Exploring the Capability of Yeasts Isolated from Colombian Fermented Cocoa Beans to Form and Degrade Biogenic Amines in a Lab-Scale Model System for Cocoa Fermentation

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Screening for Amino Acid Decarboxylation
2.3. Cocoa Samples
2.4. Lab-Scale Fermentation
2.5. pH Determination
2.6. Microbiological Analyses
2.7. Determination of Biogenic Amines
2.8. Organic Acids Extraction and Determination
2.9. Determination of Volatile Organic Compounds (VOCs)
2.10. Statistical Analyses
3. Results
3.1. Screening of Biogenic Amines (BAs) Production
3.2. Simulated Fermentation Using Cocoa Pulp Medium (SCPM)
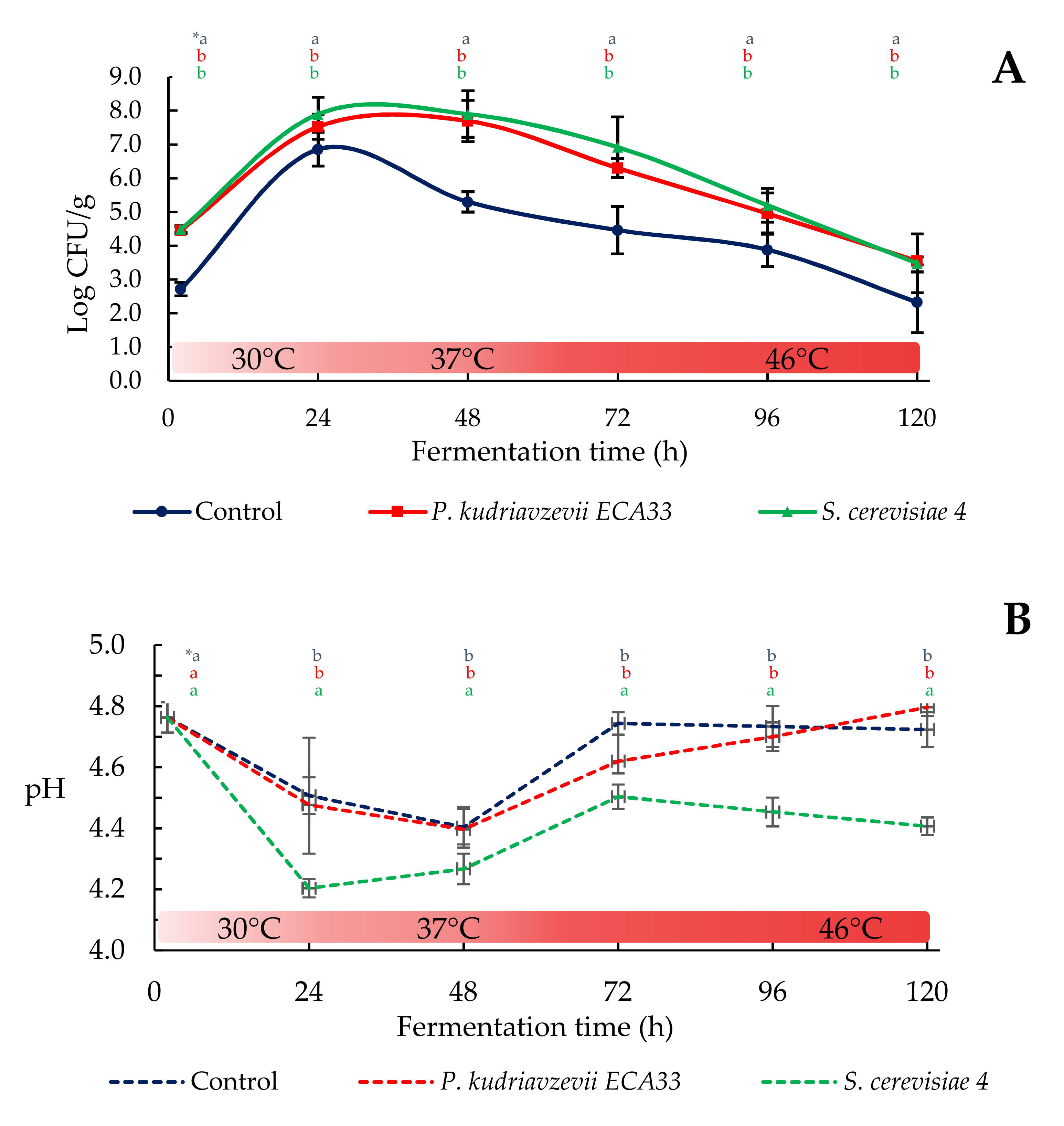
3.2.1. Microbial Growth and pH Dynamics
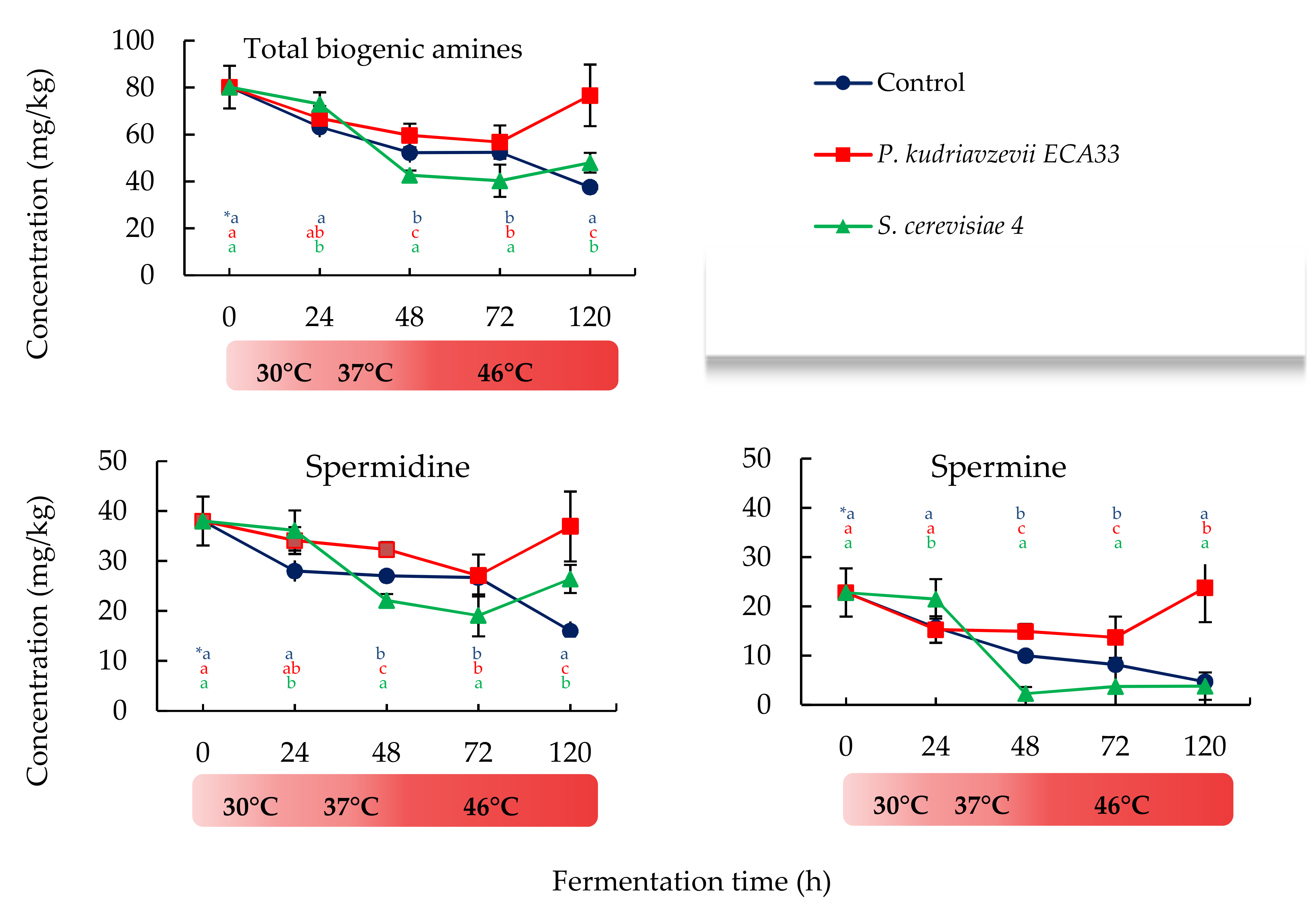
3.2.2. Changes in Biogenic Amines Content
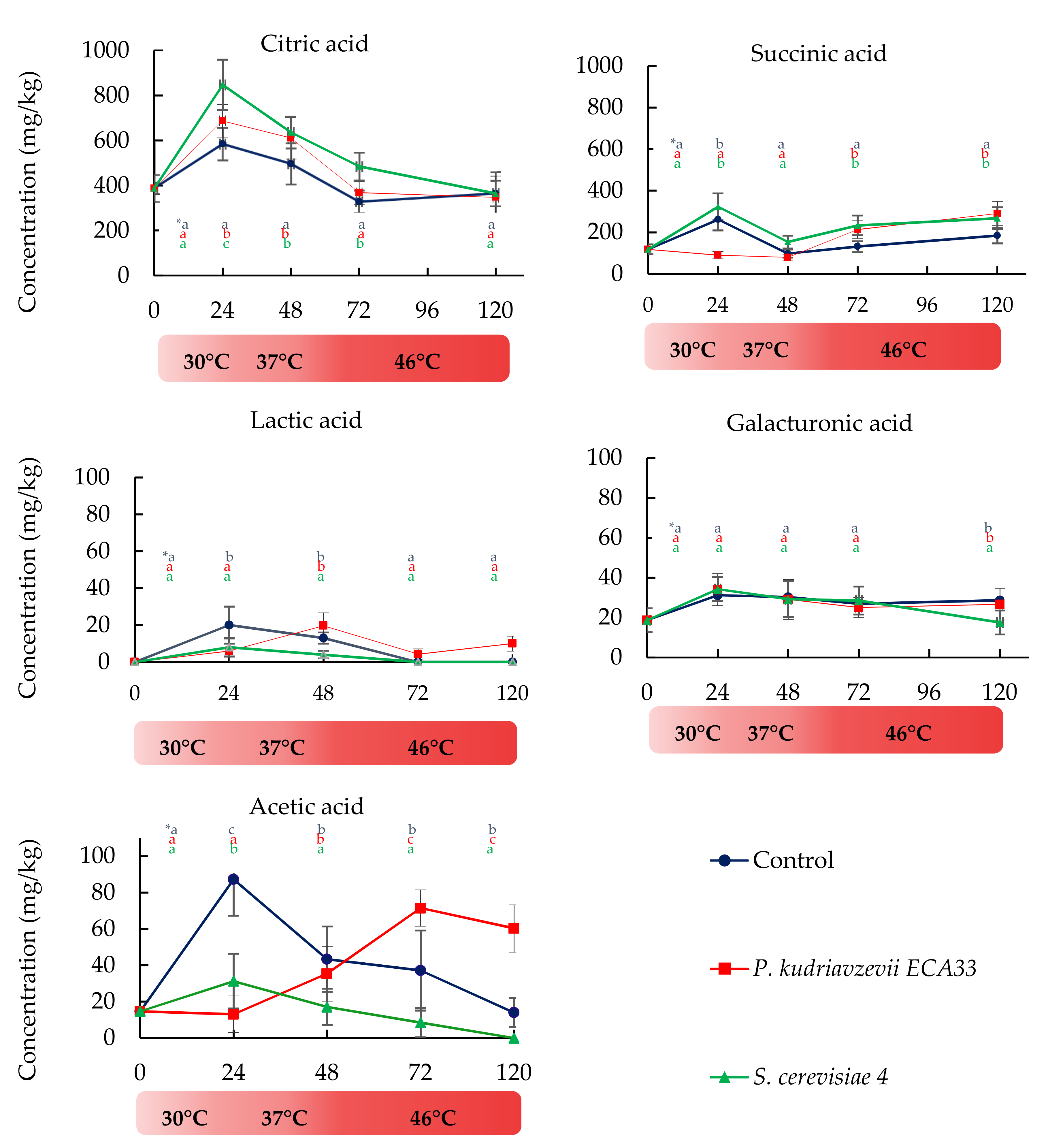
3.2.3. Organic Acids Dynamics
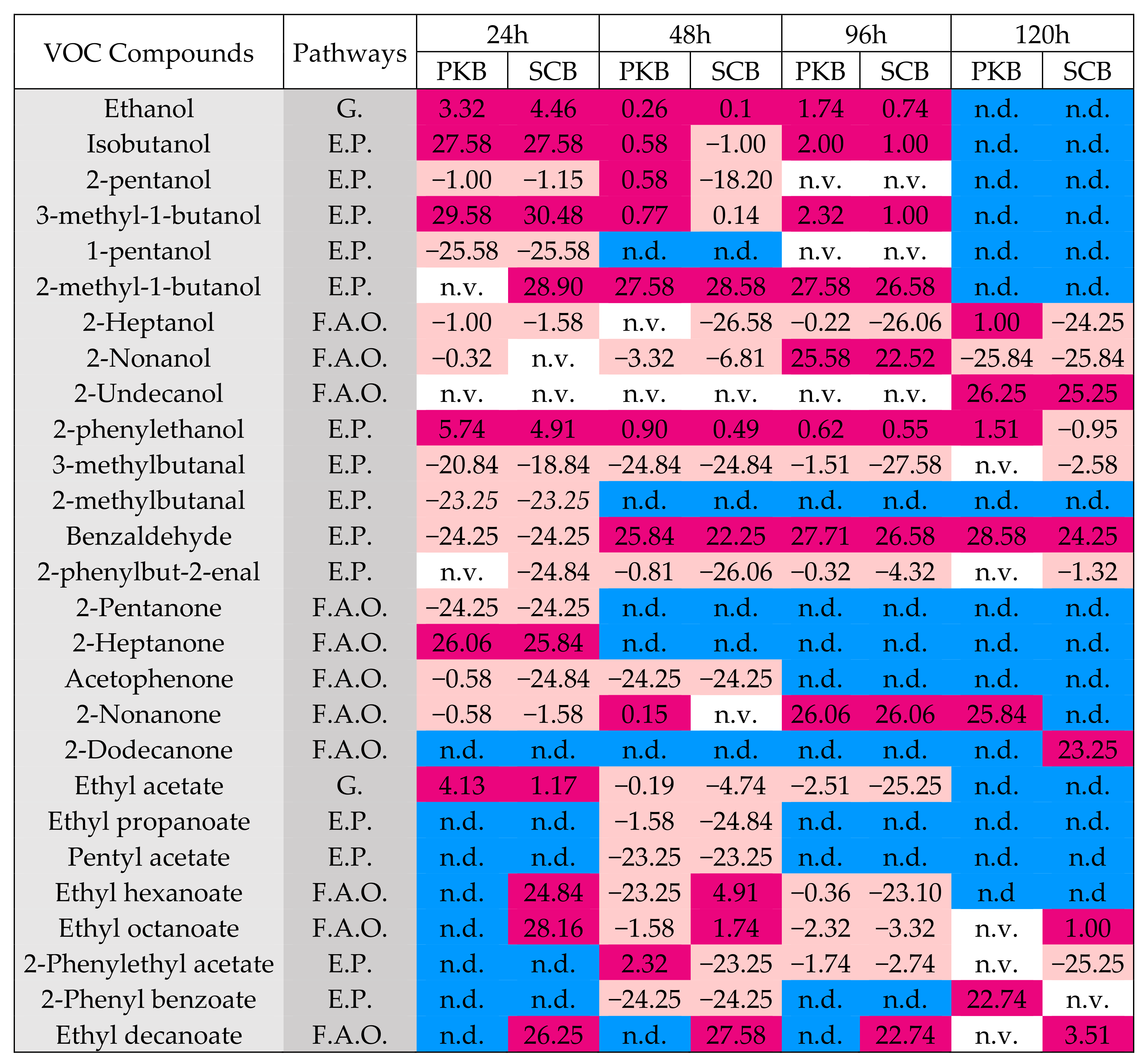
3.2.4. Volatile Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ardhana, M.M.; Fleet, G.H. The microbial ecology of cocoa bean fermentations in Indonesia. Int. J. Food Microbiol. 2003, 86, 87–99. [Google Scholar] [CrossRef]
- Ho, V.T.T.; Zhao, J.; Fleet, G. Yeasts are essential for cocoa bean fermentation. Int. J. Food Microbiol. 2014, 174, 72–87. [Google Scholar] [CrossRef]
- Koné, M.K.; Guéhi, S.T.; Durand, N.; Ban-Koffi, L.; Berthiot, L.; Tachon, A.F.; Brou, K.; Boulanger, R.; Montet, D. Contribution of predominant yeasts to the occurrence of aroma compounds during cocoa bean fermentation. Food Res. Int. 2016, 89, 910–917. [Google Scholar] [CrossRef]
- Spizzirri, U.G.; Ieri, F.; Campo, M.; Paolino, D.; Restuccia, D.; Romani, A. Biogenic amines, phenolic, and aroma-related compounds of unroasted and roasted cocoa beans with different origin. Foods 2019, 8, 306. [Google Scholar] [CrossRef] [Green Version]
- Leal Jr, G.A.; Gomes, L.H.; Efraim, P.; de Almeida Tavares, F.C.; Figueira, A. Fermentation of cacao (Theobroma cacao L.) seeds with a hybrid Kluyveromyces marxianus strain improved product quality attributes. FEMS Yeast Res. 2008, 8, 788–798. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Ospina, J.; Di Mattia, C.D.; Paparella, A.; Mastrocola, D.; Martuscelli, M.; Chaves-Lopez, C. Effect of Fermentation, Drying and Roasting on Biogenic Amines and Other Biocompounds in Colombian Criollo Cocoa Beans and Shells. Foods 2020, 9, 520. [Google Scholar] [CrossRef] [Green Version]
- Landete, J.M.; Ferrer, S.; Pardo, I. Biogenic amine production by lactic acid bacteria, acetic bacteria and yeast isolated from wine. Food Control. 2007, 18, 1569–1574. [Google Scholar] [CrossRef]
- ten Brink, B.; Damink, C.; Joosten, H.M.L.J.; Huis in’t Veld, J.H.J. Occurrence and formation of biologically active amines in foods. Int. J. Food Microbiol. 1990, 11, 73–84. [Google Scholar] [CrossRef]
- do Carmo Brito, B.D.N.; Campos Chisté, R.; da Silva Pena, R.; Abreu Gloria, M.B.; Santos Lopes, A. Bioactive amines and phenolic compounds in cocoa beans are affected by fermentation. Food Chem. 2017, 228, 484–490. [Google Scholar] [CrossRef]
- Granvogl, M.; Bugan, S.; Schieberle, P. Formation of amines and aldehydes from parent amino acids during thermal processing of cocoa and model systems: New insights into pathways of the strecker reaction. J. Agric. Food Chem. 2006, 54, 1730–1739. [Google Scholar] [CrossRef]
- Oracz, J.; Nebesny, E. Influence of roasting conditions on the biogenic amine content in cocoa beans of different Theobroma cacao cultivars. Food Res. Int. 2014, 55, 1–10. [Google Scholar] [CrossRef]
- Crafack, M.; Mikkelsen, M.B.; Saerens, S.; Knudsen, M.; Blennow, A.; Lowor, S.; Takrama, J.; Swiegers, J.H.; Petersen, G.B.; Heimdal, H.; et al. Influencing cocoa flavour using Pichia kluyveri and Kluyveromyces marxianus in a defined mixed starter culture for cocoa fermentation. Int. J. Food Microbiol. 2013, 167, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Mota-Gutierrez, J.; Botta, C.; Ferrocino, I.; Giordano, M.; Bertolino, M.; Dolci, P.; Cannoni, M.; Cocolin, L. Dynamics and biodiversity of bacterial and yeast communities during fermentation of cocoa beans. Appl. Environ. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares Menezes, A.G.; Batista, N.N.; Ramos, C.L.; Silva, A.R.d.A.e.; Efraim, P.; Pinheiro, A.C.M.; Schwan, R.F. Investigation of chocolate produced from four different Brazilian varieties of cocoa (Theobroma cacao L.) inoculated with Saccharomyces cerevisiae. Food Res. Int. 2016, 81, 83–90. [Google Scholar] [CrossRef]
- Meersman, E.; Steensels, J.; Mathawan, M.; Wittocx, P.-J.; Saels, V.; Struyf, N.; Bernaert, H.; Vrancken, G.; Verstrepen, K.J. Detailed analysis of the microbial population in Malaysian spontaneous cocoa pulp fermentations reveals a core and variable microbiota. PLoS ONE 2013, 8, e81559. [Google Scholar] [CrossRef] [Green Version]
- Ooi, T.S.; Ting, A.S.Y.; Siow, L.F. Influence of selected native yeast starter cultures on the antioxidant activities, fermentation index and total soluble solids of Malaysia cocoa beans: A simulation study. LWT 2020, 122, 108977. [Google Scholar] [CrossRef]
- Chaves-Lopez, C.; Serio, A.; Delgado-Ospina, J.; Rossi, C.; Grande-Tovar, C.D.; Paparella, A. Exploring the bacterial microbiota of colombian fermented maize dough “Masa agria” (Maiz añejo). Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Delgado-Ospina, J.; Triboletti, S.; Alessandria, V.; Serio, A.; Sergi, M.; Paparella, A.; Rantsiou, K.; Chaves-López, C. Functional Biodiversity of Yeasts Isolated from Colombian Fermented and Dry Cocoa Beans. Microorganisms 2020, 8, 1086. [Google Scholar] [CrossRef]
- Gardini, F.; Tofalo, R.; Belletti, N.; Iucci, L.; Suzzi, G.; Torriani, S.; Guerzoni, E.; Lanciotti, R. Characterization of Yeasts involved in the ripening of Pecorino Crotonese cheese. Food Microbiol. 2006, 23, 641–648. [Google Scholar] [CrossRef]
- Lee, A.H.; Neilson, A.P.; O’Keefe, S.F.; Ogejo, J.A.; Huang, H.; Ponder, M.; Chu, H.S.S.; Jin, Q.; Pilot, G.; Stewart, A.C. A laboratory-scale model cocoa fermentation using dried, unfermented beans and artificial pulp can simulate the microbial and chemical changes of on-farm cocoa fermentation. Eur. Food Res. Technol. 2019, 245, 511–519. [Google Scholar] [CrossRef]
- Sirocchi, V.; Caprioli, G.; Cecchini, C.; Coman, M.M.; Cresci, A.; Maggi, F.; Papa, F.; Ricciutelli, M.; Vittori, S.; Sagratini, G. Biogenic amines as freshness index of meat wrapped in a new active packaging system formulated with essential oils of Rosmarinus officinalis. Int. J. Food Sci. Nutr. 2013, 64, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Campos, J.; Escalona-Buendía, H.B.; Orozco-Avila, I.; Lugo-Cervantes, E.; Jaramillo-Flores, M.E. Dynamics of volatile and non-volatile compounds in cocoa (Theobroma cacao L.) during fermentation and drying processes using principal components analysis. Food Res. Int. 2011, 44, 250–258. [Google Scholar] [CrossRef]
- Ndagijimana, M.; Chaves-López, C.; Corsetti, A.; Tofalo, R.; Sergi, M.; Paparella, A.; Guerzoni, M.E.; Suzzi, G. Growth and metabolites production by Penicillium brevicompactum in yoghurt. Int. J. Food Microbiol. 2008, 127, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Delgado Ospina, J.; Grande Tovar, C.D.; Menjivar Flores, J.C.; Sánchez Orozco, M.S. Relationship between refractive index and thymol concentration in essential oils of Lippia origanoides Kunth. Chil. J. Agric. Anim. Sci. 2016, 32, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
- Pereira de Araújo, R.; Furtado de Almeida, A.-A.; Silva Pereira, L.; Mangabeira, P.A.O.; Olimpio Souza, J.; Pirovani, C.P.; Ahnert, D.; Baligar, V.C. Photosynthetic, antioxidative, molecular and ultrastructural responses of young cacao plants to Cd toxicity in the soil. Ecotoxicol. Environ. Saf. 2017, 144, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Liu, Y.; Sun, H.; Li, C.; Zhao, Z.; Liu, G. Advances in mechanisms and modifications for rendering yeast thermotolerance. J. Biosci. Bioeng. 2016, 121, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Figueroa-Hernández, C.; Mota-Gutierrez, J.; Ferrocino, I.; Hernández-Estrada, Z.J.; González-Ríos, O.; Cocolin, L.; Suárez-Quiroz, M.L. The challenges and perspectives of the selection of starter cultures for fermented cocoa beans. Int. J. Food Microbiol. 2019, 301, 41–50. [Google Scholar] [CrossRef]
- Ruiz-Capillas, C.; Herrero, A. Impact of biogenic amines on food quality and safety. Foods 2019, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Hue, C.; Gunata, Z.; Breysse, A.; Davrieux, F.; Boulanger, R.; Sauvage, F.X. Impact of fermentation on nitrogenous compounds of cocoa beans (Theobroma cacao L.) from various origins. Food Chem. 2016, 192, 958–964. [Google Scholar] [CrossRef]
- Dapkevicius, M.L.N.E.; Nout, M.J.R.; Rombouts, F.M.; Houben, J.H.; Wymenga, W. Biogenic amine formation and degradation by potential fish silage starter microorganisms. Int. J. Food Microbiol. 2000, 57, 107–114. [Google Scholar] [CrossRef]
- Visintin, S.; Ramos, L.; Batista, N.; Dolci, P.; Schwan, F.; Cocolin, L. Impact of Saccharomyces cerevisiae and Torulaspora delbrueckii starter cultures on cocoa beans fermentation. Int. J. Food Microbiol. 2017, 257, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Aprotosoaie, A.C.; Luca, S.V.; Miron, A. Flavor chemistry of cocoa and cocoa products-An overview. Compr. Rev. Food Sci. Food Saf. 2016, 15, 73–91. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Menichini, F.; Picci, N.; Puoci, F.; Spizzirri, U.G.; Restuccia, D. Technological aspects and analytical determination of biogenic amines in cheese. Trends Food Sci. Technol. 2013, 30, 38–55. [Google Scholar] [CrossRef]
- Costantini, A.; Vaudano, E.; Del Prete, V.; Danei, M.; Garcia-Moruno, E. Biogenic amine production by contaminating bacteria found in starter preparations used in winemaking. J. Agric. Food Chem. 2009, 57, 10664–10669. [Google Scholar] [CrossRef]
- Saez, J.S.; Lopes, C.A.; Kirs, V.E.; Sangorrín, M. Production of volatile phenols by Pichia manshurica and Pichia membranifaciens isolated from spoiled wines and cellar environment in Patagonia. Food Microbiol. 2011, 28, 503–509. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and safety aspects of yeast strains characterized from vineyards and spontaneous fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef]
- Simon Amoikon, T.L.; Marcotte, S.; N’Dede Djeni, T.; Celaire N’Sa, K.M.; Grondin, C.; Tinsley, C.; Casaregola, S.; Dje, M.K. A study on the potential of yeasts isolated from palm wines to produce flavouring compounds. LWT 2020, 128, 109506. [Google Scholar] [CrossRef]
- Michael, A.J. Polyamines in Eukaryotes, Bacteria, and Archaea. J. Biol. Chem. 2016, 291, 14896–14903. [Google Scholar] [CrossRef] [Green Version]
- Mah, J.-H.; Park, Y.; Jin, Y.; Lee, J.-H.; Hwang, H.-J. Bacterial production and control of biogenic amines in Asian fermented soybean foods. Foods 2019, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, B.; Farshidi, M. Innovative approaches for the degradation of biogenic amines in foods. Curr. Nutr. Food Sci. 2018, 14, 1–2. [Google Scholar] [CrossRef]
- Gardini, F.; Martuscelli, M.; Crudele, M.A.; Paparella, A.; Suzzi, G. Use of Staphylococcus xylosus as a starter culture in dried sausages: Effect on the biogenic amine content. Meat Sci. 2002, 61, 275–283. [Google Scholar] [CrossRef]
- Kalač, P. Health effects and occurrence of dietary polyamines: A review for the period 2005–mid 2013. Food Chem. 2014, 161, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, D.; Large, P.J. Enzymes metabolizing dimethylamine, trimethylamine and trimethylamine N-oxide in the yeast Sporopachydermia cereana grown on amines as sole nitrogen source. FEMS Microbiol. Lett. 1986, 35, 99–105. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific Opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef] [Green Version]
- Papalexandratou, Z.; Kaasik, K.; Kauffmann, L.V.; Skorstengaard, A.; Bouillon, G.; Espensen, J.L.; Hansen, L.H.; Jakobsen, R.R.; Blennow, A.; Krych, L.; et al. Linking cocoa varietals and microbial diversity of Nicaraguan fine cocoa bean fermentations and their impact on final cocoa quality appreciation. Int. J. Food Microbiol. 2019, 304, 106–118. [Google Scholar] [CrossRef]
- Jespersen, L.; Nielsen, D.S.; Hønholt, S.; Jakobsen, M. Occurrence and diversity of yeasts involved in fermentation of West African cocoa beans. FEMS Yeast Res. 2005, 5, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Illeghems, K.; Weckx, S.; De Vuyst, L. Applying meta-pathway analyses through metagenomics to identify the functional properties of the major bacterial communities of a single spontaneous cocoa bean fermentation process sample. Food Microbiol. 2015, 50, 54–63. [Google Scholar] [CrossRef]
- Ouattara, D.H.; Ouattara, H.G.; Droux, M.; Niamke, S.L. Analysis of environmental conditions affecting acids production in lactic acid bacteria involved in Ivorian cocoa fermentation. Sch. Acad. J. Biosci. 2019, 7, 158–165. [Google Scholar] [CrossRef]
- Moens, F.; Lefeber, T.; De Vuyst, L. Oxidation of metabolites highlights the microbial interactions and role of Acetobacter pasteurianus during cocoa bean fermentation. Appl. Environ. Microbiol. 2014, 80, 1848. [Google Scholar] [CrossRef] [Green Version]
- Mansour, S.; Beckerich, J.M.; Bonnarme, P. Lactate and amino acid catabolism in the cheese-ripening yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 6505–6512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, Y.C.; Shaffer, C.E.H.; Bennett, G.N. Microbial formation of esters. Appl. Microbiol. Biotechnol. 2009, 85, 13–25. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° of Total Strains | Tyr | Phe | Orn | His | Lys | |
|---|---|---|---|---|---|---|
| Candida parapsilosis | 10 | 50 | 10 | 60 | 0 | 0 |
| Hydropichia burtonii | 8 | 37.5 | 0 | 62.5 | 0 | 12.5 |
| Pichia kudriavzevii | 54 | 12.96 | 15.42 | 90.74 | 2.20 | 90.74 |
| Pichia manshurica | 13 | 0 | 0 | 23.07 | 0 | 0 |
| Saccharomyces cerevisiae | 38 | 23.68 | 5.26 | 28.94 | 0 | 23.68 |
| Schizosaccharomyces pombe | 4 | 25 | 0 | 25 | 0 | 0 |
| Starmerella sorbosivorans | 2 | 0 | 0 | 0 | 0 | 0 |
| Torulaspora delbruekii | 2 | 0 | 0 | 0 | 0 | 0 |
| Torulaspora pretoriensis | 2 | 0 | 0 | 0 | 0 | 0 |
| Trichosporon asahii var. asahii | 3 | 100 | 23.07 | 23.07 | 0 | 15.38 |
| Wickerharmomyces anomalus | 6 | 100 | 16.66 | 83.33 | 0 | 0 |
| Zygosaccharomyces bisporus | 3 | 0 | 33.33 | 0 | 0 | 0 |
| Biogenic Amines Content (mg/kg) | |||||
|---|---|---|---|---|---|
| Fermentation Time (h) | 0 | 24 | 48 | 96 | 120 |
| Control | |||||
| TRY | 4.8 ± 0.9 cA | 2.6 ± 0.4 bA | 0.9 ± 0.1 aA | 3.8 ± 0.5 bcA | 2.5 ± 0.3 bA |
| PHE | 2.6 ± 0.5 aA | 6.3 ± 0.5 bA | 6.7 ± 1.9 bA | 6.1 ± 1.3 bA | 6.2 ± 0.3 bA |
| PUT | 9.0 ± 0.6 cA | 6.3 ± 0.7 bA | 3.8 ± 0.6 aA | 3.7 ± 0.0 aA | 4.7 ± 0.6 abA |
| CAD | 0.8 ± 0.1 aA | 3.4 ± 0.5 bA | 2.9 ± 0.2 bA | 3.3 ± 0.3 bA | 3.1 ± 0.8 bA |
| TYR | 2.2 ± 0.1 cA | 1.1 ± 0.2 bA | 1.0 ± 0.1 bA | 0.6 ± 0.1 aA | 0.4 ± 0.0 aA |
| P kudriavzevii ECA33 | |||||
| TRY | 4.8 ± 0.9 cA | 4.6 ± 0.1 bcB | 3.6 ± 0.1 abB | 4.3 ± 0.1 bcA | 4.6 ± 0.5 bcB |
| PHE | 2.6 ± 0.5 aA | 2.4 ± 0.2 aB | 1.9 ± 0.0 aB | 3.8 ± 0.1 bB | 3.9 ± 0.8 bB |
| PUT | 9.0 ± 0.6 bA | 8.6 ± 0.5 bB | 5.5 ± 1.1 aB | 5.8 ± 1.2 aB | 5.7 ± 0.3 aA |
| CAD | 0.8 ± 0.1 aA | 0.6 ± 0.1 aB | 2.3 ± 0.4 cA | 1.5 ± 0.3 bC | 1.5 ± 0.2 bB |
| TYR | 2.2 ± 0.1 dA | 1.4 ± 0.0 cA | 1.0 ± 0.2 bA | 0.6 ± 0.1 aA | 0.4 ± 0.1 aA |
| S. cerevisiae 4 | |||||
| TRY | 4.8 ± 0.9 abA | 1.7 ± 0.1 aA | 5.8 ± 1.1 bC | 6.2 ± 1.0 bB | 7.2 ± 1.9 bC |
| PHE | 2.6 ± 0.5 bcA | 1.6 ± 0.1 abC | 3.2 ± 0.8 cC | 2.4 ± 0.2 bcC | 1.0 ± 0.1 aC |
| PUT | 9.0 ± 0.6 cA | 7.8 ± 1.6 cB | 7.7 ± 0.7 cA | 6.8 ± 1.2 aB | 7.4 ± 0.0 bC |
| CAD | 0.8 ± 0.1 aA | 2.8 ± 0.4 bA | 0.8 ± 0.2 aC | 1.0 ± 0.1 aC | 1.3 ± 0.1 aB |
| TYR | 2.2 ± 0.1 cA | 1.3 ± 0.2 bA | 0.8 ± 0.05 aA | 1.1 ± 0.1 abC | 0.8 ± 0.1 aB |
| Variable | TRY | PHE | PUT | CAD | TYR | Total BA |
|---|---|---|---|---|---|---|
| Control | 0.70 | −0.56 | 0.78 | −0.35 | 0.55 | 1.00 |
| P. kudriavzevii ECA33 | 0.90 | 0.21 | 0.82 | −0.93 | 0.60 | 1.00 |
| S. cerevisiae 4 | 0.63 | 0.51 | 0.43 | −0.93 | 0.35 | 1.00 |
| Temperature | 0.15 | 0.23 | −0.68 | 0.26 | −0.89 | −0.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado-Ospina, J.; Acquaticci, L.; Molina-Hernandez, J.B.; Rantsiou, K.; Martuscelli, M.; Kamgang-Nzekoue, A.F.; Vittori, S.; Paparella, A.; Chaves-López, C. Exploring the Capability of Yeasts Isolated from Colombian Fermented Cocoa Beans to Form and Degrade Biogenic Amines in a Lab-Scale Model System for Cocoa Fermentation. Microorganisms 2021, 9, 28. https://doi.org/10.3390/microorganisms9010028
Delgado-Ospina J, Acquaticci L, Molina-Hernandez JB, Rantsiou K, Martuscelli M, Kamgang-Nzekoue AF, Vittori S, Paparella A, Chaves-López C. Exploring the Capability of Yeasts Isolated from Colombian Fermented Cocoa Beans to Form and Degrade Biogenic Amines in a Lab-Scale Model System for Cocoa Fermentation. Microorganisms. 2021; 9(1):28. https://doi.org/10.3390/microorganisms9010028
Chicago/Turabian StyleDelgado-Ospina, Johannes, Laura Acquaticci, Junior Bernardo Molina-Hernandez, Kalliopi Rantsiou, Maria Martuscelli, Astride Franks Kamgang-Nzekoue, Sauro Vittori, Antonello Paparella, and Clemencia Chaves-López. 2021. "Exploring the Capability of Yeasts Isolated from Colombian Fermented Cocoa Beans to Form and Degrade Biogenic Amines in a Lab-Scale Model System for Cocoa Fermentation" Microorganisms 9, no. 1: 28. https://doi.org/10.3390/microorganisms9010028