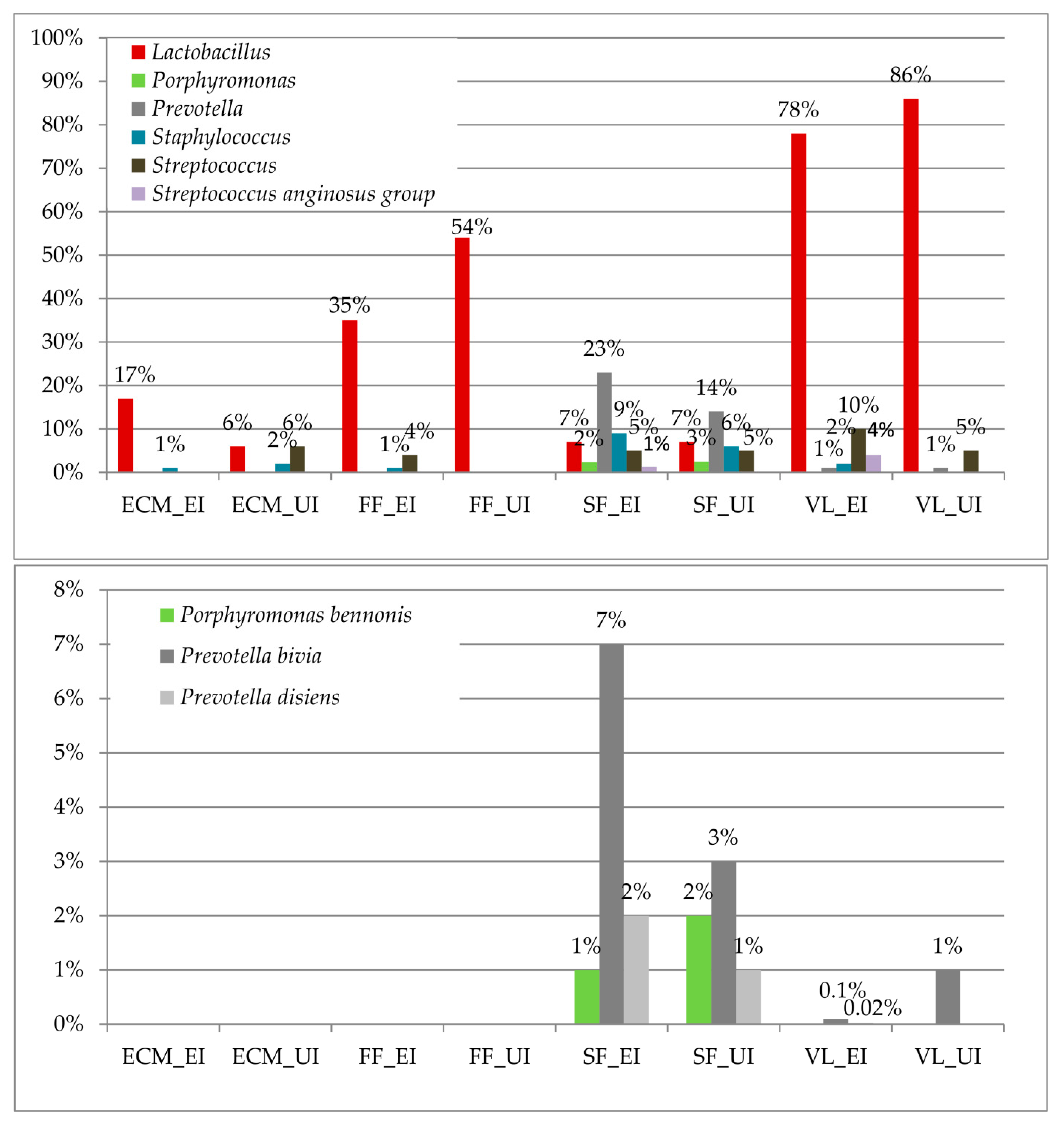
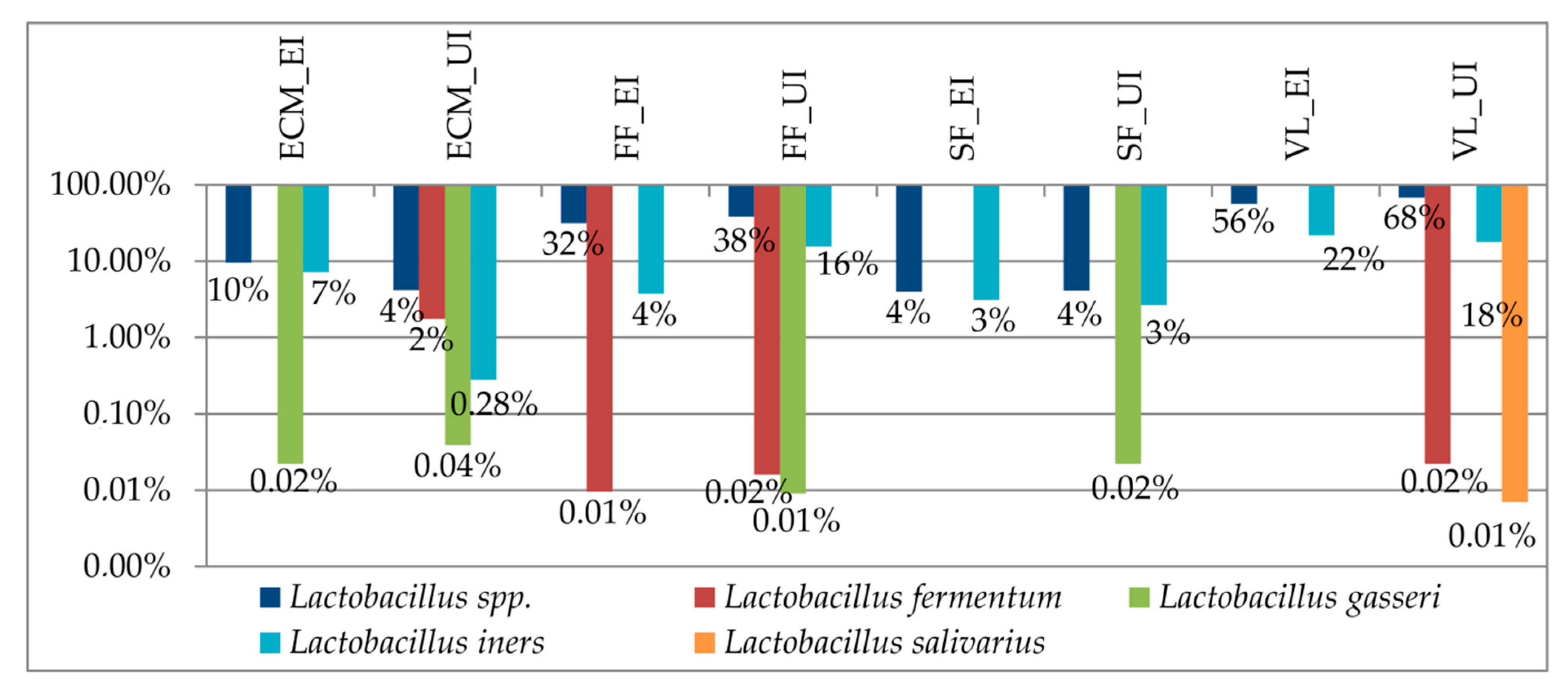
3.3. Taxonomic Biomarkers
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Compliance with Ethical Standards:
References
- Pelzer, E.S.; Allan, J.A.; Cunningham, K.; Mengersen, K.; Allan, J.M.; Launchbury, T.; Beagley, K.; Knox, C.L. Microbial colonization of follicular fluid: Alterations in cytokine expression and adverse assisted reproduction technology outcomes. Hum. Reprod. 2011, 26, 1799–1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, K.A.; Zarek, S.M.; Catherino, W.H. Gynecologic health and disease in relation to the microbiome of the female reproductive tract. Fertil. Steril. 2015, 104, 1351–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franasiak, J.M.; Scott, R.T., Jr. Reproductive tract microbiome in assisted reproductive technologies. Fertil. Steril. 2015, 104, 1364–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graspeuntner, S.; Bohlmann, M.K.; Gillmann, K.; Speer, R.; Kuenzel, S.; Mark, H.; Hoellen, F.; Lettau, R.; Griesinger, G.; Konig, I.R.; et al. Microbiota-based analysis reveals specific bacterial traits and a novel strategy for the diagnosis of infectious infertility. PLoS ONE 2018, 13, e0191047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riganelli, L.; Iebba, V.; Piccioni, M.; Illuminati, I.; Bonfiglio, G.; Neroni, B.; Calvo, L.; Gagliardi, A.; Levrero, M.; Merlino, L.; et al. Structural variations of vaginal and endometrial microbiota: Hints on female infertility. Front. Cell. Infect. Microbiol. 2020, 10, 350. [Google Scholar] [CrossRef]
- Yeruva, T.; Rajkumar, H.; Donugama, V. Vaginal lactobacilli profile in pregnant women with normal & abnormal vaginal flora. Indian J. Med. Res. 2017, 146, 534–540. [Google Scholar]
- Koedooder, R.; Mackens, S.; Budding, A.; Fares, D.; Blockeel, C.; Laven, J.; Schoenmakers, S. Identification and evaluation of the microbiome in the female and male reproductive tracts. Hum. Reprod. Update 2019, 25, 298–325. [Google Scholar] [CrossRef]
- Hou, D.; Zhou, X.; Zhong, X.; Settles, M.L.; Herring, J.; Wang, L.; Abdo, Z.; Forney, L.J.; Xu, C. Microbiota of the seminal fluid from healthy and infertile men. Fertil. Steril. 2013, 100, 1261–1269. [Google Scholar] [CrossRef]
- Monteiro, C.; Marques, P.I.; Cavadas, B.; Damiao, I.; Almeida, V.; Barros, N.; Barros, A.; Carvalho, F.; Gomes, S.; Seixas, S. Characterization of microbiota in male infertility cases uncovers differences in seminal hyperviscosity and oligoasthenoteratozoospermia possibly correlated with increased prevalence of infectious bacteria. Am. J. Reprod. Immunol. 2018, 79, e12838. [Google Scholar] [CrossRef]
- Alfano, M.; Ferrarese, R.; Locatelli, I.; Ventimiglia, E.; Ippolito, S.; Gallina, P.; Cesana, D.; Canducci, F.; Pagliardini, L.; Vigano, P.; et al. Testicular microbiome in azoospermic men-first evidence of the impact of an altered microenvironment. Hum. Reprod. 2018, 33, 1212–1217. [Google Scholar] [CrossRef]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, B.J.; Stojanov, M. Sperm microbiota and its impact on semen parameters. Front. Microbiol. 2019, 10, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandar, R.; Punab, M.; Borovkova, N.; Lapp, E.; Kiiker, R.; Korrovits, P.; Metspalu, A.; Krjutskov, K.; Nolvak, H.; Preem, J.K.; et al. Complementary seminovaginal microbiome in couples. Res. Microbiol. 2015, 166, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Borovkova, N.; Korrovits, P.; Ausmees, K.; Turk, S.; Joers, K.; Punab, M.; Mandar, R. Influence of sexual intercourse on genital tract microbiota in infertile couples. Anaerobe 2011, 17, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.E.; Soules, M.R.; Klein, N.A.; Fujimoto, V.Y.; Agnew, K.J.; Eschenbach, D.A. Bacteria in the transfer catheter tip influence the live-birth rate after in vitro fertilization. Fertil. Steril. 2000, 74, 1118–1124. [Google Scholar] [CrossRef]
- Egbase, P.E.; al-Sharhan, M.; al-Othman, S.; al-Mutawa, M.; Udo, E.E.; Grudzinskas, J.G. Incidence of microbial growth from the tip of the embryo transfer catheter after embryo transfer in relation to clinical pregnancy rate following in-vitro fertilization and embryo transfer. Hum. Reprod. 1996, 11, 1687–1689. [Google Scholar] [CrossRef] [Green Version]
- Fanchin, R.; Harmas, A.; Benaoudia, F.; Lundkvist, U.; Olivennes, F.; Frydman, R. Microbial flora of the cervix assessed at the time of embryo transfer adversely affects in vitro fertilization outcome. Fertil. Steril. 1998, 70, 866–870. [Google Scholar] [CrossRef]
- Koedooder, R.; Singer, M.; Schoenmakers, S.; Savelkoul, P.H.M.; Morre, S.A.; de Jonge, J.D.; Poort, L.; Cuypers, W.; Beckers, N.G.M.; Broekmans, F.J.M.; et al. The vaginal microbiome as a predictor for outcome of in vitro fertilization with or without intracytoplasmic sperm injection: A prospective study. Hum. Reprod. 2019, 34, 1042–1054. [Google Scholar] [CrossRef]
- Garolla, A.; Pizzol, D.; Bertoldo, A.; Menegazzo, M.; Barzon, L.; Foresta, C. Sperm viral infection and male infertility: Focus on hbv, hcv, hiv, hpv, hsv, hcmv, and aav. J. Reprod. Immunol. 2013, 100, 20–29. [Google Scholar] [CrossRef]
- Marci, R.; Gentili, V.; Bortolotti, D.; Lo Monte, G.; Caselli, E.; Bolzani, S.; Rotola, A.; Di Luca, D.; Rizzo, R. Presence of hhv-6a in endometrial epithelial cells from women with primary unexplained infertility. PLoS ONE 2016, 11, e0158304. [Google Scholar] [CrossRef]
- Souho, T.; Benlemlih, M.; Bennani, B. Human papillomavirus infection and fertility alteration: A systematic review. PLoS ONE 2015, 10, e0126936. [Google Scholar] [CrossRef]
- Foresta, C.; Patassini, C.; Bertoldo, A.; Menegazzo, M.; Francavilla, F.; Barzon, L.; Ferlin, A. Mechanism of human papillomavirus binding to human spermatozoa and fertilizing ability of infected spermatozoa. PLoS ONE 2011, 6, e15036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, Y.M.; Lee, J.F.; Huang, H.Y.; Soong, Y.K.; Yang, F.P.; Pao, C.C. The effect of human papillomavirus infection on sperm cell motility. Fertil. Steril. 1997, 67, 1152–1155. [Google Scholar] [CrossRef]
- Foresta, C.; Noventa, M.; De Toni, L.; Gizzo, S.; Garolla, A. Hpv-DNA sperm infection and infertility: From a systematic literature review to a possible clinical management proposal. Andrology 2015, 3, 163–173. [Google Scholar] [CrossRef]
- Depuydt, C.E.; Verstraete, L.; Berth, M.; Beert, J.; Bogers, J.P.; Salembier, G.; Vereecken, A.J.; Bosmans, E. Human papillomavirus positivity in women undergoing intrauterine insemination has a negative effect on pregnancy rates. Gynecol. Obstet. Investig. 2016, 81, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Campisciano, G.; Gheit, T.; De Seta, F.; Cason, C.; Zanotta, N.; Delbue, S.; Ricci, G.; Ferrante, P.; Tommasino, M.; Comar, M. Oncogenic virome benefits from the different vaginal microbiome-immune axes. Microorganisms 2019, 7, 414. [Google Scholar] [CrossRef] [Green Version]
- Matsha, T.; Erasmus, R.; Kafuko, A.B.; Mugwanya, D.; Stepien, A.; Parker, M.I. Human papillomavirus associated with oesophageal cancer. J. Clin. Pathol. 2002, 55, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Rai, M.A.; Khanani, M.R.; Khan, M.N.; Ali, S.H. Hiv-1 subtype a infection in a community of intravenous drug users in pakistan. BMC Infect. Dis. 2006, 6, 164. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using qiime 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Fettweis, J.M.; Serrano, M.G.; Sheth, N.U.; Mayer, C.M.; Glascock, A.L.; Brooks, J.P.; Jefferson, K.K.; Buck, G.A. Species-level classification of the vaginal microbiome. BMC Genom. 2012, 13 (Suppl. 8), S17. [Google Scholar] [CrossRef] [Green Version]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Munoz, M.E.; Arrieta, M.C.; Ramer-Tait, A.E.; Walter, J. A critical assessment of the “sterile womb” and “in utero colonization” hypotheses: Implications for research on the pioneer infant microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Feng, X.; Li, N.; Zhao, W.; Wei, L.; Chen, Y.; Yang, W.; Ma, H.; Yao, B.; Zhang, K.; et al. Human papillomavirus in semen and the risk for male infertility: A systematic review and meta-analysis. BMC Infect. Dis. 2017, 17, 714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ault, K.A. Epidemiology and natural history of human papillomavirus infections in the female genital tract. Infect. Dis. Obstet. Gynecol. 2006, 2006, 40470. [Google Scholar] [CrossRef] [PubMed]
- Selman, H.; Mariani, M.; Barnocchi, N.; Mencacci, A.; Bistoni, F.; Arena, S.; Pizzasegale, S.; Brusco, G.F.; Angelini, A. Examination of bacterial contamination at the time of embryo transfer, and its impact on the ivf/pregnancy outcome. J. Assist. Reprod. Genet. 2007, 24, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Jakobsson, T.; Forsum, U. Lactobacillus iners: A marker of changes in the vaginal flora? J. Clin. Microbiol. 2007, 45, 3145. [Google Scholar] [CrossRef] [Green Version]
- Ferris, M.J.; Norori, J.; Zozaya-Hinchliffe, M.; Martin, D.H. Cultivation-independent analysis of changes in bacterial vaginosis flora following metronidazole treatment. J. Clin. Microbiol. 2007, 45, 1016–1018. [Google Scholar] [CrossRef] [Green Version]
- Pelzer, E.S.; Harris, J.E.; Allan, J.A.; Waterhouse, M.A.; Ross, T.; Beagley, K.W.; Knox, C.L. Tunel analysis of DNA fragmentation in mouse unfertilized oocytes: The effect of microorganisms within human follicular fluid collected during ivf cycles. J. Reprod. Immunol. 2013, 99, 69–79. [Google Scholar] [CrossRef]
- Pelzer, E.S.; Allan, J.A.; Waterhouse, M.A.; Ross, T.; Beagley, K.W.; Knox, C.L. Microorganisms within human follicular fluid: Effects on ivf. PLoS ONE 2013, 8, e59062. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, T.; Chen, Y.; Luo, T.; Tan, B.; Chen, H.; Xin, H. Evaluation of the inhibitory effects of vaginal microorganisms on sperm motility in vitro. Exp. Ther. Med. 2020, 19, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Stsepetova, J.; Baranova, J.; Simm, J.; Parm, U.; Roop, T.; Sokmann, S.; Korrovits, P.; Jaagura, M.; Rosenstein, K.; Salumets, A.; et al. The complex microbiome from native semen to embryo culture environment in human in vitro fertilization procedure. Reprod. Biol. Endocrinol. 2020, 18, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Van Weert, J.M.; Repping, S.; Van Voorhis, B.J.; van der Veen, F.; Bossuyt, P.M.; Mol, B.W. Performance of the postwash total motile sperm count as a predictor of pregnancy at the time of intrauterine insemination: A meta-analysis. Fertil. Steril. 2004, 82, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Brincat, D.; Catania, S.; Wismayer, P.S.; Calleja-Agius, J. Male factors in art outcome prediction. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2015, 31, 169–175. [Google Scholar] [CrossRef]
- Aroutcheva, A.; Ling, Z.; Faro, S. Prevotella bivia as a source of lipopolysaccharide in the vagina. Anaerobe 2008, 14, 256–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaambo, E.; Africa, C.; Chambuso, R.; Passmore, J.S. Vaginal microbiomes associated with aerobic vaginitis and bacterial vaginosis. Front. Public Health 2018, 6, 78. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| CHAO1 | |||
|---|---|---|---|
| Explained Infertility | Unexplained Infertility | p Value | |
| Vaginal lavages | 42 (95% CI = 36–48) | 42 (95% CI = 35–49) | 0.4 |
| Follicular Fluids | 59 (95% CI = 53–65) | 63 (95% CI = 49–77) | 0.3 |
| Seminal Fluids | 94 (95% CI = 79–109) | 130 (95% CI = 101–159) | 0.08 |
| ECM | 24 (95% CI = 19–29) | 38 (95% CI = 27–49) | 0.1 |
| SHANNON | |||
| Explained Infertility | Unexplained Infertility | p Value | |
| Vaginal lavages | 1 (95% CI = 0.7–1.3) | 1.4 (95% CI = 1.1–1.7) | 0.1 |
| Follicular Fluids | 2.6 (95% CI = 2.2–3) | 2.6 (95% CI = 2.2–3) | 0.7 |
| Seminal Fluids | 3.7 (95% CI = 3.4–4) | 4 (95% CI = 3.7–4.3) | 0.09 |
| ECM | 2.7 (95% CI = 2.4–3) | 3 (95% CI = 2.7–3.3) | 0.4 |
| Genus | LDA Score | p Value |
|---|---|---|
| Lactobacillus | 6.6 | <0.001 |
| Porphyromonas | 5.09 | 0.01 |
| Prevotella | 6.08 | <0.001 |
| Staphylococcus | 5.67 | 0.05 |
| Streptococcus | 5.74 | 0.01 |
| Streptococcus anginosus group | 5.27 | <0.001 |
| Species | LDA Score | p Value |
|---|---|---|
| Porphyromonas bennonis | 5.06 | 0.003 |
| Prevotella bivia | 5.6 | <0.001 |
| Prevotella disiens | 4.93 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campisciano, G.; Iebba, V.; Zito, G.; Luppi, S.; Martinelli, M.; Fischer, L.; De Seta, F.; Basile, G.; Ricci, G.; Comar, M. Lactobacillus iners and gasseri, Prevotella bivia and HPV Belong to the Microbiological Signature Negatively Affecting Human Reproduction. Microorganisms 2021, 9, 39. https://doi.org/10.3390/microorganisms9010039
Campisciano G, Iebba V, Zito G, Luppi S, Martinelli M, Fischer L, De Seta F, Basile G, Ricci G, Comar M. Lactobacillus iners and gasseri, Prevotella bivia and HPV Belong to the Microbiological Signature Negatively Affecting Human Reproduction. Microorganisms. 2021; 9(1):39. https://doi.org/10.3390/microorganisms9010039
Chicago/Turabian StyleCampisciano, Giuseppina, Valerio Iebba, Gabriella Zito, Stefania Luppi, Monica Martinelli, Leo Fischer, Francesco De Seta, Giuseppe Basile, Giuseppe Ricci, and Manola Comar. 2021. "Lactobacillus iners and gasseri, Prevotella bivia and HPV Belong to the Microbiological Signature Negatively Affecting Human Reproduction" Microorganisms 9, no. 1: 39. https://doi.org/10.3390/microorganisms9010039