Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications


1
Department of Microbiology, North West University Mafikeng Campus, Private Bag X2046, Mmabatho 2735, South Africa
2
Food Security and Safety Niche Area, Faculty of Agriculture, Science and Technology, North West University, Mmabatho, Mafikeng 2735, South Africa
*
Author to whom correspondence should be addressed.
Microorganisms 2020, 8(12), 1934; https://doi.org/10.3390/microorganisms8121934
Submission received: 2 November 2020
/
Revised: 20 November 2020
/
Accepted: 26 November 2020
/
Published: 6 December 2020
(This article belongs to the Special Issue Natural Antimicrobial Compounds)
Abstract
:Over the last century, endophytic fungi have gained tremendous attention due to their ability to produce novel bioactive compounds exhibiting varied biological properties and are, therefore, utilized for medicinal, pharmaceutical, and agricultural applications. Endophytic fungi reside within the plant tissues without showing any disease symptoms, thus supporting the physiological and ecological attributes of the host plant. Ground breaking lead compounds, such as paclitaxel and penicillin, produced by endophytic fungi have paved the way for exploring novel bioactive compounds for commercial usage. Despite this, limited research has been conducted in this valuable and unique niche area. These bioactive compounds belong to various structural groups, including alkaloids, peptides, steroids, terpenoids, phenols, quinones, phenols, and flavonoids. The current review focuses on the significance of endophytic fungi in producing novel bioactive compounds possessing a variety of biological properties that include antibacterial, antiviral, antifungal, antiprotozoal, antiparasitic, antioxidant, immunosuppressant, and anticancer functions. Taking into consideration the portal of this publication, special emphasis is placed on the antimicrobial and antiviral activities of metabolites produced by endophytes against human pathogens. It also highlights the importance of utilization of these compounds as potential treatment agents for serious life-threatening infectious diseases. This is supported by the fact that several findings have indicated that these bioactive compounds may significantly contribute towards the fight against resistant human and plant pathogens, thus motivating the need enhance the search for new, more efficacious and cost-effective antimicrobial drugs.
1. Introduction
In recent years, there has been a dramatic shift towards a more sustainable, eco-friendly, and natural way of living. Many researchers are of the opinion that there is an alarming increase in drug resistance worldwide and the problem is escalating tremendously, thus rendering the current antimicrobial agents ineffective [1,2,3]. Approximately two million people worldwide are infected with antibiotic-resistant pathogens, resulting in at least 23,000 mortalities per annum [4]. According to the World Health Organization (WHO), antimicrobial resistance (AMR) has emerged as one of the most serious public health concerns of the 21st century [5]. Moreover, due to the many disadvantages and side effects associated with current antimicrobial agents, it is not surprising that a large proportion of individuals, especially those who live in developing countries, are utilizing naturally available bioactive alternatives for their primary healthcare.
In developing countries, especially those in Africa, Asia, and Latin America, more than 80% of the population utilize medicinal plants to address their primary healthcare needs and wellness [6]. With over 400,000 diverse plant species inhabiting the planet, a majority are capable of treating a broad spectrum of ailments. Due to the fact that they possess a wide variety of biological properties, this has increased interest in research designed to search for “ideal” bioactive compounds that may be of benefit to mankind [7,8,9]. However, over-propagation and excessive usage might lead to endangerment and extinction of the plants. In-depth studies have shown that endophytic fungi are capable of colonizing plant tissues, providing protection, and are a rich source of natural bioactive compounds [10].
In addition, the ratio of fungal species to vascular plants is approximately 1:6, which is estimated at 1.5 million fungal species [11]. Endophytes are microorganisms that colonize internal plant tissues without causing any apparent harm to the host plant throughout their life cycle. This interaction is considered as mutualistic symbiosis, whereby both partners benefit from the association [12,13,14]. The endophytes aid in the physiological and ecological roles of the plants, resulting in protection and survival mechanisms. Moreover, endophytic fungi promote growth, prevent abiotic and biotic stresses, such as drought conditions, salinity, extreme temperatures, heavy metal toxicity, and oxidative stress, and provide protection from insect and herbivores [13,15,16]. Endophytes have the ability to prevent resistance mechanisms by overcoming pathogenic invasion using secondary metabolites.
Bioactive compounds are primarily responsible for the beneficial characteristics displayed by endophytic fungi. History has shown that such metabolic compounds might aid in ground-breaking discoveries, e.g., the discovery of penicillin produced by Penicillium chrysogenum, which was a milestone in the development of antibiotic drugs, as a “front-line” antibiotic which saved millions of lives; hence, it was referred to as “wonder drug” [17]. Another “gold” bioactive compound is paclitaxel (taxol), which is produced by Taxomyces andreanae for chemotherapy purposes [18]. These bioactive metabolites might be structurally classified into alkaloids, benzopyranones, chinones, peptides, phenols, quinones, flavonoids, steroids, terpenoids, tetralones, xanthones, and others [19]. They have exhibited numerous biological properties, including antibacterial, antifungal, immunosuppressants, antiviral, antiparasitic, antioxidant, anti-inflammatory, and anticancer properties [20].
New biotechnological advances concentrate on the search for and utilization of novel bioactive compounds extracted from endophytic fungi. Despite all of this, only a tiny portion of endophytic fungi have been isolated and investigated for their biological activities. In this current review, we focus in depth on the various biological properties demonstrated by endophytic fungi. We also identify novel bioactive compounds associated with the activities. Furthermore, we shed light on the activities against resistant pathogenic microorganisms. Unexplored niche areas such as these will progress the drug development process tremendously. Endophytic fungi are an abundant source of natural bioactive compounds which are novel, renewable, and low in toxicity and more efficacious, more potent, more affordable, safer, and less resistant than conventional antimicrobial agents. Hence, this will alleviate the massive burden on public healthcare systems and aid in the medical and pharmaceutical industries.
2. Biological Properties of Novel Bioactive Compounds from Endophytic Fungi
Various biological activities, such as antibacterial, antifungal, immunosuppressant, antiviral, antiparasitic, antioxidant, anti-inflammatory, and anticancer, were exhibited by endophytic fungi with a wealth of bioactive metabolites. In this section, we investigate endophytic fungi as a source of countless bioactive compounds which might be beneficial for human health.
2.1. Bioactive Compounds as an Alternative Antibacterial Agents
Since the golden era of penicillin, antibiotics have become a staple conventional medicine and have reduced the fatality rate by saving millions of lives, as was proven in World War II [21]. In the 1940s, it was the first prescribed antibiotic utilized in the treatment of life-threatening infections. Shortly afterward, the development of antibiotic resistance, and particularly the detection of multiple antibiotic-resistant (MAR) bacteria, has resulted to downturn reduction of the activity of antibiotics [22]. The dramatic increase in microbial resistance reduces the efficacy of existing antibiotics and, therefore, negatively affects their applications in human medicine. Over the last century, a number of antibiotics have emerged in the market, and despite this developmental processes, there is evidence of antibiotic resistance being developed against all these antibiotics (Figure 1) [23]. According to the World health Organization (WHO), antibiotic resistance remains one of the leading threats to human health and poses a severe financial burden on healthcare systems worldwide [24].
Aharwal and co-workers [25], recognized the severity of antibiotic resistance and emphasized the importance of innovative research and development strategies. Several studies have established that endophytes have resistance mechanisms in place to overcome pathogenic invasion through the production of secondary compounds [15,19,26]. Bioactive metabolites are low-molecular-weight, organic natural substances produced by microorganisms that possess activities at low concentrations against other microorganisms, in respect to bioactive compounds with potential antibiotic properties. Antibacterial compounds produced by endophytes have been shown to occupy a broad spectrum of structural classes, such as alkaloids, peptides, steroids, terpenoids, phenols, quinines, and flavonoids [27].
Recently, numerous scientists have demonstrated that endophytic fungi have a variety of beneficial, novel, and effective bioactive metabolites possessing antibacterial activity. Endophytic Alternaria alternate (AE1) was isolated from healthy and mature leaves of Azadirachta indica plants located in Santiniketan. The fungal extracts exhibited excellent antimicrobial activities against both Gram-positive and Gram-negative bacteria (Bacillus subtilis MTCC 121, Listeria monocytogenes MTCC 657, Staphylococcus aureus MTCC 96 and Staphylococcus epidermidis MTCC 2639, Salmonella typhimurium MTCC 98, Pseudomonas aeruginosa MTCC 741, and Escherichia coli MTCC 1667) with minimum inhibitory concentration (MIC) values of 300–400 μg/mL. The mode of action was determined as being cidal and further analysis demonstrated that cell lysis or leakage of cell membrane was occurring. Gas chromatography–mass spectrometry (GC-MS) analysis confirmed that the fungal extracts consist of several bioactive compounds [28]. The bioactive compounds produced from endophytic fungal extracts of Penicillium sp. demonstrated strong effectiveness against Enterococcus faecalis with an MIC value of 62.50 mg/mL [29]. Linoleic acid (9,12-octadecadienoic acid (Z,Z)) and cyclodecasiloxane produced by endophytic Alternaria sp. were successfully isolated from native South African Pelargonium sidoides, and this is considered as a first report. These fungal metabolites inhibited several food-borne and food spoilage bacteria, including Bacillus cereus, Escherichia coli, and Enterococcus faecium, and E. gallinarum showed a 2–12-mm zone of inhibition [30]. Despite the fact that root-knot nematodes such as Meloidogyne incognita are persistent parasites in plants and contribute to about 5% of global crop losses [31], a biocontrol investigation revealed that the endophytic fungus Piriformospora indica was capable of inhibiting the root-knot nematode parasite and, thus, enhanced plant growth [31].
Kjer et al. (2009) discovered two new secondary metabolites (10-oxo-10H-phenaleno [1,2,3-de] chromene-2-carboxylic acids and xanalteric acids I and II, Figure 2) which were produced by the fungus Alternaria sp., isolated from the mangrove (Sonneratia albacollected) plant located in China. The metabolites exhibited strong antibacterial properties against Enterococcus faecalis, Pseudomonas aeroginosa, and Staphylococcus epidermidis [32].
Ding and colleagues (2019) also reported compounds 1 and 2 as novel isocoumarin derivatives (Figure 2) with a distinctive butanetriol group at C-3 using NMR and MS. Furthermore, the bioactive compounds exhibited some activity against Gram-negative bacteria. Compounds 1 and 2 showed effectiveness against E. coli, with MIC values of 32 μg/mL [34]. Besides that, other studies also discovered novel ester metabolites isolated from endophytic fungus from the eastern larch that possess antibacterial efficacy against Vibrio salmonicida, Pseudomonas aeruginosa, and Staphylococcus aureus. These compounds were identified as 8,1′,5′-trihydroxy-3′,4′dihydro-1′H-[2,4′]binaphthalenyl-1,4,2′-trione, (1), and 2-methyloctanoic acid 6-oxo-2-propenyl-3,6-dihydro-2H-pyran-3-yl ester, (2) [35]. Figure 2 represent some bioactive compounds are with antibacterial effect.
2.2. Bioactive Compounds as an Alternative to Antifungal Agents
The diversity of fungal species is currently estimated at 2.2 to 3.8 million worldwide, with soil fungal populations contributing a significant portion, and fungi are regarded as ubiquitous [36]. This suggests that there might be more fungal interaction, whether beneficial or pathogenic. Recently, fungal diseases were accountable for over a 1.6 million mortality rate per annum, with over one billion individuals in a state of severe morbidity. In the agricultural sector, fungal pathogens might lead to damage or loss of crops, economic losses, and, eventually, affect food security and food production [37]. In addition, there has been a dramatic increase in the resistance of microorganisms to current antimicrobial agents, including antifungal agents. Despite the inefficacy of antifungal agents, there is currently a lack of therapeutic agents on the market. Fungal diseases are generally underrated and overlooked, although they cause secondary infections to hospitalized patients. Pathogenic fungi are opportunistic and affect immunocompromised individuals, thus posing a serious burden on current health care systems worldwide [38].
Research advances in modern antifungal agents rely highly on screening for novel bioactive compounds that have gained popularity in the drug development process and will aid in the fight against animal and human fungal pathogens. Peláez and co-workers (2000) established that new triterpene glycoside exhibited good antifungal inhibitory activities against Candida and Aspergillus sp. with 19 and 30 mm, respectively. Furthermore, in vivo studies conducted on mouse models showed moderate efficacy against candidiasis [39]. Recent studies revisited formerly undescribed bioactive compounds—strobilurin G, favolon, pterulinic acid, and 2,3-dihydro-1-benzoxepin derivative isolated from Favolaschia calocera—which displayed antifungal effectiveness. The minimum inhibitory concentration (MIC) displayed by compound 1 was ≤ 9.37 and ≤ 18.75 μg/mL against Candida tenuis and Mucor plumbeus, respectively [40]. Similar studies showed that endophytic fungi (Penicillium sp. (C7B) and Trichoderma sp. (B1C, C4E, C4D)) were effective against Escherichia coli, Staphylococcus aereus, and Vibrio alginolyticus, displaying clear-zone diameters of 17.91 ± 0.84 mm; 17.78 ± 0.83 mm; 17.66 ± 0.83 mm; 16.72 ± 1.15 mm, and 13.65 ± 0.27 mm, respectively [41].
“Pestalachlorides A” (C21H21Cl2NO5) and “B” (C20H18Cl2O5) are chlorinated benzophenone derivatives that inhibited plant pathogenic fungi, Fusarium culmorum, Gibberella zeae, and Verticillium albo-atrum, exhibiting MIC activities of 7.2, 144.4, and 114.4 Μm respectively [42]. Well known compounds also include iridoide, glcopyranoside and saponin (Figure 3).
Endophytic Humicola sp. (JS-0112 strain) proved to be an ideal candidate for the development of novel fungicides by controlling plant pathogenic Sclerotinia homoeocarpa [43]. Current studies have discovered two novel polyketides with an unprecedented “C12-C6” carbon skeleton isolated from endophytic Phomopsis sp. (CFS42). Moreover, the compounds inhibited activities against Bipolaris sorokinian, Alternaria alternat, Fursarium avenaceum, and Curvularia lunata. Therefore, this reiterates the untapped wealth of novel bioactive compounds produced by endophytic fungi [44].
2.3. Bioactive Compounds for Treating Cancer Cells (Anticancer Activity)
According to the World Health Organization (WHO), cancer is a group of diseases caused by the malignant growth of cells or tumor resulting from uncontrolled cell division [45]. Cancer was primarily responsible for one in six deaths in among humans in annum of 2018, resulting in an overall estimate of 9.6 million deaths worldwide. Cancer was reported to be the second leading cause of deaths worldwide. Men are often more susceptible to lung, prostate, colorectal, stomach, and liver cancer, while women often suffer from breast, colorectal, lung, cervical, and thyroid cancer [45]. The majority (70%) of deaths caused by cancer occur in low- and middle-income countries in Africa, Asia, and Central and South America [45]. Currently, therapeutic processes for treating cancer patients present tremendous challenges to both the physician and the patient, resulting from the lack of precision as well as lack of bioavailability, since most anticancer agents are naturally lipophilic and have a high first-pass effect. In addition, these agents are also non-specific towards their targets and, therefore, may interact with non-cancerous tissues. Moreover, cancer patients most often experience adverse effects after therapeutic procedures that are also associated with high toxicity [46,47,48]. Due to these complications, cancers and their associated diseases are considered to be a serious healthcare burden to patients globally [49]. This explains the reasons for continuous focused investigations aimed at discovering new natural bioactive compounds, especially from endophytes, that may serve as alternative agents to combat cancer [47,48].
The ground-breaking study resulting in the discovery of paclitaxel, also known as the “golden” compound, produced renewed hope in the search for novel anticancer agents, and the compound gained significant popularity because of its distinctive mode of action when compared to other anticancer agents. Paclitaxel obstructs the uncontrolled duplication of cancerous cells, thereby reducing their growth and spread. Paclitaxel with a chemical formula of C47H51NO14 is formulated from ‘taxol”, the first anticancer drug to generate billions of dollars. The endophytic fungus (Taxomyces andreanae) isolated from the Pacific yew bark (Taxus brevifolia) produces the anticancer bioactive compound paclitaxel. A number of studies have also isolated taxol and its related compounds from a variety of endophytes, including Grammothele lineata [50], Aspegillus aculeatinus [51], Alternaria brassicicola [52], and Acremonium, Colletotrichum, and Fusarium spp., from an ecologically altered Taxus baccata [53]. Taxol and its associated compounds were characterized and confirmed using UV absorption, HPLC, FTIR spectra, and LC–ESI–MS [50,51,52,53]. Despite its very effective anticancer properties, taxol is rare, which amplifies the need to enhance the search for alternative sources to this very important bioactive metabolite [51]. In addition, plants that produce taxol appear to be “rare”, which, coupled with the fact that the production of taxol from Taxus plants is a complex process, means that an alternative will be to constantly assess taxol-producing endophytic fungi due to the ease and practicality of obtaining these valuable compounds. In order to understand the complexity of the processes or molecular mechanisms involved in the production of taxol, [51] assessed the transcriptome of Aspergillus aculeatinus Tax-6, indicating that genes involved in two pathways (mevalonate (MVA) and nonmevalonate (MEP)) need to be expressed. Despite the fact that the potential of endophytes to produce taxol varies, with Grammothele lineata producing 382.2 μg/L [50], Alternaria brassicicola producing 140.8 μg/L taxol [52], and Acremonium, Colletotrichum, and Fusarium spp., which also produced up to 116.19 μg/L [53], the production of a very high yield (560 µg/L) by the mutant strain A. aculeatinus BT-2 when compared to the wild-type A. aculeatinus Tax-6 strain [51] indicates the importance of genetic manipulation in the search for these very important bioactive compounds. In addition to the previously mentioned studies, other endophytic fungi, such as Pestalotiopsis microspore, Alternaria alternata, Periconia sp., Pithomyces sp., Chaetomella raphigera, Monochaetia sp., Seimatoantlerium nepalense, Botryodiplodia theobromae, Phyllosticta spinarum, Pestalotiopsis terminaliae, and Bartalinia robillardoides, have also been found to produce anticancer bioactive agents that chemically belong to taxols [48,54]. Several taxol-producing endophytes therefore provided a cheaper and more accessible alternative for anticancer agents [54], and it is on this basis that taxol compounds were approved by the Food and Drug Administration (FDA) for the treatment of advanced breast cancer, lung cancer, and refractory ovarian cancer [55].
In addition to paclitaxel, camptothecin (C20H16N2O4), a bioactive alkaloid compound initially isolated from Camptotheca acuminata wood in China, also exhibited anticancer properties [56,57]. Furthermore, another anticancer compound, “chaetoglobosin U”, was produced by the fungus Chaetomium globosum, which lives synergistically in the stem of healthy Imperata cylindrica [58]. Other novel bioactive compounds, such as cytoglobosins C17 and D18 alkaloids from the endophytic fungus Chaetomium globosum, inhibited antitumor activities on the cancer cell line A549 [59]. Several bioactive compounds have been shown to possess anticancer activities (Figure 4). Li et al. (2013) [58] also established that the endophyte Chaetomium globosum from the Ginkgo biloba plant contained three novel compounds that include azaphilone alkaloids, chaetomugilides A–C, and chaetoviridin E, all of which displayed high cytotoxic activities against the human cancer cell line HePG2 [58]. Other endophytic fungi belonging to the genera Xylaria, Phoma, Hypoxylon, and Chalara have also been reported to produce cytochalasins, which possess antitumor activities. In addition, three novel cytochalasins, namely cytochalasin H, cytochalasin J, and cytochalasin E, were also extracted from Rhinocladiella sp. isolated from Tripterygium wilfordii [60]. Podophyllotoxin (C22H22O8), a lignan-type bioactive compound which was originally isolated from Podophyllum peltatum L. in the 1880s, and its derivatives are well-known to possess a variety of biological properties and are used as cathartic, purgative, antiviral, vesicant, antibacterial, antihelminthic, and antitumor agents. Etoposide and teniposide are chemotherapeutic medications produced from podophyllotoxin, and they are currently available in the market. In addition, endophytes Fusarium oxysporum, Aspergillus fumigatus, Phialocephala fortinii, and Trametes hirsute, as well as some belonging to the genera Trichoderma, Penicillium, and Phomopsis, have also been reported to produce the compound podophyllotoxin, which exhibits anticancer activities [61,62].
The discovery of other lead compounds was intensified by the isolation of camptothecin from the endophytic fungi Fusarium solani housed in Camptotheca acuminata Decaisne (Nyssaceae) wood located in China. Camptothecin (C20H16N2O4) is an alkaloid topoisomerase compound that displayed remarkable potent antineoplastic efficacy. Camptothecin and 10-hydroxycamptothecin were utilized in the development of the drugs topotecan and irinotecan, which are chemotherapeutic. Since then, precursor 9-methoxycamptothecin and 10-hydroxycamptothecin have been reported to exhibit powerful anticancer properties [63,64]. In addition to these findings, some reports have revealed that camptothecin and methoxy camptothecin are capable of enhancing the development of fruits and seed germination phases, thus assisting in protection against seed-borne pathogens [65].
Another significant group of bioactive compounds are phenylpropanoids, which are naturally synthesized by plants. However, research has shown that endophytes also produce phenylpropanoids. These compounds belong to the largest group of secondary metabolites that possess an aromatic ring with a 3-carbon propene tail, thus resulting in a C6-C3 carbon skeleton. These compounds are from the amino acids phenylalanine and tyrosine [66]. In cancerous cells, phenylpropanoids prevent overexpression of histone deacetylase (HDAC), thus inhibiting the cell cycle and inducing apoptosis [66]. Phenylpropanoid derivatives possessing antimicrobial activities have been extracted from the endophytic fungus Aspergillus sp. (ZJ-68), associated with mangrove [67]. In another study, fusarubin and anhydrofusarubin isolated from Cladosporium species inhibited cell growth and induced apoptosis of human cancer cell lines HL-60, U937, and Jurkat [68]. Although both compounds significantly increased apoptosis of these cancerous cells with increase in concentrations, fusarubin significantly decreased the percentage of cells in the S phase while increasing those in the G2/M phase [68]. On the contrary, anhydrofusarubin increased the percentage of cells in the G0/G1 phase but decreased those in the S and G2/M phases [68]. These findings provide a valid basis for the need to focus on investigations aimed at constantly assessing the potential of endophytic fungi to produce bioactive compounds, given the evidence that they have an array of metabolites, such as alkaloids, macrolides, terpenoids, flavonoids, glycosides, xanthones, isocoumarins, quinones, phenylpropanoids, aliphatic metabolites, and lactones, with powerful anticancer properties [69]. Nonetheless, only a small proportion of endophytes have been investigated so far, thus requiring more research in the niche area.
2.4. Bioactive Compounds as a Potential Antioxidant Agent
Oxidation is a chemical reaction resulting in the loss of electrons from an atom, and this may produce free radicals. Naturally occurring free radicals are unstable molecules produced during chemical reactions such as digestion. These free radicals may participate in chain reactions which might potentially cause cell damage in the human body [70,71]. This is mainly due to the fact that after an atom losses an electron, the cell becomes imbalanced, thus resulting in cell damage. Hence, when exposed to oxidative stress, cells may suffer from a wide range of diseases that may also include chronic complications in humans [72]. Extensive studies have demonstrated that exposure of cells to oxidative stress contributes to cellular degeneration, cancer, atherosclerosis, coronary heart ailments, diabetes, Alzheimer’s disease, and hepatic and kidney damage as well as other neurodegenerative disorders [73]. Some severe side effects of oxidative stress and their associated diseases in humans are listed in Table 1.
Antioxidant agents are utilized to combat, prevent, and treat diseases that are linked with the presence of reactive oxygen species (ROS), and these agents have displayed very high efficacy against damage caused by ROS. ROS have been reported to boost the immune system by enabling cell signaling to occur. Various industries, such as the food, pharmaceutical, and agricultural sectors, use antioxidant compounds for beneficial purposes. Irrespective of the health concerns associated with oxidative stress, the search for safer, more efficacious, and cost-effective natural antioxidants is highly anticipated. Novel natural bioactive compounds have been reported to serve as a shelter against oxidative damage by preventing or reducing free radicals and reactive oxygen species. Various studies have demonstrated that molecules, including phenolic acids, phenylpropanoids, and flavonoids, lignin, melanin, and tannins, exhibited antioxidant activity [74,75]. There is considerable evidence that endophytic fungi produce several antioxidant compounds that are responsible for the stress tolerance in host plants. The endophytic fungus Fusarium oxysporum from the leaves of Otoba gracilipes exhibited antioxidant activity with as much as 51.5% of a scavenging effect on 2,2-diphenyl-1-picrylhydrazyl (DPPH) after 5 min of reaction [76]. A total of forty-one bioactive compounds from the endophyte Xylaria sp. were isolated from the medicinal plant Ginkgo biloba, and these compounds displayed antibacterial, antioxidant, anti-cardiovascular, anticancer, and antimicrobial properties. Phenolic and flavonoid compounds, among others, have been shown to possess very effective antioxidant properties [77]. Recently, studies conducted on the endophytic fungus Alternaria alternata AE1 isolated from Azadirachta indica also revealed that it produced secondary metabolites that possess very effective antioxidant properties [28]. DPPH free radical and superoxide radical scavenging tests of the secondary metabolites displayed antioxidant potentials with an IC50 value of 38.0 and 11.38 μg/mL, respectively [28]. Chemical characterization of methanol extracts of two filamentous fungal strains revealed the presence of the residues chlorogenic acid, neochlorogenic acid, rutin, and quercetin 3-acetyl-glucoside (Figure 5), and the fungal extracts displayed significant antioxidant activities [78]. In addition, another investigation identified the biomolecules pestacin, isopestacin, and 1,3-dihydro isobenzofurans from the endophytic fungus Pestalotiopsis microspore housed in Terminalia morobensis, which also displayed very effective antioxidant activities [79].
Polysaccharides produced by plants and microorganisms have been widely studied due to their natural antioxidants properties. Three polysaccharides, namely exopolysaccharide (EPS), water-extracted mycelial polysaccharide (WPS), and sodium hydroxide-extracted mycelial polysaccharide (SPS), produced by Fusarium oxysporum Dzf17 from Guazuma tomentosa displayed antioxidant properties [80]. In addition, total phenol and flavonoids detected from the culture filtrate of Phyllosticta sp. displayed significant antioxidant properties. When subjected to the 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid (ABTS) and DPPH radical assays, these compounds exhibited EC50 values of 580.02 and 2030.25 μg/mL, respectively [81].
Novel trichothecene macrolides (Figure 5) produced by the endophytic fungus Nerium oleander L (Apocynaceae) isolated from Trachelospermum jasminoides also displayed antioxidant potentials [82]. Using chemical analysis, six new macrolides comprising myrothecines D–G (1–4), 16-hydroxymytoxin B, and 14′-dehydrovertisporin and four 10,13-cyclotrichothecane derivatives were detected in three endophytes of Myrothecium roridum IFB-E008, IFB-E009, and IFB-E012. In addition, an investigation into their antimicrobial properties revealed that the compounds exhibited sufficient cytotoxicity based on their bioactive data [83].
2.5. Bioactive Compounds for Treating Infectious Parasites
Parasitic infections in humans are caused by protozoa, helminths, and ectoparasites that live on or in a host organism and make use of the host resources for their survival. Disease-causing parasites are known to contribute significantly to the rate of morbidity and mortality in humans worldwide, especially in developing countries that have a large proportion of vulnerable populations [84]. This, therefore, presents significant challenges to already overburdened public healthcare systems, thus resulting in huge economic losses. Approximately 48.4 million cases of parasitic disease resulting in one million deaths are reported annually [84]. Despite this, there are limited highly effective antiparasitic drugs current available in the market, especially given the challenges faced with resistance of parasites to the drugs [85]. Moreover, there is evidence which indicates that parasitic organisms are rapidly developing resistance against anti-parasitic drugs and the resistant strains are spreading at an alarming rate. This, therefore, calls for the need to intensify the search for novel, more potent, and less toxic compounds that may be more effective against these pathogens [86,87,88]. However, there is also substantial evidence which indicates that endophytes possess a pool of novel bioactive compounds that might be very useful in the discovery of anti-parasitic drugs.
Diaporthe phaseolorum-92C (92C), an endophytic fungus that inhabits the roots of Combretum lanceolatum, displayed significant anti-parasitic activity against Trypanosoma cruzi by reducing up to 82% of the number of amastigotes and trypomastigotes. The bioactive molecule 18-des-hydroxy Cytochalasin H exhibited nematocidal activity and reduced the viability of promastigotes of Leishmania amazonenses with an IC50 of 9.2 μg/mL [89]. Another study demonstrated that oxylipin (9Z,11E)-13-oxooctadeca-9,11-dienoic acid obtained from fungal extracts of the endophytic fungus Penicillium herquei strain BRS2A-AR was potent against Plasmodium falciparum 3D7, Trypanosoma brucei, Leishmani donovani, and Leishmania sp., with IC50 values higher than 100 μM, therefore displaying very excellent anti-parasitic activities [90]. Alternaria alternata P1210 from the roots of the halophyte Salicornia sp. produced two new biosynthesized dimeric compounds belonging to the class alternariol, namely (±)-alternarlactones A and B. Preliminary results revealed that these compounds possessed anti-parasitic potentials [91].
Mao and colleagues (2019) discovered two novel decalin/tetramic acid hybrid metabolites—hyalodendrins A and B, isolated from the endophytic fungus Hyalodendriella sp. Ponipodef12 using spectroscopic chemical analysis. These compounds were capable of inhibiting the growth of fourth-instar larvae of Aedes aegypti [92]. Bioassay was carried out using one hundred and fifty-two (n = 152) endophytic fungi to determine their anti-plasmodial activity using a 96-well microtiter plate. The results showed that extracts from Fusarium sp. AMst1 (IC50 = 1.16–1.43 µg/mL), Trichoderma afroharzianum AMrb7 (IC50 = 1.71–2.31 µg/mL), and Penicillium tropicum AMb3 (IC50 = 1.90 µg/mL) possess significantly effective anti-plasmodial activities against Plasmodium falciparum strains [93]. The findings of another study in Brazil demonstrated that the endophyte Phyllosticta capitalensis from Tibouchina granulosa (Desr.) Cogn. (Melastomataceae) inhibited Leishmania species and Trypanossoma cruzi. Based on high-resolution mass spectrum analysis (UHPLC-HRMS), 18 compounds were identified from P. capitalensis and crude extracts from the endophyte displayed growth inhibitory activities against Leishmania amazonensis, L. infantum, and Trypanosoma cruzi with IC50 values of 17.2, 82.0, and 50.13 μg/mL, respectively. Given that the diseases Leishmaniasis and Chagas are abandoned tropical diseases caused by protozoa and infect over 12 million individuals globally, these findings present a significant hope for mankind [94]. Citrinin, cochiloquinone A, palmarumycin CP18 and others (Figure 6) are amongst compounds exhibiting antiparasitic activities.
2.6. Bioactive Compounds with the Potential of Serving as Immunosuppressive Drugs
Immunosuppressive drugs, also known as antirejection medications, are utilized to suppress, reduce, or prevent allograft rejection during organ transplant in patients [95]. They therefore play a significant role in the treatment of autoimmune disorders, such as rheumatoid arthritis, lupus, psoriasis, and insulin-dependent diabetes [95]. Currently, the effectiveness of immunosuppressive drugs is affected by a number of side effects, and given that their demand is high, there is a need to hasten the search for safer but more reliable drugs in order to alleviate these problems. A number of studies have established that endophytes are capable of producing bioactive molecules with immunosuppressive potentials [96,97]. Data generated using chemical analysis revealed the presence of a novel amide derivative (-)mycousnine enamine biomolecule that was produced by the endophyte Mycosphaerella nawae ZJLQ129, isolated from Smilax china leaves [98]. Furthermore, cyclosporin A and (-)mycousnine enamine have selectively inhibited T cell proliferation by blocking the expression of the surface activation antigens CD25 and CD69. These findings confirm that endophytic fungi may serve as a potential source for potent immunosuppressants that have low toxicity but high selectivity [98]. In addition to these, a total of nine polyketides, consisting of two novel benzophenone derivatives, peniphenone and methyl peniphenone, and seven known xanthones (Figure 7), were extracted from the endophytic fungus Penicillium sp. ZJ-SY2, which was associated with mangrove Sonneratia apetala leaves. These compounds exhibited excellent immunosuppressive properties, with IC50 values ranging from 5.9 to 9.3 μg/mL [97]. Xylaria longipes HFG1018, obtained from the basidiomycete Fomitopsis betulinus, which is associated with rotting of wood, produced eighteen new nor-isopimarane diterpenes, xylarinorditerpenes A–R (1–18), some of which possessed immunosuppressive potential [99].
2.7. Bioactive Compounds with Antiviral Properties
Viruses are microorganisms that multiply only within living cells and are a leading cause of mortality and morbidity in humans globally. Current antiviral drugs and vaccines are crucial in combating life-threatening diseases in humans [100]. In addition, the emergence of resistance of viruses to the available antiviral drugs reduces the efficacy of their therapeutic potentials, thus resulting in a severe public health concern globally. In effect, ideal antiviral drugs should be potent against the target viral strains but with minimal side effects to the host cells. This therefore affirms that the modes of action of antiviral drugs are usually directed at preventing or inhibiting the infection by targeting viral proteins or the host cellular factors that viruses exploit in order to reproduce and gain control of cellular processes [101,102]. To address this problem, the search for and the discovery and development of new, cost-effective, and more potent antiviral drugs as well as vaccines is mandatory. Studies have been carried out to assess the potential of endophytes to produce promising natural bioactive compounds with antiviral properties [102].
Isoindolones compounds, namely emerimidines A and B, emeriphenolicins A and D, as well as other compounds, including aspernidines A and B, austin, austinol, dehydroaustin, and acetoxydehydroaustin, were discovered in the endophytic fungus Emericella sp. (HK-ZJ) from the mangrove plant Aegiceras corniculatum. A bioassay using the cytopathic effect (CPE) test revealed that the fungal extracts displayed potency against influenza A viral (H1N1) [103]. Fungal extracts of Nigrospora sphaerica (No.83-1-1-2), Alternaria alternata (No.58-8-4-1), and Phialophora sp. (No.96-1-8-1) exhibited some antiviral activity against herpes simplex virus (HSV). Extraction and identification of compounds revealed two novel heptaketides, (+)-(2S,3S,4aS)-altenuene (1a) and (−)-(2S,3S,4aR)-isoaltenuene, along with six recognizable compounds, (−)-(2R,3R,4aR)-altenuene, (+)-(2R,3R,4aS)-isoaltenuene, 5′-methoxy-6-methyl-biphenyl-3,4,3′-triol, alternariol (4), alternariol-9-methyl ether, and 4-hydroxyalternariol-9-methyl ether [104].
Recently, endophytic fungi were isolated from medicinal plants of Egyptian origin, which displayed significant antiviral properties against herpes simplex (HSV-2) and vesicular stomatitis viruses (VSV) [105]. It was identified that the endophyte Pleospora tarda was responsible for the potent antiviral agents classified as alternariol and alternariol-(9)-methyl compounds [105]. Lui et al. (2019) identified a new rare 14-nordrimane sesquiterpenoid, extracted from the endophyte Phoma sp., isolated from the roots of Aconitum vilmorinianum [106]. The compounds also inhibited the growth of influenza A virus (A/Puerto Rico/8/34, H1N1). Other bioactive compounds with antifungal activities included (–)-6-methoxymellein, 7-hydroxy-3, 5-dimethyl-isochromen-1-one, norlichexanthone, 6-methylsalicylic acid, and gentisyl alcohol [106]. In addition, the hydroanthraquinone (Figure 8) derivative, 6-O-demethyl-4-dehydroxyaltersolanol A, azaphilones, 8,11-didehydrochermesinone B, and (7S)-7-hydroxy-3,7-dimethyl-isochromene-6,8-dione have recently been identified as compounds from the culture extract of Nigrospora sp. YE3033 which resides in the plant Aconitum carmichaeli. A preliminary bioassay indicated that these compounds displayed strong antiviral activity against the influenza viral strain A/Puerto Rico/8/34 (H1N1) [107].
An investigation of endophytes associated with Penicillium sp. FKI-7127 led to the detection of the brefeldin A compound (Figure 8), exhibiting potent antiviral properties [108]. Several endophytic fungal strains, including Fusarium equiseti, Scopulariopsis fusca, and Geotrichum candidum, were obtained from brown alga Padina pavonica, located in the Red Sea. Out of these fungi, F. equiseti exhibited the highest antiviral activity against hepatitis C virus (HCV) NS3-NS4A protease, with an IC50 of 27.0 µg/mL. Structural characterization using MS and NMR spectral analysis of the metabolites from this endophyte showed the presence of two diketopiperazines (Figure 8) (cyclo-L-AlaL-Leu and cyclo(L-Tyr-L-Pro)) and two nucleosides (cordycepin and Ara-A) [109]. To date, more than one hundred endophytic fungi isolated from desert plants have been established to possess potent antiretroviral activities against human immunodeficiency virus type 1 (HIV-1). The extracts from these fungi demonstrated less than 30% cytotoxic activities in T lymphocytes [110]. Three unidentified chromanones were extracted from the fungal strain Phomopsis sp. CGMCC No. 5416, obtained from the stems of Achyranthes bidentata. These compounds displayed promising antiviral activities against HIV-1 [111]; we therefore suggest that fungal secondary metabolites may serve as important sources for discovering new antiviral drugs or lead compounds.
2.8. Bioactive Compounds as Potential Antitubercular Drugs
Tuberculosis (TB) is a life-threatening disease usually affecting the lungs and is caused by the bacteria Mycobacterium tuberculosis. The risk of contracting TB is significantly higher among individuals whose immune system has been compromised and children under the age of 5 years, particularly in developing countries, such as China, Indonesia, the Philippines, Pakistan, Nigeria, Bangladesh, and South Africa [112]. Infectious TB is most often associated with high fatality rates in humans worldwide [112,113]. Given that TB is more common in people with compromised immune systems, there is evidence of co-infection between TB and HIV in patients, and the World Health Organization (WHO) reported that of the 1.5 million patients who died from TB in 2018, 251,000 were also infected with HIV [112]. In addition, the multi-drug resistance of Mycobacterium tuberculosis against available drugs is increasing at an alarming rate and has eventually become an issue of severe public health concern [113]. Approximately 484,000 new cases of resistance to the first-line antimicrobial drug rifampicin, which was regarded as the most potent drug, were reported, which is now a severe public health threat [114] and, thus, requires urgent attention. Nonetheless, tuberculosis is curable and preventable; thus, the constant search for natural bioactive compounds, especially from endophytes, now paves the way for the discovery of new, more effective, alternative agents to combat tuberculosis. Gliocladium sp. MR41 was capable of producing polyols 3 and 4 (Figure 9), compounds that displayed inhibitory activities against M. tuberculosis at a minimum inhibitory concentration (MIC) of 0.78 µg/mL [115]. Other compounds with potential antituberculosis activities include phomoenamide, abyssomicin, tenuazonic acid and phomonitroester (Figure 9).
These were the first reports of such compounds being produced by a fungus [115]. Glycyrrhiza glabra L. plant cultivated in the Kashmir Himalayas harbored a wide diversity of endophytic fungi that comprised, but were not limited to, Fusarium oxysporum strain (KT166447) and Colletotrichum gleosporoides strain (KT166445), which displayed strong inhibitory potentials against Mycobacterium tuberculosis (M. tb) strain H37Rv, with MIC values of 18.5 and 75 µg/mL, respectively [116]. In a recent study involving a review of findings between 2014 and 2015 detailing various biological activities, including antituberculosis, and that endorses fungal strains as a source of endless bioactive compounds, it was revealed that endophytes provide a renewed hope for the detection and development of potential antitubercular drugs [117]. Table 2 listed several endophytic fungi isolated from various host plants that were capable of producing compounds with several bioactive activities.
3. Conclusions
Currently, we are losing the fight against ineffective, toxic, and expensive therapeutic antimicrobial drugs. However, endophytes provide a suitable alternative since they are a warehouse filled with novel bioactive compounds with endless possibilities of biological properties. Over the past few years, endophytic fungi have attracted tremendous attention in the drug development process as they are ubiquitous and abundantly availability. Numerous studies have reported novel, beneficial bioactive compounds exhibiting biological properties, such as antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, insecticidal, immunomodulatory, antiviral, anticancer activities, anthelmintic, etc., that were successfully isolated from endophytic fungi. Despite this, limited research has been conducted on the valuable bioactive compounds from endophytic fungi. Research priorities need to shift towards biotechnological advances to accelerate the screening of new biomolecules for the treatment of numerous life-threatening diseases, thus safe-guarding human health. All things considered, an untapped wealth of novel bioactive compounds resides within endophytes, thus ensuring the discovery of new bioactive compounds for potential applications in the agricultural, food, medical, and pharmaceutical industries.
Author Contributions
Both authors contributed to the drafting, writing, and editing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
We would like to thank the North West University, School of Biological Science, Department of Microbiology and the National Research Foundation (NRF, Grant UID No: 105510) for the financial support.
Acknowledgments
The authors acknowledge the Department of Microbiology, School of Biological Science, North West University, South Africa, for their continues support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: Causes and control strategies. Antimicrob. Resist. Infect. Control. 2017, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- World Health Organization (WHO). Traditional Medicine. World Health Organization. 2008. Available online: http://www.who.int/mediacentre/factsheets/2003/fs134/en/ (accessed on 11 September 2015).
- Mukherjee, P.K. Trichoderma–Genetics, Genomics and Beyond. Indian Phytopathol. 2015, 68, 1–7. [Google Scholar]
- Li, F.; Weng, J. Demystifying traditional herbal medicine with modern approach. Nat. Plants 2017, 3, 17109. [Google Scholar] [CrossRef]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1123. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Kaul, S.; Gupta, S.; Ahmed, M.; Dhar, M.K. Endophytic fungi from medicinal plants: A treasure hunt for bioactive metabolites. Phytochem. Rev. 2012, 11, 487–505. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.S.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ren, L.; Li, C.; Gao, C.; Liu, X.; Wang, M.; Luo, L. Effects of endophytic fungi diversity in different coniferous species on the colonization of Sirex noctilio (Hymenoptera: Siricidae). Sci. Rep. 2019, 9, 5077. [Google Scholar] [CrossRef] [PubMed]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.M.; Salim, S.S.; Hassan, S.E.D.; Ismail, M.A.; Fouda, A. Role of endophytes in plant health and abiotic stress management. In Microbiome in Plant Health and Disease; Springer: Singapore, 2019; pp. 119–144. [Google Scholar]
- Tan, S.Y.; Tatsumura, Y. Alexander Fleming (1881–1955): Discoverer of penicillin. Singap. Med. J. 2015, 56, 366–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [Green Version]
- Fadiji, A.E.; Babalola, O.O. Elucidating mechanisms of endophytes used in plant protection and other bioactivities with multifunctional prospects. Front. Bioeng. Biotechnol. 2020, 8, 467. [Google Scholar] [CrossRef]
- Toghueo, R.M.K. Bioprospecting endophytic fungi from Fusarium genus as sources of bioactive metabolites. Mycology 2020, 11, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and treats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Centers for Disease Control and Prevention, Office of Infectious Disease. Antibiotic Resistance Threats in the United States. Available online: http://www.cdc.gov/drugresistance/threat-report-2013 (accessed on 21 June 2020).
- World Health Organization (WHO). Fact Sheets. Antibiotic Resistance. Available online: http://www.who.int/news-room/fact-sheet/detail/antibiotic-resistance (accessed on 21 June 2020).
- Ravindra, P.A.; Suneel, K.; Sardul, S.S. Endophytic mycoflora as a source of biotherapeutic compounds for disease treatment. J. Appl. Pharm. Sci. 2016, 6, 242–254. [Google Scholar]
- Pavithra, G.; Sumant, B.; Meenakshi, R.; Seweta, S. Role of Endophytic Microbes against Plant Pathogens. Asian J. Plant Sci. 2020, 19, 54–62. [Google Scholar]
- Tirsit, T. Pharmaceutical effects of bioactive compounds from endophytes. Int. J. Innov. Pharm. Sci. Res. 2018, 1, 1–13. [Google Scholar]
- Chatterjee, S.; Ghosh, R.; Mandal, N.C. Production of bioactive compounds with bactericidal and antioxidant potential by endophytic fungus Alternaria alternata AE1 isolated from Azadirachta indica A. Juss. PLoS ONE 2019, 14, e0214744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria do Carmo, M.; Carvalho, C.M.; Faria, F.S.; Nobreza, A.M.; Thiago, M. Antibacterial Activity of Phytochemical extracts and endophytic fungi of Carapa guianensis against Enterococcus faecalis in endodontic infections an in vitro study. Open Dent. J. 2019, 13, 249–254. [Google Scholar]
- Manganyi, M.C.; Tchatchouang, C.D.K.; Regnier, T.; Bezuidenhout, C.C.; Ateba, C.N. Bioactive compound produced by endophytic fungi isolated from Pelargonium sidoides against selected bacteria of clinical importance. Mycobiology 2019, 47, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Varkey, S.; Anith, K.N.; Narayana, R.; Aswini, S. A consortium of rhizobacteria and fungal endophyte suppress the root-knot nematode parasite in tomato. Rhizosphere 2018, 5, 38–42. [Google Scholar] [CrossRef]
- Kjer, J.; Wray, V.; Edrada-Ebel, R.; Ebel, R.; Pretsch, A.; Lin, W.H.; Proksch, P. Xanalteric acids I and II and related phenolic compounds from an endophytic Alternaria sp. isolated from the mangrove plant Sonneratia alba. J. Nat. Prod. 2009, 72, 2053–2057. [Google Scholar] [CrossRef]
- Uzma, F.; Mohan, C.D.; Siddaiah, C.N.; Chowdappa, S. Endophytic fungi: Promising source of novel bioactive compounds. In Advances in Endophytic Fungal Research; Springer: Cham, Switzerland, 2019; pp. 243–265. [Google Scholar]
- Ding, Z.; Tao, T.; Wang, L.; Zhao, Y.; Huang, H.; Zhang, D.; Liu, M.; Wang, Z.; Han, J. Bioprospecting of Novel and Bioactive Metabolites from Endophytic Fungi Isolated from Rubber Tree Ficus elastica Leaves. Microbiol. Biotechnol. 2019, 29, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Findlay, J.A.; Li, G.; Johnson, J.A. Bioactive compounds from an endophytic fungus from eastern larch (Larix laricina) needles. Can. J. Chem. 1997, 75, 716–719. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Luecking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5, 0052-2016. [Google Scholar]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The Still Underestimated Problem of Fungal Diseases Worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomazzi, J.; Baethgen, L.; Carneiro, L.C.; Millington, M.A.; Denning, D.W.; Colombo, A.L.; Pasqualotto, A.C. The burden of serious human fungal infections in Brazil. Mycoses 2015, 59, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Peláez, F.; Cabello, A.; Platas, G.; Díez, M.T.; del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Chepkirui, C.; Richter, C.; Matasyoh, J.C.; Stadler, M. Monochlorinated calocerins A-D and 9-oxostrobilurin derivatives from the basidiomycete Favolaschia calocera. Phytochemistry 2016, 132, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Haryani, Y.; Hilma, R.; Delfira, N.; Martalinda, T.; Puspita, F.; Friska, A.; Juwita, D.; Farniga, A.; Ardi, F. Potential Antibacterial Activity of Endophytic Fungi Penicillium sp. and Trichoderma sp. Derived From Mangrove Ceriops Tagal (Perr.) CB Robb and Bruguiera sp. J. Phys. Conf. Ser. 2019, 1351, 012100. [Google Scholar] [CrossRef]
- Li, E.; Jiang, L.; Guo, L.; Zhang, H.; Che, Y. Pestalachlorides A-C, antifungal metabolites from the plant endophytic fungus Pestalotiopsis adusta. Bioorg. Med. Chem. 2008, 16, 7894–7899. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Choi, S.; Kim, S.; Lee, J.H.; Park, A.R.; Yu, N.H.; Yoon, H.; Bae, C.H.; Yeo, J.H.; Choi, G.J.; et al. The Hsp90 Inhibitor, Monorden, Is a Promising Lead Compound for the Development of Novel Fungicides. Front. Plant Sci. 2020, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.L.; Wei, W.J.; Li, H.Y.; Wang, L.D.; Dong, S.H.; Gao, K. Phomotide A, a novel polyketide, from the endophytic fungus Phomopsis sp. CFS42. Tetrahedron Lett. 2020, 61, 151468. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Fact Sheets. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 24 June 2020).
- Batra, H.; Pawar, S.; Bahl, D. Curcumin in combination with anti-cancer drugs: A nanomedicine review. Pharmacol. Res. 2019, 139, 91–105. [Google Scholar] [CrossRef]
- Peng, C.; Xu, J.; Yu, M.; Ning, X.; Huang, Y.; Du, B.; Hernandez, E.; Kapur, P.; Hsieh, J.T.; Zheng, J. Tuning the in-Vivo Transport of Anticancer Drugs Using Renal-Clearable Gold Nanoparticles. Angew. Chem. 2019, 131, 8567–8571. [Google Scholar]
- Tewari, D.; Rawat, P.; Singh, P.K. Adverse drug reactions of anticancer drugs derived from natural sources. Food Chem. Toxicol. 2019, 123, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 7, 315. [Google Scholar] [CrossRef]
- Das, A.; Rahman, M.I.; Ferdous, A.S.; Amin, A.; Rahman, M.M.; Nahar, N.; Uddin, M.A.; Islam, M.R.; Khan, H. An endophytic Basidiomycete, Grammothele lineata, isolated from Corchorus olitorius, produces paclitaxel that shows cytotoxicity. PLoS ONE 2017, 12, e0178612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, W.; Tang, T.; Ling, F. Comparative transcriptome analysis of a taxol-producing endophytic fungus, Aspergillus aculeatinus Tax-6, and its mutant strain. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.; Vasundhara, M. Isolation of taxol producing endophytic fungus Alternaria brassicicola from non-Taxus medicinal plant Terminalia arjuna. World J. Microbiol. Biotechnol. 2019, 35, 74. [Google Scholar] [CrossRef] [PubMed]
- El-Bialy, H.A.; El-Bastawisy, H.S. Elicitors stimulate paclitaxel production by endophytic fungi isolated from ecologically altered Taxus baccata. J. Radiat. Res. Appl. 2020, 13, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–226. [Google Scholar] [CrossRef]
- Kumar, G.; Chandra, P.; Choudhary, M. Endophytic fungi: A potential source of bioactive compounds. Chem. Sci. Rev. Lett. 2017, 6, 2373–2381. [Google Scholar]
- Shweta, S.; Zuehlke, S.; Ramesha, B.T. Endophytic fungal strains of Fusarium solani, from Apodytes dimidiate E. Mey. ex Arn (Icacinaceae) produce camptothecin, 10-hydroxycamptothecin and 9-methoxycamptothecin. Phytochemistry 2010, 71, 117–122. [Google Scholar] [CrossRef]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A. Plant antitumor agents. VI. The isolation and structure of taxol, a novel anti leukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef]
- Li, X.; Tian, Y.; Yang, S.-X.; Zhang, Y.-M.; Qin, J.-C. Cytotoxic azaphilone alkaloids from Chaetomium globosum TY1. Bioorg. Med. Chem. Lett. 2013, 23, 2945–2947. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Negi, C.; Yadav, A.N.; Yadav, N.; Singh, K.; Saxena, A.K. Endophytic fungi from medicinal plants: Biodiversity and biotechnological applications. In Microbial Endophytes; Woodhead Publishing: Cambridge, UK, 2020; pp. 273–305. [Google Scholar]
- Wagenaar, M.M.; Corwin, J.; Strobel, G.; Clardy, J. Three new cytochalasins produced by an endophytic fungus in the genus Rhinocladiella. J. Nat. Prod. 2000, 63, 1692–1695. [Google Scholar] [CrossRef] [PubMed]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on endophytic fungi related to secondary metabolites and biotransformation applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.C.; Nazir, A.; Chawla, R.; Arora, R.; Riyaz-ul-Hasan, S.; Amna, T.; Ahmed, B.; Verma, V.; Singh, S.; Sagar, R.; et al. The endophytic fungus Trametes hirsuta as a novel alternative source of podophyllotoxin and related aryl tetralin lignans. J. Biotechnol. 2006, 122, 494–510. [Google Scholar] [CrossRef]
- Tidke, S.A.; Kumar, R.K.; Ramakrishna, D.; Kiran, S.; Kosturkova, G.; Gokare, R.A. Current understanding of endophytes: Their relevance, importance, and industrial potentials. J. Biotechnol. Biochem. 2017, 3, 43–59. [Google Scholar] [CrossRef]
- Baker, S.; Satish, S. Endophytes: Natural warehouse of bioactive compounds. Drug Invent. Today 2012, 4, 548–553. [Google Scholar]
- Sarika, G.; Amruta, N.; Kandikattu, H.K.; Basavaraju, G.V.; Suma, H.K.; Manjunath, B.L.; Sravani, C.H. Chemical profiling of camptothecin and methoxy camptothecin in Nothapodytes nimmoniana Grah.(Mabb.) during seed development, seed germination and their effects on seed-borne pathogens. S. Afr. J. Bot. 2019, 123, 113–123. [Google Scholar] [CrossRef]
- Neelam, K.A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. Nutr. 2019, 1–21. [Google Scholar]
- Cai, R.; Jiang, H.; Zang, Z.; Li, C.; She, Z. New benzofuranoids and phenylpropanoids from the mangrove endophytic fungus, Aspergillus sp. ZJ-68. Mar. Drugs 2019, 17, 478. [Google Scholar] [CrossRef] [Green Version]
- Adorisio, S.; Fierabracci, A.; Muscari, I.; Liberati, A.M.; Cannarile, L.; Thuy, T.T.; Sung, T.V.; Sohrab, H.; Hasan, C.M.; Ayroldi, E.; et al. Fusarubin and anhydrofusarubin isolated from a Cladosporium species inhibit cell growth in human cancer cell lines. Toxins 2019, 11, 503. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.W.; Zhang, Y.L.; Lin, F.C.; Hu, Y.Z.; Zhang, C.L. Natural products with antitumor activity from endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 1056–1074. [Google Scholar] [CrossRef] [PubMed]
- Elochukwui, C. Generation and reaction of free radicals in the human body: A major cause of aging and chronic degenerative diseases. EC Nutr. 2015, 1, 132–136. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 1, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Nasri, H.; Rafieian-Kopaei, M. Oxidative stress and aging prevention. Int. J. Prev. Med. 2013, 4, 1101–1102. [Google Scholar]
- Patipong, T.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Induction of antioxidative activity and antioxidant molecules in the halotolerant cyanobacterium Halothece sp. PCC7418 by temperature shift. Nat. Prod. Commun. 2019, 14, 1934578–19865680. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Doyle, S.; Murphy, R. Filamentous fungi as a source of natural antioxidants. Food Chem. 2015, 185, 389–397. [Google Scholar] [CrossRef]
- Caicedo, N.H.; Davalos, A.F.; Puente, P.A.; Rodríguez, A.Y.; Caicedo, P.A. Antioxidant activity of exo-metabolites produced by Fusarium oxysporum: An endophytic fungus isolated from leaves of Otoba gracilipes. Microbiol. Open 2019, 8, 903. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Dong, M.; Chen, X.; Jiang, M.; Lv, X.; Yan, G. Antioxidant activity and phenolics of an endophytic Xylaria sp. from Ginkgo biloba. Food Chem. 2007, 105, 548–554. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Pintea, A. Phenolic compounds, flavonoids, lipids and antioxidant potential of apricot (Prunus armeniaca L.) pomace fermented by two filamentous fungal strains in solid state system. Chem. Cen. J. 2017, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Strobel, G.; Ford, E.; Worapong, J.; Harper, J.K.; Arif, A.M.; Grant, D.M.; Fung, P.C.W.; Ming Wah Chau, R. Isopestacin, an isobenzofuranone from Pestalotiopsis microspora, possessing antifungal and antioxidant activities. Phytochemistry 2002, 60, 179–183. [Google Scholar] [CrossRef]
- Li, P.; Luo, C.; Sun, W.; Lu, S.; Mou, Y.; Peng, Y.; Zhou, L. In-vitro antioxidant activities of polysaccharides from endophytic fungus Fusarium oxysporum Dzf17. Afr. J. Microbiol. Res. 2011, 5, 5994–5997. [Google Scholar]
- Srinivasan, K.; Jagadish, L.K.; Shenbhagaraman, R.; Muthumary, J. Antioxidant activity of endophytic fungus Phyllosticta sp. isolated from Guazuma tomentosa. J. Phytol. 2010, 1, 37–41. [Google Scholar]
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Endophytic fungi from Nerium oleander L (Apocynaceae): Main constituents and antioxidant activity. World J. Microbiol. Biotechnol. 2007, 23, 1253–1263. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Ai, C.Z.; Song, Y.C.; Wang, F.W.; Jiao, R.H.; Zhang, A.H.; Man, H.Z.; Tan, R.X. Cytotoxic trichothecene macrolides produced by the endophytic Myrothecium roridum. J. Nat. Prod. 2019, 82, 1503–1509. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization estimates of the global and regional disease burden of 11 foodborne parasitic diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef] [Green Version]
- Skinner-Adams, T.S.; Sumanadasa, S.D.; Fisher, G.M.; Davis, R.A.; Doolan, D.L.; Andrews, K.T. Defining the targets of antiparasitic compounds. Drug Discov. Today 2016, 21, 725–739. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Wang, C.; Tian, B. Modelling intra-host competition between malaria parasites strains. Comput. Appl. Math. 2020, 39, 48. [Google Scholar] [CrossRef]
- Vermelho, A.B.; Rodrigues, G.C.; Supuran, C.T. Why hasn’t there been more progress in new Chagas disease drug discovery? Expert Opin. Drug Discov. 2020, 15, 145–158. [Google Scholar] [CrossRef]
- Rottmann, M.; McNamara, C.; Yeung, B.K.; Lee, M.C.; Zou, B.; Russell, B.; Seitz, P.; Plouffe, D.M.; Dharia, N.V.; Tan, J.; et al. Spiroindolones, a potent compound class for the treatment of malaria. Science 2010, 329, 1175–1180. [Google Scholar] [CrossRef] [Green Version]
- Brissow, E.R.; da Silva, I.P.; de Siqueira, K.A.; Senabio, J.A.; Pimenta, L.P.; Januário, A.H.; Magalhães, L.G.; Furtado, R.A.; Tavares, D.C.; Junior, P.A.S.; et al. 18-Des-hydroxy Cytochalasin: An antiparasitic compound of Diaporthe phaseolorum-92C, an endophytic fungus isolated from Combretum lanceolatum Pohl ex Eichler. Parasit. Res. 2017, 116, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Hayibor, K.; Kwain, S.; Osei, E.; Nartey, A.P.; Tetevi, G.M.; Owusu, K.B.A.; Camas, M.; Camas, A.S.; Kyeremeh, K. Ghanaian mangrove wetland endophytic fungus, Penicillium herquei strain BRS2A-AR produces (9Z, 11E)-13-oxooctadeca-9, 11-dienoic acid with activity against Trichomonas mobilensis. Int. J. Biol. Chem. Sci. 2019, 13, 1918–1937. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.N.; Pusch, S.; Shi, Y.M.; Richter, C.; Macia-Vicente, J.G.; Schwalbe, H.; Kaiser, M.; Opatz, T.; Bode, H.B. (±)-Alternarlactones A and B, Two Antiparasitic Alternariol-like Dimers from the Fungus Alternaria alternata P1210 isolated from the halophyte Salicornia sp. Org. Chem. 2019, 84, 11203–11209. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Wang, W.; Su, R.; Gu, G.; Liu, Z.L.; Lai, D.; Zhou, L. Hyalodendrins A and B, New decalin-type tetramic acid larvicides from the endophytic fungus Hyalodendriella sp. Ponipodef12. Molecules 2020, 25, 114. [Google Scholar] [CrossRef] [Green Version]
- Toghueo, R.M.K.; Kemgne, E.A.M.; Eke, P.; Kanko, M.I.M.; Dize, D.; Sahal, D.; Boyom, F.F. Antiplasmodial potential and GC-MS fingerprint of endophytic fungal extracts derived from Cameroonian Annona muricata. J. Ethnopharmacol. 2019, 235, 111–121. [Google Scholar] [CrossRef]
- Golias, H.C.; Polonio, J.C.; dos Santos Ribeiro, M.A.; Polli, A.D.; da Silva, A.A.; Bulla, A.M.; Volpato, H.; Nakamura, C.V.; Meurer, E.C.; Azevedo, J.L.; et al. Tibouchina granulosa (Vell.) Cogn (Melastomataceae) as source of endophytic fungi: Isolation, identification, and antiprotozoal activity of metabolites from Phyllosticta capitalensis. Braz. J. Microbiol. 2020, 51, 557–569. [Google Scholar] [CrossRef]
- Van Laar, J.M. Immunosuppressive Drugs. In Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Firestein, G.S., Budd, R.C., Gabriel, S.E., McInnes, I.B., O’Dell, J.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Chapter 62; pp. 983–998. [Google Scholar]
- Wang, W.X.; Cheng, G.G.; Li, Z.H.; Ai, H.L.; He, J.; Li, J.; Feng, T.; Liu, J.K. Curtachalasins, immunosuppressive agents from the endophytic fungus Xylaria cf. curta. Org. Biomol. Chem. 2019, 17, 7985–7994. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Liu, W.; Liu, Y.; Huang, X.; She, Z. Polyketides with immunosuppressive activities from mangrove endophytic fungus Penicillium sp. ZJ-SY2. Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.W.; Wang, J.L.; Chen, J.; Chen, J.J.; Shen, J.W.; Feng, X.X.; Kubicek, C.P.; Lin, F.C.; Zhang, C.L.; Chen, F.Y. A novel derivative of (-) mycousnine produced by the endophytic fungus Mycosphaerella nawae, exhibits high and selective immunosuppressive activity on T cells. Front. Microbiol. 2017, 8, 1251. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.P.; Zhao, Z.Z.; Cheng, G.G.; Zhao, K.; Han, K.Y.; Zhou, L.; Feng, T.; Li, Z.H.; Liu, J.K. Immunosuppressive Nor-isopimarane Diterpenes from cultures of the Fungicolous Fungus Xylaria longipes HFG1018. J. Nat. Prod. 2020, 83, 401–412. [Google Scholar] [CrossRef]
- Andersen, P.I.; Ianevski, A.; Lysvand, H.; Vitkauskiene, A.; Oksenych, V.; Bjørås, M.; Telling, K.; Lutsar, I.; Dampis, U.; Irie, Y.; et al. Discovery and development of safe-in-man broad-spectrum antiviral agents. Int. J. Infec. Dis. 2020, 93, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Linnakoski, R.; Reshamwala, D.; Veteli, P.; Cortina-Escribano, M.; Vanhanen, H.; Marjomäki, V. Antiviral agents from fungi: Diversity, mechanisms and potential applications. Front. Microbiol. 2018, 9, 2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, S.; Devincenzo, J.P.; Toovey, S.; Wu, J.Z.; Whitley, R.J. Comparison of antiviral resistance across acute and chronic viral infections. Antivir. Res. 2018, 158, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, S.; Zhu, T.; Lin, Z.; Gu, J.; Li, D.; Gu, Q. Antiviral isoindolone derivatives from an endophytic fungus Emericella sp. associated with Aegiceras corniculatum. Phytochemistry 2011, 72, 1436–1442. [Google Scholar] [CrossRef]
- He, J.W.; Chen, G.D.; Gao, H.; Yang, F.; Li, X.X.; Peng, T.; Guo, L.D.; Yao, X.S. Heptaketides with antiviral activity from three endolichenic fungal strains Nigrospora sp., Alternaria sp. and Phialophora sp. Fitoterapia 2012, 83, 1087–1091. [Google Scholar] [CrossRef]
- Selim, K.A.; Elkhateeb, W.A.; Tawila, A.M.; El-Beih, A.A.; Abdel-Rahman, T.M.; El-Diwany, A.I.; Ahmed, E.F. Antiviral and antioxidant potential of fungal endophytes of Egyptian medicinal plants. Fermentation 2018, 4, 49. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.S.; Jiang, J.X.; Huang, R.; Wang, Y.T.; Jiang, B.G.; Zheng, K.X.; Wu, S.H. A new antiviral 14-nordrimane sesquiterpenoid from an endophytic fungus Phoma sp. Phytochem. Lett. 2019, 29, 75–78. [Google Scholar] [CrossRef]
- Zhang, S.P.; Huang, R.; Li, F.F.; Wei, H.X.; Fang, X.W.; Xie, X.S.; Lin, D.G.; Wu, S.H.; He, J. Antiviral anthraquinones and azaphilones produced by an endophytic fungus Nigrospora sp. Aconitum carmichaeli. Fitoterapia 2016, 112, 85–89. [Google Scholar] [CrossRef]
- Raekiansyah, M.; Mori, M.; Nonaka, K.; Agoh, M.; Shiomi, K.; Matsumoto, A.; Morita, K. Identification of novel antiviral of fungus-derived brefeldin A against dengue viruses. Trop. Med. Health 2017, 45, 32. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; Al-Farawati, R. Chemical constituents and antiviral activity from marine endophytic fungi from Red Sea alga Padina pavonica. J. Chem. Soc. Pak. 2017, 39, 478. [Google Scholar]
- Wellensiek, B.P.; Ramakrishnan, R.; Bashyal, B.P.; Eason, Y.; Gunatilaka, A.L.; Ahmad, N. Inhibition of HIV-1 replication by secondary metabolites from endophytic fungi of desert plants. Open Virol. J. 2013, 7, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Wu, K.; Xu, Y.; Xia, X.; Wang, X.; Ge, M.; Shao, L. Three novel chromanones with biological activities from the endophytic fungus Phomopsis CGMCC No. 5416. J. Antibiot. 2020, 73, 194–199. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Consolidated Guidelines on Tuberculosis: Tuberculosis Preventive Treatment: Module 1: Prevention: Tuberculosis Preventive Treatment. Available online: https://apps.who.int/iris/bitstream/handle/10665/331170/9789240001503-eng.pdf (accessed on 25 June 2020).
- Schön, T.; Miotto, P.; Köser, C.U.; Viveiros, M.; Böttger, E.; Cambau, E. Mycobacterium tuberculosis drug-resistance testing: Challenges, recent developments and perspectives. Clin. Microbiol. Infect. 2017, 23, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Fact sheets. Tuberculosis. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 27 June 2020).
- Uc-Cachón, A.H.; Gamboa-Angulo, M.; Borges-Argáez, R.; Reyes-Estebanez, M.; Said-Fernández, S.; Molina-Salinas, G.M. Antitubercular activity of the fungus Gliocladium sp. MR41 strain. Iran. J. Pharm. Res. IJPR 2019, 18, 860. [Google Scholar] [PubMed]
- Shah, A.; Rather, M.A.; Shah, A.M.; Mushtaq, S.; Hussain, A.; Rasool, S.; Parry, Z.A.; Qazi, P.H. Evaluating the in vitro antituberculosis, antibacterial and antioxidant potential of fungal endophytes isolated from Glycyrrhiza glabra L. Ann. Phytomed. 2016, 5, 140–146. [Google Scholar] [CrossRef]
- Mayer, A.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine pharmacology in 2014–2015: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, antiviral, and anthelmintic activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2020, 18, 5. [Google Scholar]
- Ikram, M.; Ali, N.; Jan, G.; Hamayun, M.; Jan, F.G.; Iqbal, A. Novel antimicrobial and antioxidative activity by endophytic Penicillium roqueforti and Trichoderma reesei isolated from Solanum surattense. Acta Physiol. Plant. 2019, 41, 164. [Google Scholar] [CrossRef]
- Chaithra, M.; Vanitha, S.; Ramanathan, A.; Jegadeeshwari, V.; Rajesh, V.; Hegde, V.; Apshara, E. Profiling Secondary Metabolites of Cocoa (Theobroma cacao L.) Endophytic Fungi Lasiodiplodia pseudotheobromae PAK-7 and Lasiodiplodia theobromae TN-R-3 and their antimicrobial activities. Curr. J. Appl. Sci. Technol. 2020, 39, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Wonglom, P.; Ito, S.I.; Sunpapao, A. Volatile organic compounds emitted from endophytic fungus Trichoderma asperellum T1 mediate antifungal activity, defense response and promote plant growth in lettuce (Lactuca sativa). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Yehia, R.S.; Osman, G.H.; Assaggaf, H.; Salem, R.; Mohamed, M.S. Isolation of potential antimicrobial metabolites from endophytic fungus Cladosporium cladosporioides from endemic plant Zygophyllum mandavillei. S. Afr. J. Bot. 2020, 129, 1–7. [Google Scholar] [CrossRef]
- Cao, X.; Shi, Y.; Wu, S.; Wu, X.; Wang, K.; Sun, H.; He, S.; Dickschat, J.S.; Wu, B. Polycyclic meroterpenoids, talaromyolides E−K for antiviral activity against pseudorabies virus from the endophytic fungus Talaromyces purpureogenus. Tetrahedron 2020, 76, 131349. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Lin, X.P.; Wang, Z.; Zhou, X.F.; Qin, X.C.; Kaliyaperumal, K.; Zhang, T.Y.; Tu, Z.C.; Liu, Y. Asteltoxins with Antiviral Activities from the Marine Sponge-Derived Fungus Aspergillus sp. SCSIO XWS02F40. Molecules 2015, 21, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seetharaman, P.; Gnanasekar, S.; Chandrasekaran, R.; Chandrakasan, G.; Kadarkarai, M.; Sivaperumal, S. Isolation and characterization of anticancer flavone chrysin (5, 7-dihydroxy flavone)-producing endophytic fungi from Passiflora incarnata L. leaves. Ann. Microbiol. 2017, 67, 321–331. [Google Scholar] [CrossRef]
- Da Rosa, B.V.; Kuhn, K.R.; Ugalde, G.A.; Zabot, G.L.; Kuhn, R.C. Antioxidant compounds extracted from Diaporthe schini using supercritical CO 2 plus cosolvent. Bioprocess Biosyst. Eng. 2020, 43, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Druzian, S.P.; Pinheiro, L.N.; Susin, N.M.B.; Dal Prá, V.; Mazutti, M.A.; Kuhn, R.C.; de Marsillac Terra, L. Production of metabolites with antioxidant activity by Botryosphaeria dothidea in submerged fermentation. Bioprocess Biosyst. Eng. 2020, 43, 13–20. [Google Scholar] [CrossRef]
- Ran, X.; Zhang, G.; Li, S.; Wang, J. Characterization and antitumor activity of camptothecin from endophytic fungus Fusarium solani isolated from Camptotheca acuminate. Afr. Health Sci. 2017, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Rather, M.A.; Hassan, Q.P.; Aga, M.A.; Mushtaq, S.; Shah, A.M.; Hussain, A.; Baba, S.A.; Ahmad, Z. Discovery of anti-microbial and anti-tubercular molecules from Fusarium solani: An endophyte of Glycyrrhiza glabra. J. Appl. Microbiol. 2017, 122, 1168–1176. [Google Scholar] [CrossRef]
Figure 1.
Timeline of events in the development of antibiotic resistance [23].
Figure 1.
Timeline of events in the development of antibiotic resistance [23].



Figure 3.
Chemical structures of the bioactive compounds with antifungal activities [33].
Figure 3.
Chemical structures of the bioactive compounds with antifungal activities [33].



Figure 5.
Chemical structures of the bioactive compounds with antioxidant activities [12].
Figure 5.
Chemical structures of the bioactive compounds with antioxidant activities [12].


Figure 6.
Chemical structures of the bioactive compounds with antiparasitic activities [20].
Figure 6.
Chemical structures of the bioactive compounds with antiparasitic activities [20].

Figure 7.
Chemical structures of the bioactive compounds with immunosuppressive activities [99].
Figure 7.
Chemical structures of the bioactive compounds with immunosuppressive activities [99].




{kind=link}
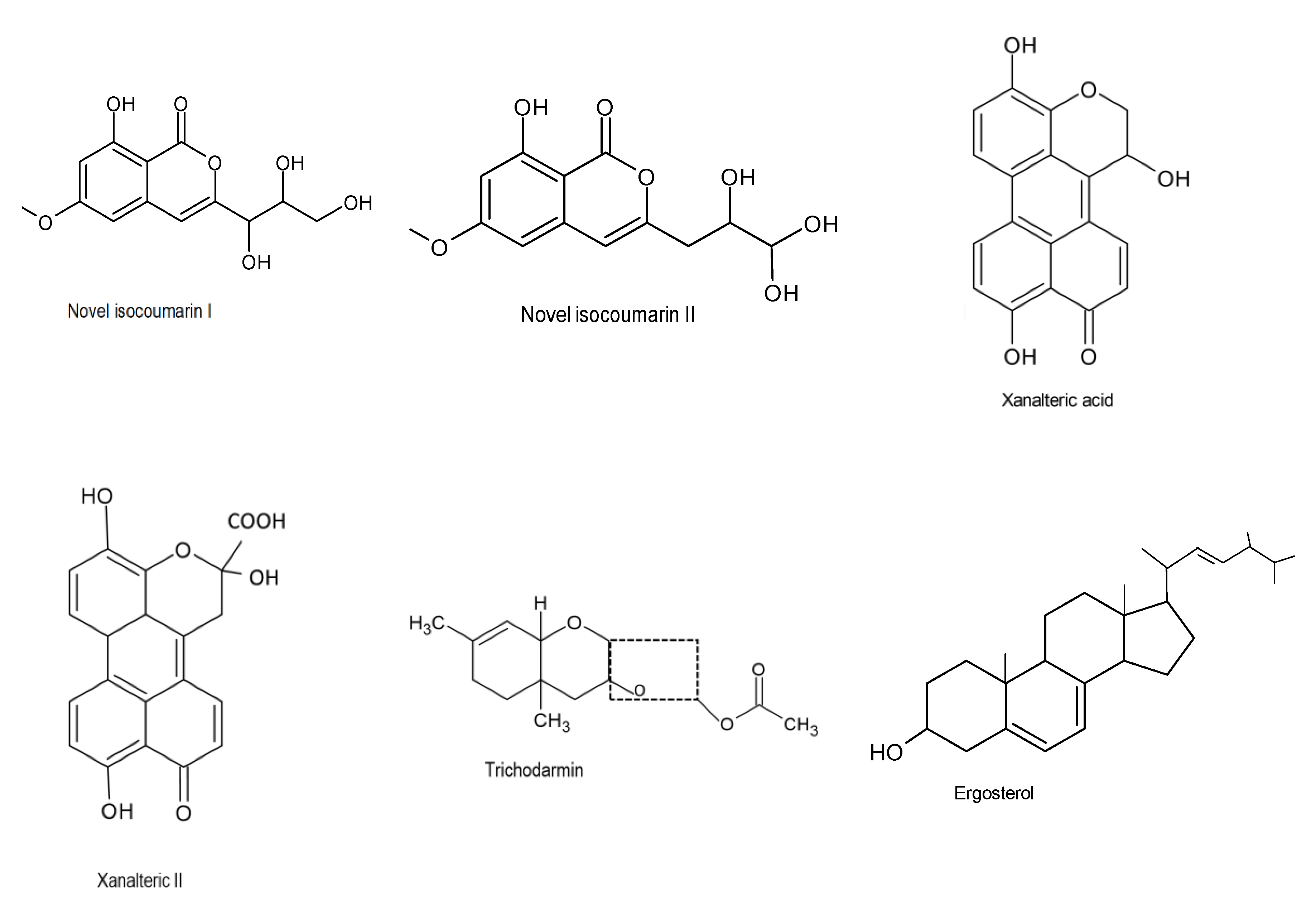{kind=link}
{kind=link}
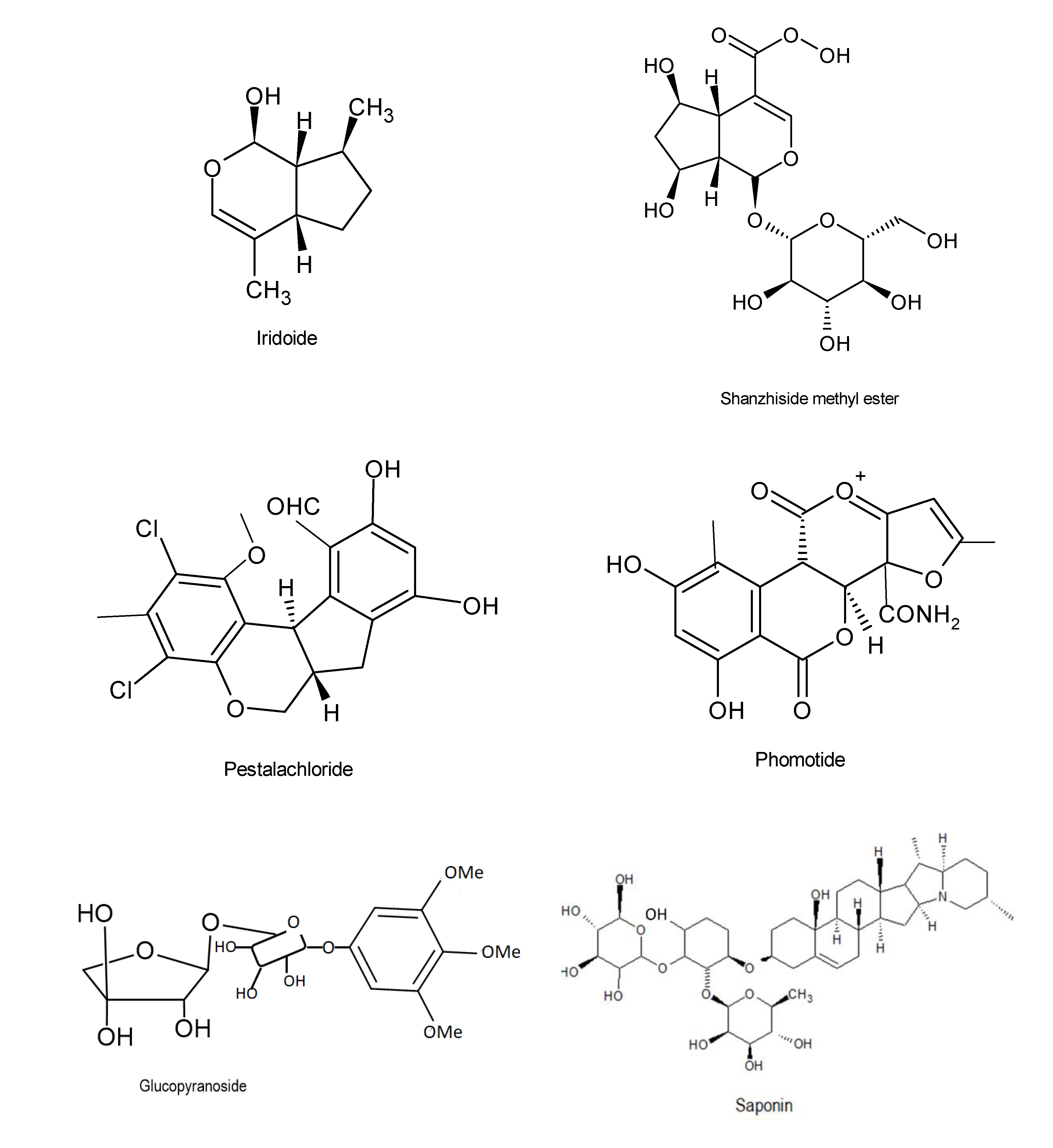{kind=link}
{kind=link}
{kind=link}
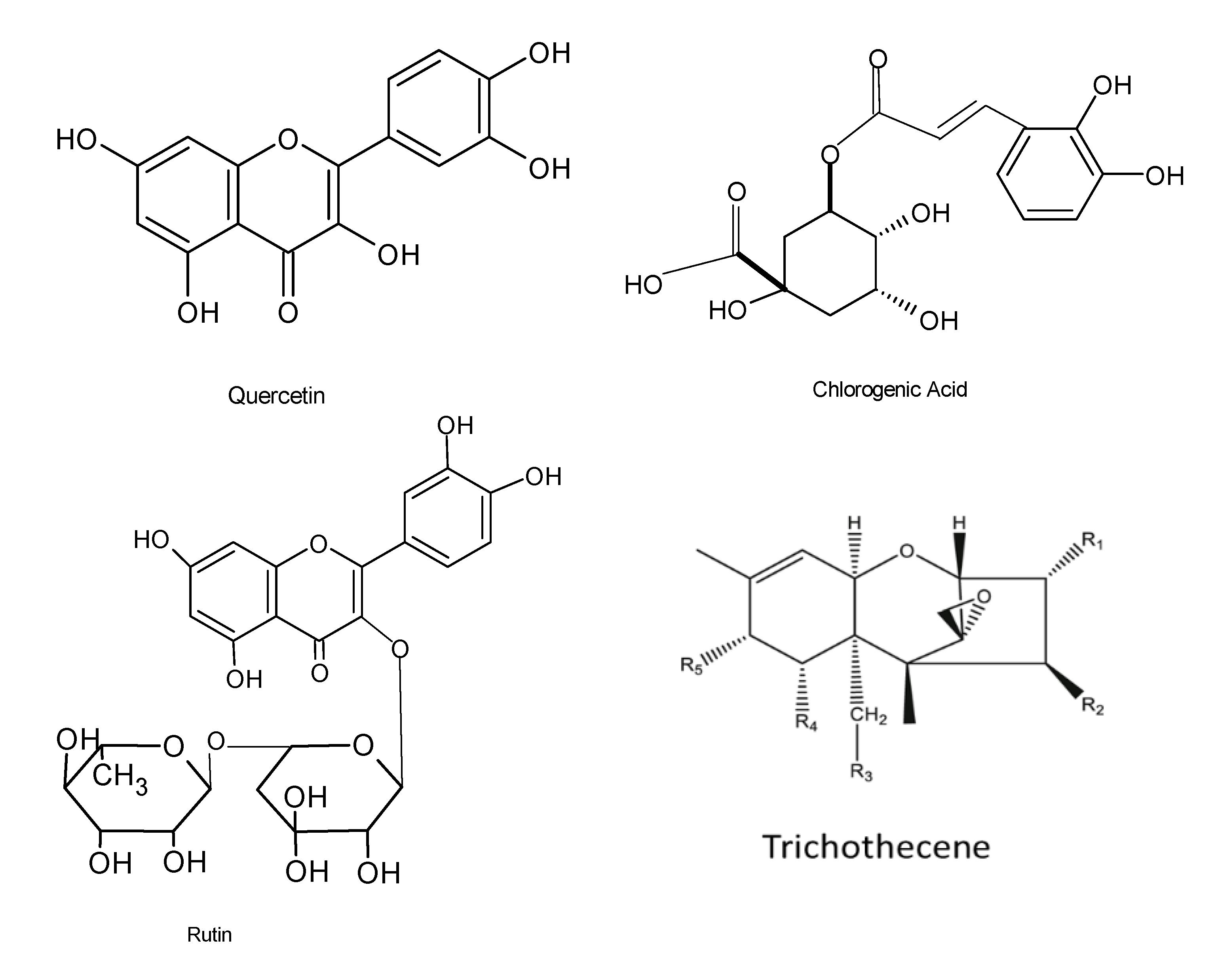{kind=link}
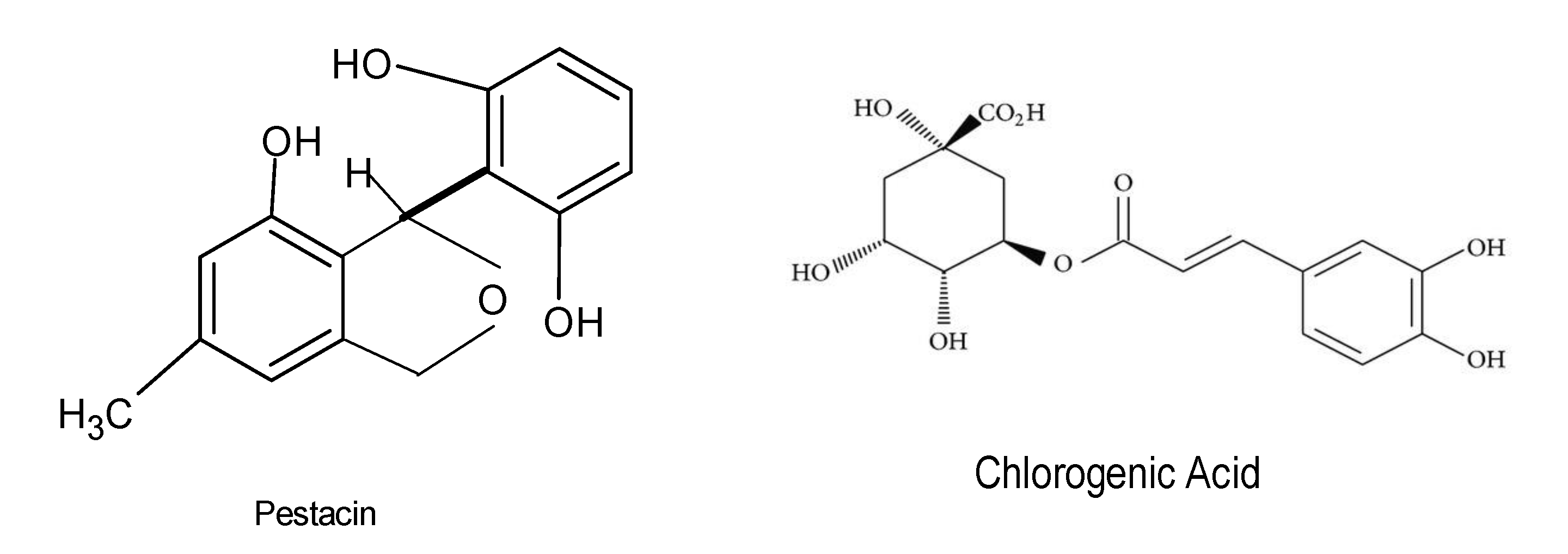{kind=link}
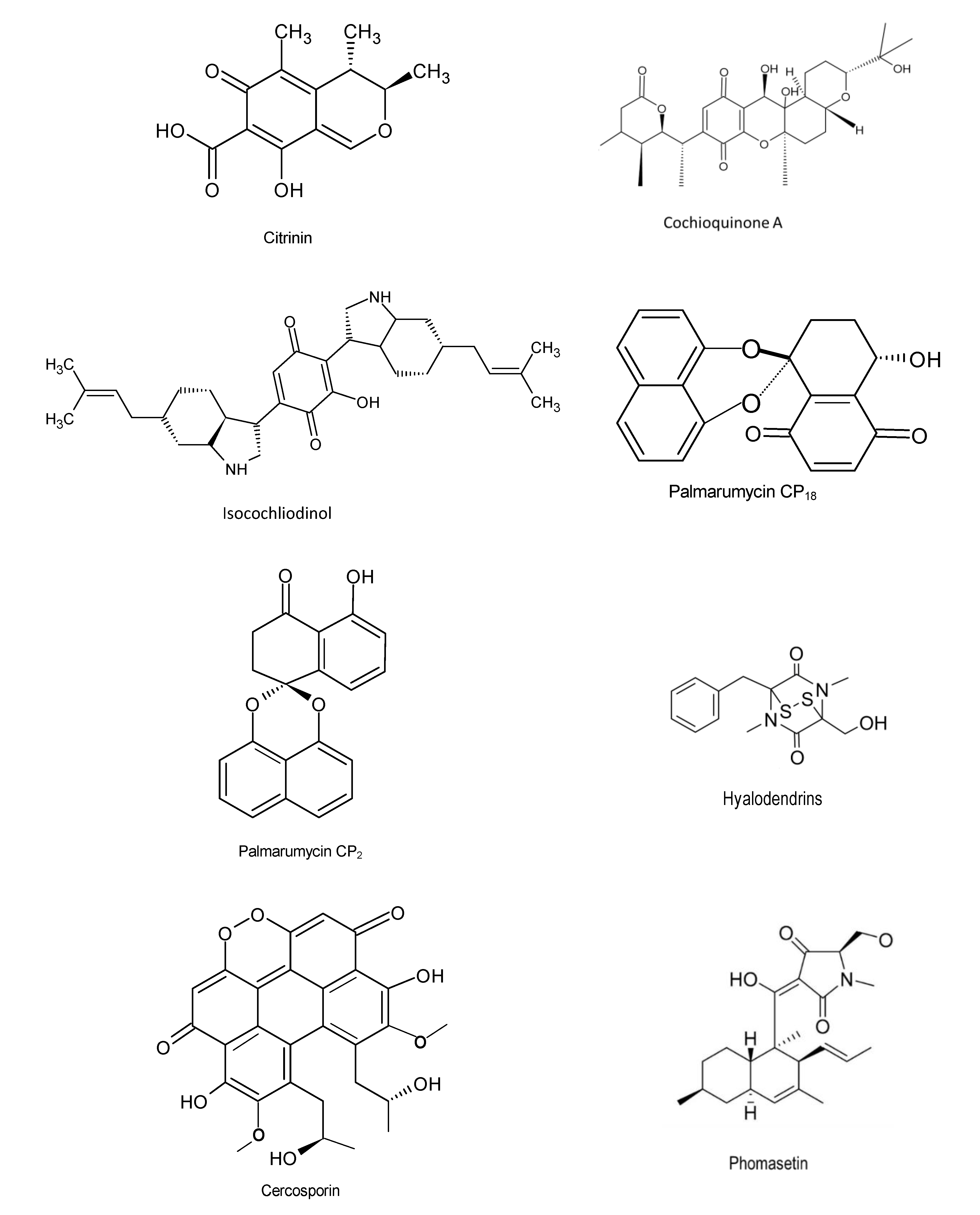{kind=link}
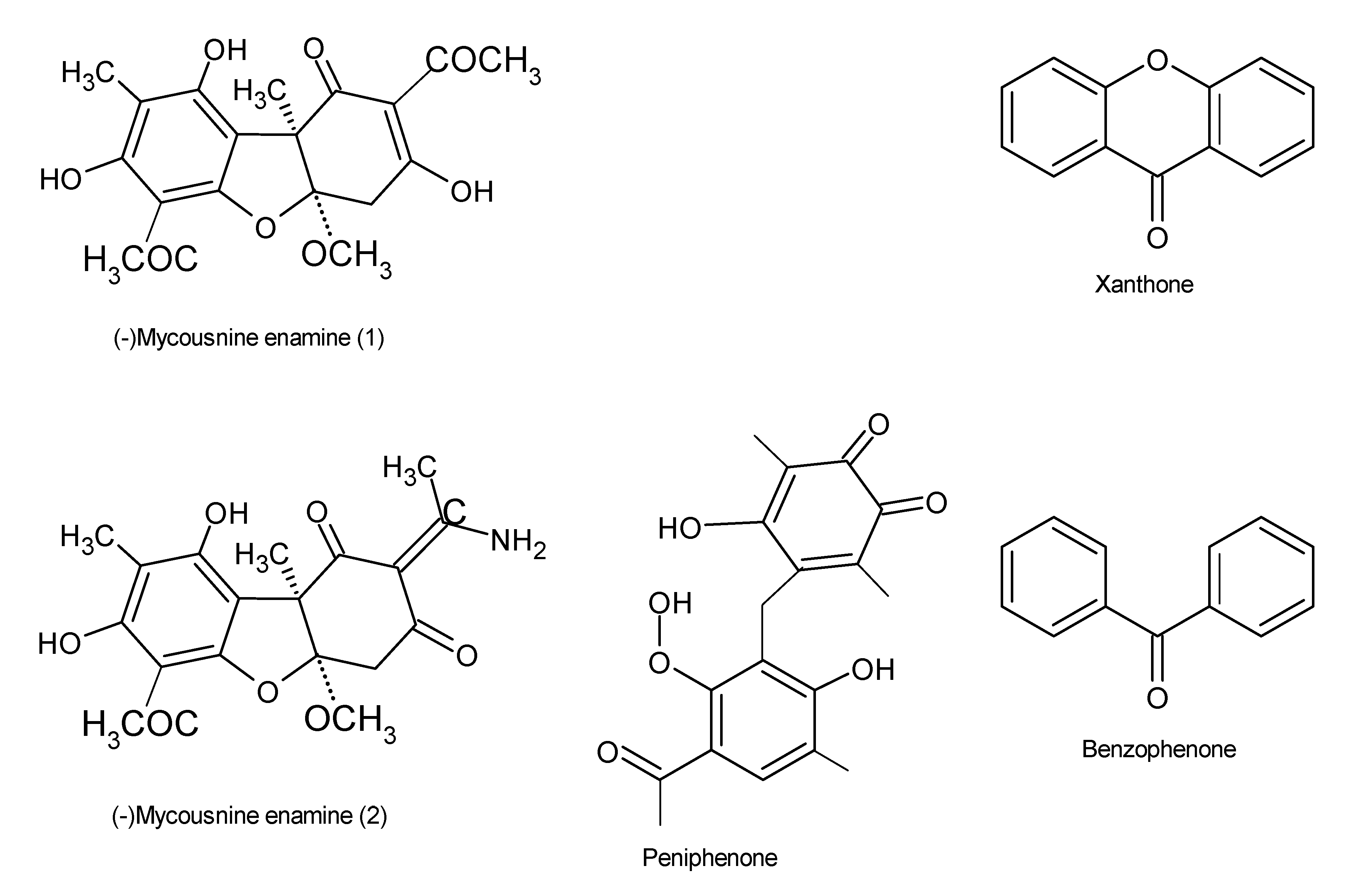{kind=link}
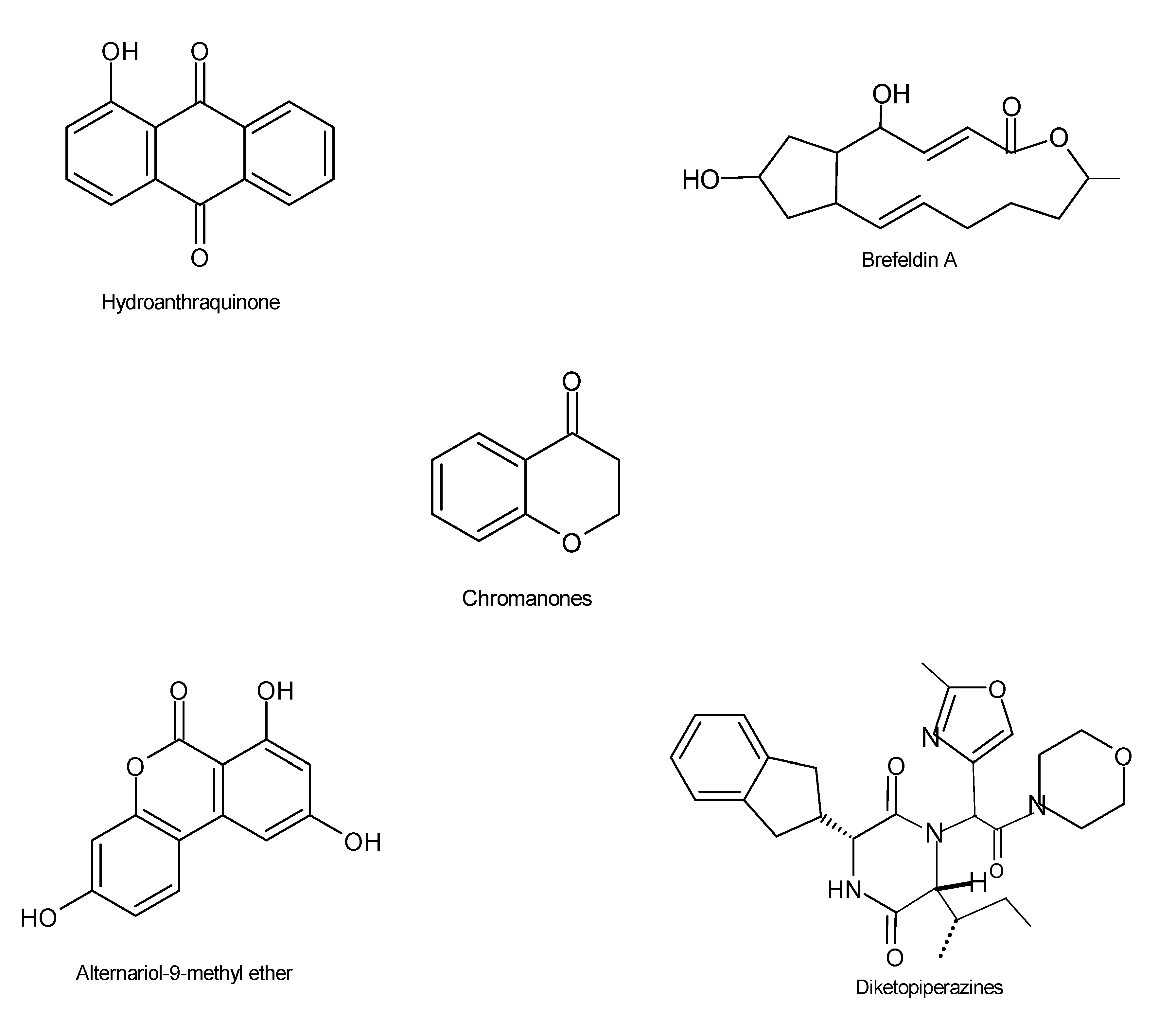{kind=link}
{kind=link}
Table 1.
Summarization of diseases and side effects of oxidative stress.
| OXIDATIVE STRESS | |
|---|---|
| Neurological | Multi-System Effects |
| Attention-deficit/hyperactivity disorder (ADHD) Alzheimer’s disease Anxiety and depression Asperger syndrome Autism Multiple sclerosis Parkinson’s disease | Diabetes Cancer Inflammation Fibromyalgia Lyme disease Chronic fatigue syndrome Metabolic syndrome Anxiety Hyperthyroidism Sleep apnea |
| Cardiovascular | Joints/Skin |
| Cardiovascular Disease Angina Pectoris Hypertension Atherosclerosis | Gout Dermatitis Rheumatoid arthritis Carpal tunnel syndrome |
| Gastrointestinal Disorder | Respiratory |
| Crohn’s Disease Gastroesophageal reflux disease (GERD) Gastric ulcers Celiac disease Functional dyspepsia | Chronic obstructive pulmonary disease (COPD) Asthma |
Table 2.
List of some bioactive compounds produced by endophytic fungi possessing biological activities.
Table 2.
List of some bioactive compounds produced by endophytic fungi possessing biological activities.
| Endophytic Fungi | Host Plant | Bioactive Compounds | Biological Properties | Activity Level | Ref. |
|---|---|---|---|---|---|
| Penicillium funiculosum Fes1711 and Trichoderma harzianum Fes1712 | Ficus elastica | Isocoumarin derivatives | Antibacterial activity | MIC = 32 μg/mL | [34] |
| Phomopsis sp. CFS42 | Cephalotaxus fortunei | Polyketides | Antifungal activity | MIC = 2.5 μg/mL | [44] |
| Chaetomium globosum | Ginkgo biloba | Azaphilone alkaloids | Anticancer activity | IC50 = 53.4 μM | [58] |
| Alternaria alternata AE1 | Azadirachta indica | Phenolics and flavonoids | Antioxidant properties | IC50 = 38 μg/mL | [28] |
| Mycosphaerella nawae ZJLQ129 | Smilax china | Amide derivative | Immunosuppressant activity | 30 and 300 nM | [98] |
| Phomopsis sp. CGMCC No. 5416 | Achyranthes bidentata | Chromanones | Antiviral activity | IC50 =32.5 μg/ ml | [111] |
| Gliocladium sp. MR41 | Culture collection | Polyols | Antitubercular properties | MIC = 3.13 µg/mL | [116] |
| Penicillium roqueforti and Trichoderma reesei | Solanum surattense | Ferulic acid, cinnamic acid, quercetin, and rutin | Antibacterial activity | MBC = 2.5 µg/mL | [118] |
| Lasiodiplodia pseudotheobromae PAK-7 and L. theobromae TN-R-3 | Theobroma cacao L. | dl-Mevalonic acid lactone, Methyl 6-O-[1-methylpropyl]-á-d-galactopyranoside | Antibacterial activity | MIC= 21 mm | [119] |
| Trichoderma asperellum T1 | Culture collection | 6-pentyl-2H-pyran-2-one (6-PP) | Antifungal and plant promoting properties | 61.31% Inhibition | [120] |
| Cladosporium cladosporioides | Zygophyllum mandavillei | 3-phenylpropionic acid, 5′-hydroxyasperentin | Antifungal activity | MIC = 15.62 μg/mL | [121] |
| Talaromyces purpureogenus | Grateloupia filicina | Talaromyolide K | Antiviral activity | 60.11% Inhibition | [122] |
| Aspergillus sp. SCSIO XWS02F40 | Callyspongia sp. | Asteltoxins | Antiviral activity | IC50 = 3.5 μg/mL | [123] |
| Diaporthe schini | Solanum americanum | 1,4-diaza-2,5-dioxo-3-isobutyl bicyclo[4.3.0]nonane and benzeneethanol | Antioxidant activity | DPPH radical = 96.62% | [124] |
| Botryosphaeria dothidea | Pampa and Atlantic Forest Plants | Hexahydropyrrolizin-3-one and (2-methylpropyl) ester | Antioxidant activity | IC50 = 0.206 mg/mL | [125] |
| Fusarium solani S-019 | Camptotheca acuminate | Camptothecin | Anticancer activity | 50 µg/mL | [126] |
| Alternaria alternata KT380662 | Passiflora incarnata L. | Flavone chrysin (5,7-dihydroxy flavone) | Anticancer activity | IC50 = 37.97 μg/mL | [127] |
| Diaporthe phaseolorum 92C | Combretum lanceolatum | 18-Des-hydroxy Cytochalasin | Antiparasitic activity | IC50 = 50 μg/mL | [124] |
| Phyllosticta capitalensi | Tibouchina granulosa | Brefeldin and heptelidic acid | Antiparasitic activity | IC50 = 50.13 μg/mL, | [89] |
| Fusarium solani | Glycyrrhiza glabra | Fusarubin, 3-O-methylfusarubin, and javanicin | Antitubercular activity | MIC = 8 μg/mL | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Manganyi, M.C.; Ateba, C.N. Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms 2020, 8, 1934. https://doi.org/10.3390/microorganisms8121934
AMA Style
Manganyi MC, Ateba CN. Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms. 2020; 8(12):1934. https://doi.org/10.3390/microorganisms8121934
Chicago/Turabian StyleManganyi, Madira Coutlyne, and Collins Njie Ateba. 2020. "Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications" Microorganisms 8, no. 12: 1934. https://doi.org/10.3390/microorganisms8121934
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.