Disrupting Bordetella Immunosuppression Reveals a Role for Eosinophils in Coordinating the Adaptive Immune Response in the Respiratory Tract

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Animal Experiments
2.3. Enzyme-Linked Immunosorbent Assays and Cytokine Assays
2.4. Flow Cytometry
2.5. Ethics Statement
2.6. Statistical Analysis
3. Results
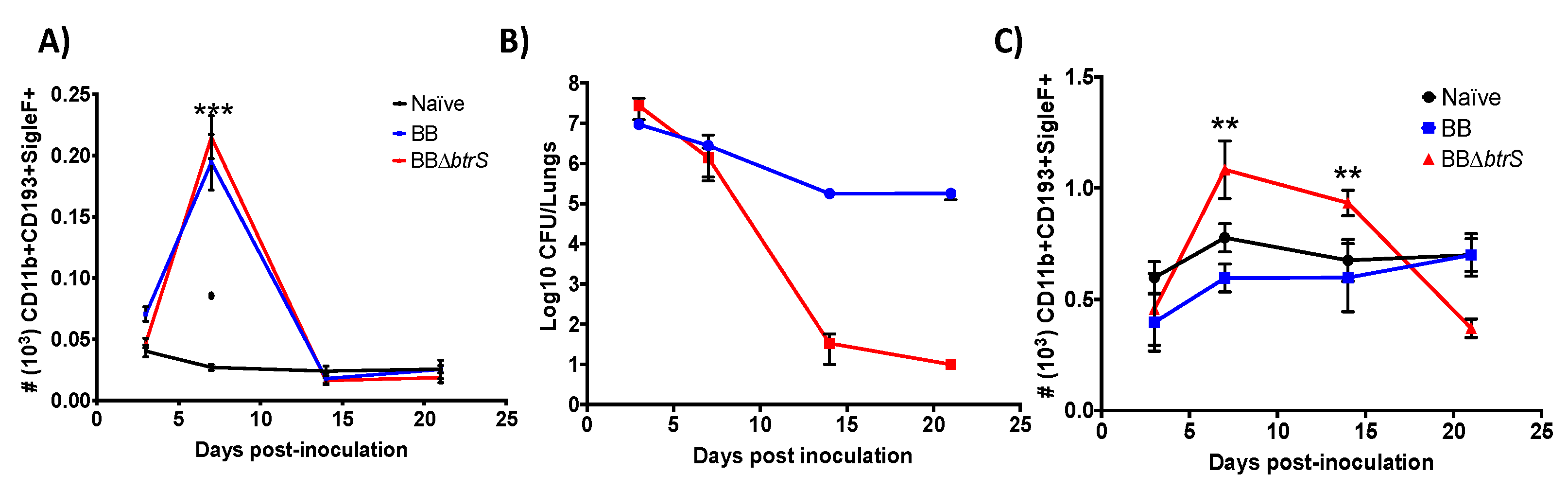
3.1. Splenic eosinophils Numbers Increase in Response to B. bronchiseptica
3.2. Bordetella bronchiseptica Blocks Eosinophil Recruitment to the Lungs
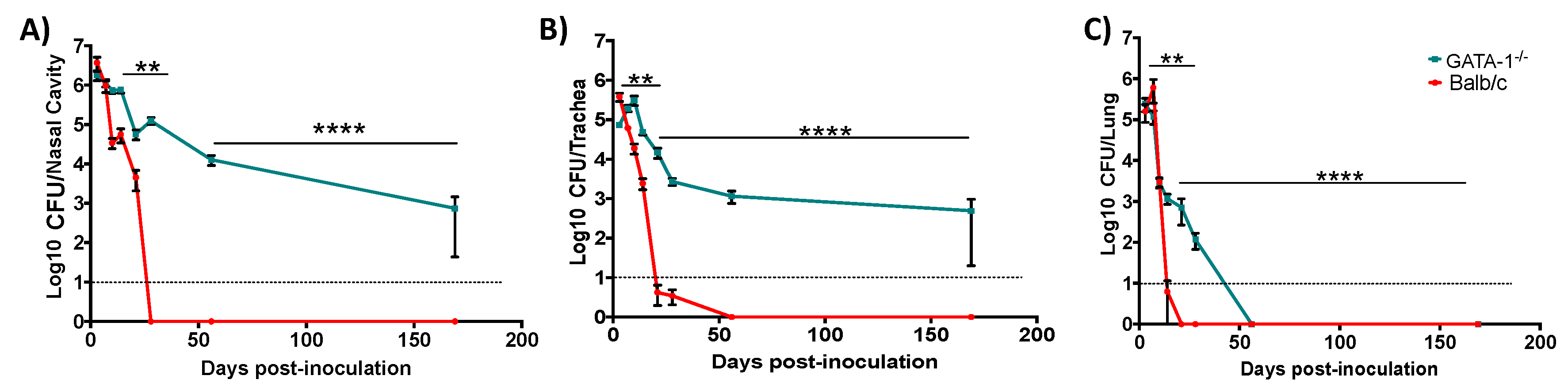
3.3. Eosinophil-Deficient Mice Fail to Clear Respiratory Infection
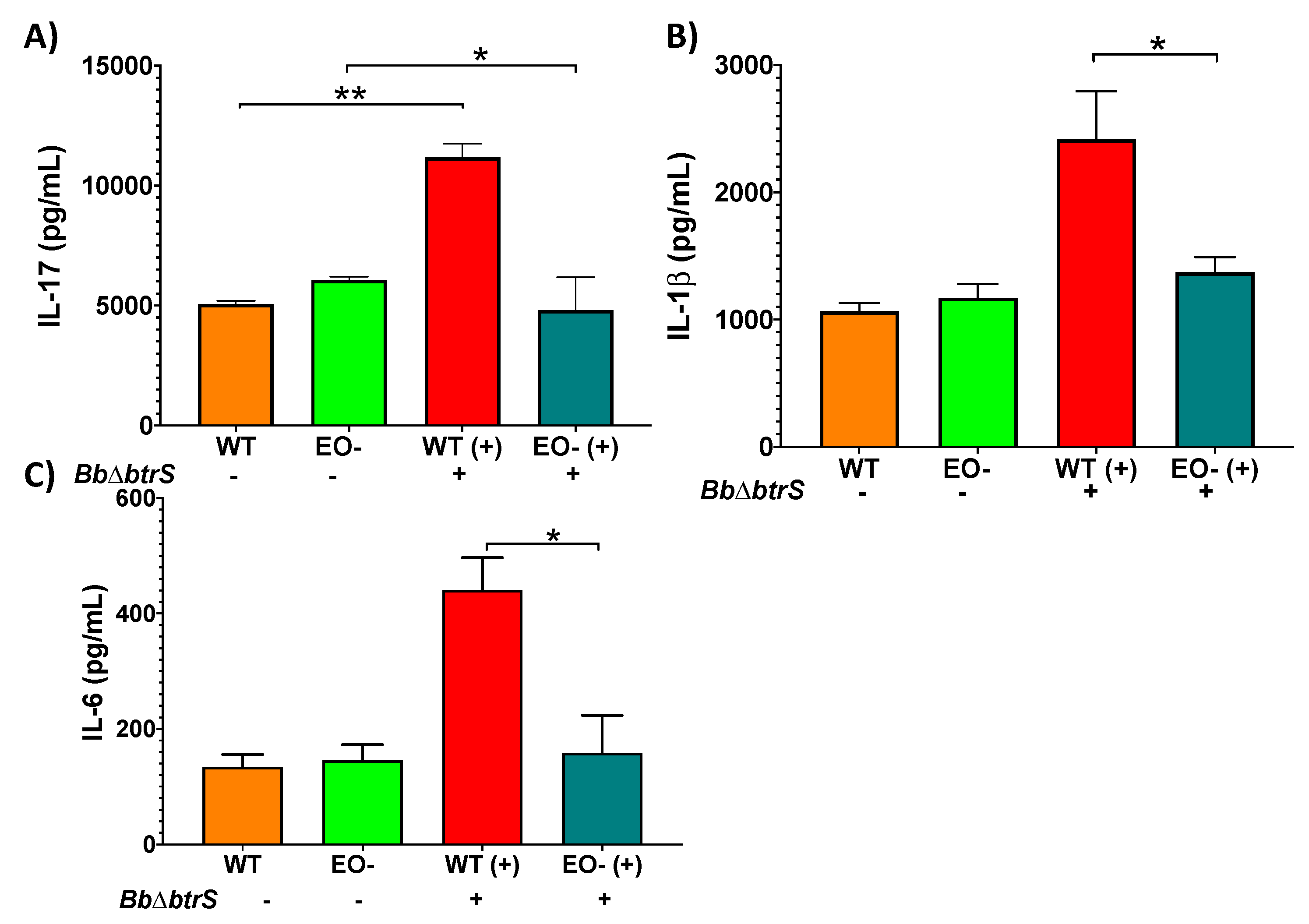
3.4. Eosinophils Contribute to Cytokine Secretion in Respiratory Mucosal Surfaces
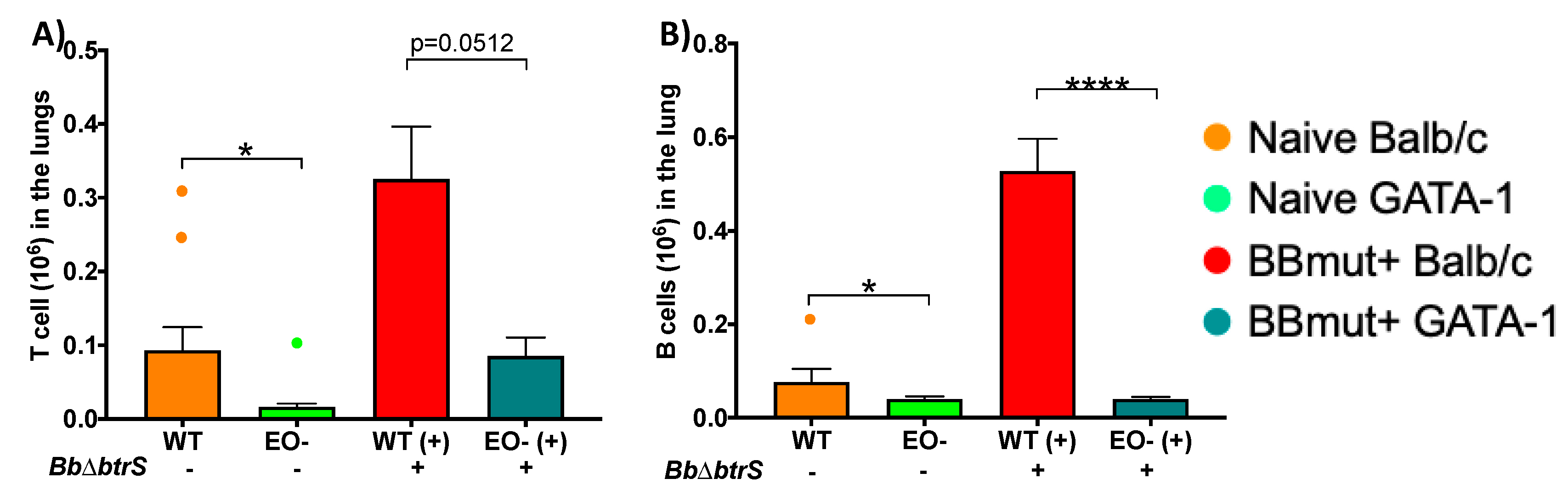
3.5. Eosinophils Mediate Lymphocyte Recruitment to the Lungs
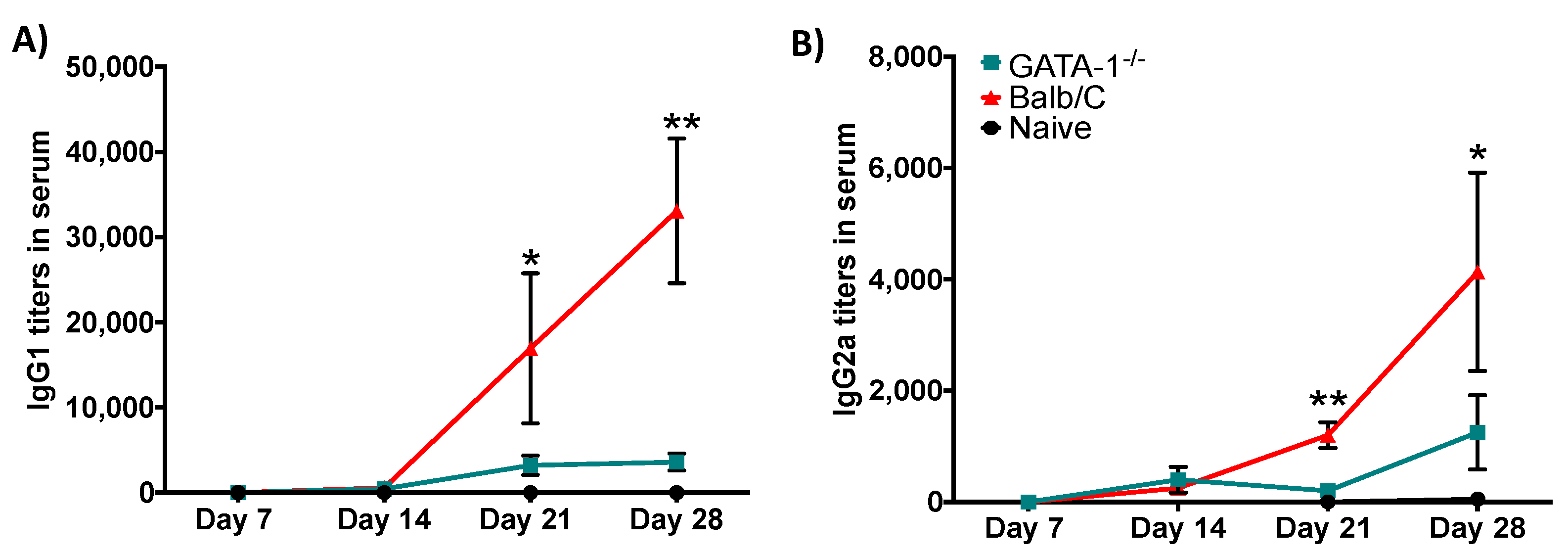
3.6. Eosinophils Are Required for an Efficient Antibody Response
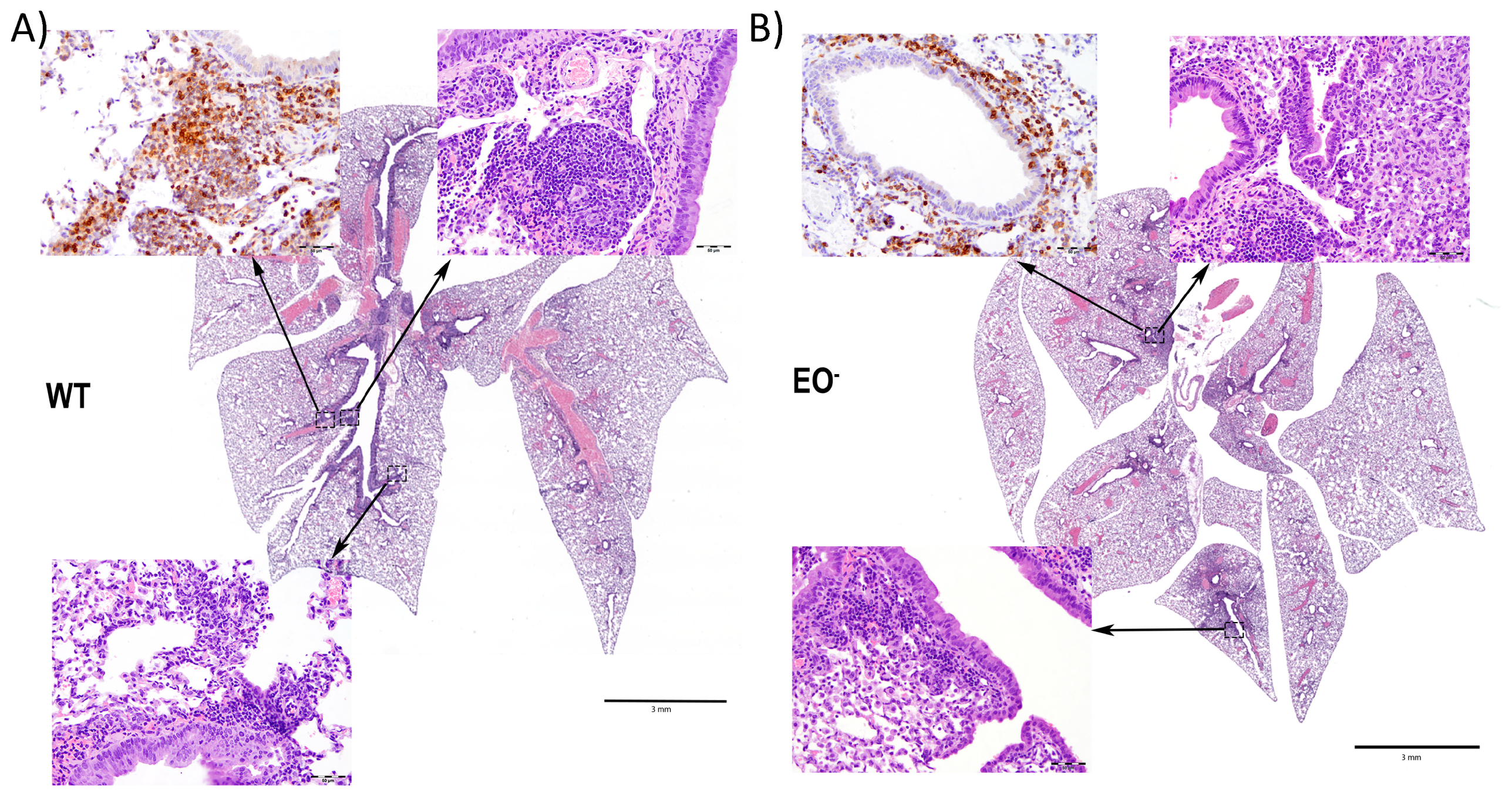
3.7. Eosinophils Contribute to an Organized Local Immune Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansen, K. Allergic and Psychical Factors in Asthma. Proc. R. Soc. Med. 1929, 22, 789–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Abrera, V.S.E. Further observations on cases of asthma and bronchitis associated with high eosinophilia and with mites in the sputum. Ind. Med. Gaz. 1946, 81, 414–417. [Google Scholar]
- Gundel, R.H.; Letts, L.G.; Gleich, G.J. Human eosinophil major basic protein induces airway constriction and airway hyperresponsiveness in primates. J. Clin. Investig. 1991, 87, 1470–1473. [Google Scholar] [CrossRef] [PubMed]
- Michel, F.B.; Bousquet, J.; Godard, P. Bronchial epithelium and asthma. Bull. Acad. Natl. Med. 1992, 176, 683–692. [Google Scholar]
- Boushey, H.A.; Fahy, J.V. Basic mechanisms of asthma. Environ. Health Perspect. 1995, 103 (Suppl. 6), 229–233. [Google Scholar] [CrossRef]
- Heymann, P.W.; Rakes, G.P.; Hogan, A.D.; Ingram, J.M.; Hoover, G.E.; Platts-Mills, T.A. Assessment of eosinophils, viruses and IgE antibody in wheezing infants and children. Int. Arch. Allergy Immunol. 1995, 107, 380–382. [Google Scholar] [CrossRef]
- Koller, D.Y.; Herouy, Y.; Götz, M.; Hagel, E.; Urbanek, R.; Eichler, I. Clinical value of monitoring eosinophil activity in asthma. Arch. Dis. Child. 1995, 73, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Thomas, L.H.; Warner, J.A. The eosinophil and its role in asthma. Gen. Pharmacol. 1996, 27, 593–597. [Google Scholar] [CrossRef]
- Boushey, H.A. Pathogenesis of asthma. Clin. Cornerstone 1998, 1, 1–8. [Google Scholar] [CrossRef]
- Katz, L.E.; Gleich, G.J.; Hartley, B.F.; Yancey, S.W.; Ortega, H.G. Blood eosinophil count is a useful biomarker to identify patients with severe eosinophilic asthma. Ann. Am. Thorac. Soc. 2014, 11, 531–536. [Google Scholar] [CrossRef]
- MANSMANN, J.A. The study of the eosinophile in allergic states. Am. J. Med. Technol. 1946, 12, 25–30. [Google Scholar] [PubMed]
- Weller, P.F.; Lim, K.; Wan, H.C.; Dvorak, A.M.; Wong, D.T.; Cruikshank, W.W.; Kornfeld, H.; Center, D.M. Role of the eosinophil in allergic reactions. Eur. Respir. J. Suppl. 1996, 22, 109s–115s. [Google Scholar] [PubMed]
- Piliponsky, A.M.; Pickholtz, D.; Gleich, G.J.; Levi-Schaffer, F. Human eosinophils induce histamine release from antigen-activated rat peritoneal mast cells: A possible role for mast cells in late-phase allergic reactions. J. Allergy Clin. Immunol. 2001, 107, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D.; Foster, P.S. Eosinophils: Changing perspectives in health and disease. Nat. Rev. Immunol. 2013, 13, 9–22. [Google Scholar] [CrossRef]
- Ueki, S.; Takeda, M. Allergic inflammatory diseases with luminal eosinophilia and extracellular trap cell death (etosis). Arerugi 2019, 68, 1126–1131. [Google Scholar] [CrossRef]
- Rothenberg, M.E.; Mishra, A.; Brandt, E.B.; Hogan, S.P. Gastrointestinal eosinophils. Immunol. Rev. 2001, 179, 139–155. [Google Scholar] [CrossRef]
- Hogan, S.P.; Rosenberg, H.F.; Moqbel, R.; Phipps, S.; Foster, P.S.; Lacy, P.; Kay, A.B.; Rothenberg, M.E. Eosinophils: Biological properties and role in health and disease. Clin. Exp. Allergy 2008, 38, 709–750. [Google Scholar] [CrossRef]
- Shamri, R.; Xenakis, J.J.; Spencer, L.A. Eosinophils in innate immunity: An evolving story. Cell Tissue Res. 2011, 343, 57–83. [Google Scholar] [CrossRef] [Green Version]
- Lacy, P.; Rosenberg, H.F.; Walsh, G.M. Eosinophil overview: Structure, biological properties, and key functions. Methods Mol. Biol 2014, 1178, 1–12. [Google Scholar] [CrossRef]
- Lee, J.J.; Jacobsen, E.A.; McGarry, M.P.; Schleimer, R.P.; Lee, N.A. Eosinophils in health and disease: The LIAR hypothesis. Clin. Exp. Allergy 2010, 40, 563–575. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K. Mechanism of parasite killing by eosinophils in parasitic infections. Nihon Rinsho 1993, 51, 657–663. [Google Scholar]
- Grezel, D.; Capron, M.; Grzych, J.M.; Fontaine, J.; Lecocq, J.P.; Capron, A. Protective immunity induced in rat schistosomiasis by a single dose of the Sm28GST recombinant antigen: Effector mechanisms involving IgE and IgA antibodies. Eur. J. Immunol. 1993, 23, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, K.; Sugaya, H.; Ishida, K. The role of eosinophils in Angiostrongylus cantonensis infection. Parasitol Today 1994, 10, 231–233. [Google Scholar] [CrossRef]
- Padigel, U.M.; Lee, J.J.; Nolan, T.J.; Schad, G.A.; Abraham, D. Eosinophils can function as antigen-presenting cells to induce primary and secondary immune responses to Strongyloides stercoralis. Infect. Immun. 2006, 74, 3232–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padigel, U.M.; Hess, J.A.; Lee, J.J.; Lok, J.B.; Nolan, T.J.; Schad, G.A.; Abraham, D. Eosinophils act as antigen-presenting cells to induce immunity to Strongyloides stercoralis in mice. J. Infect. Dis. 2007, 196, 1844–1851. [Google Scholar] [CrossRef] [Green Version]
- Nfon, C.K.; Makepeace, B.L.; Njongmeta, L.M.; Tanya, V.N.; Bain, O.; Trees, A.J. Eosinophils contribute to killing of adult Onchocerca ochengi within onchocercomata following elimination of Wolbachia. Microbes Infect. 2006, 8, 2698–2705. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Jimenez-Saiz, R.; Verschoor, C.P.; Walker, T.D.; Goncharova, S.; Llop-Guevara, A.; Shen, P.; Gordon, M.E.; Barra, N.G.; Bassett, J.D.; et al. Indigenous enteric eosinophils control DCs to initiate a primary Th2 immune response in vivo. J. Exp. Med. 2014, 211, 1657–1672. [Google Scholar] [CrossRef] [PubMed]
- Chu, V.T.; Beller, A.; Rausch, S.; Strandmark, J.; Zänker, M.; Arbach, O.; Kruglov, A.; Berek, C. Eosinophils promote generation and maintenance of immunoglobulin-A-expressing plasma cells and contribute to gut immune homeostasis. Immunity 2014, 40, 582–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, H.F.; Masterson, J.C.; Furuta, G.T. Eosinophils, probiotics, and the microbiome. J. Leukoc. Biol. 2016, 100, 881–888. [Google Scholar] [CrossRef]
- Mishra, A.; Hogan, S.P.; Lee, J.J.; Foster, P.S.; Rothenberg, M.E. Fundamental signals that regulate eosinophil homing to the gastrointestinal tract. J. Clin. Investig. 1999, 103, 1719–1727. [Google Scholar] [CrossRef] [Green Version]
- Rothenberg, M.E. Gastrointestinal eosinophils. Allergy 2001, 56 (Suppl. 67), 21–22. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.P.; Waddell, A.; Fulkerson, P.C. Eosinophils in infection and intestinal immunity. Curr. Opin. Gastroenterol. 2013, 29, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Femling, J.K.; Cherny, V.V.; Morgan, D.; Rada, B.; Davis, A.P.; Czirják, G.; Enyedi, P.; England, S.K.; Moreland, J.G.; Ligeti, E.; et al. The antibacterial activity of human neutrophils and eosinophils requires proton channels but not BK channels. J. Gen. Physiol. 2006, 127, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Linch, S.N.; Kelly, A.M.; Danielson, E.T.; Pero, R.; Lee, J.J.; Gold, J.A. Mouse eosinophils possess potent antibacterial properties in vivo. Infect. Immun. 2009, 77, 4976–4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatano, Y.; Taniuchi, S.; Masuda, M.; Tsuji, S.; Ito, T.; Hasui, M.; Kobayashi, Y.; Kaneko, K. Phagocytosis of heat-killed Staphylococcus aureus by eosinophils: Comparison with neutrophils. APMIS 2009, 117, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.J. Microbicidal activity of human eosinophils. J. Reticuloendothel Soc. 1972, 12, 332–339. [Google Scholar] [PubMed]
- Cline, M.J.; Hanifin, J.; Lehrer, R.I. Phagocytosis by human eosinophils. Blood 1968, 32, 922–934. [Google Scholar] [CrossRef]
- Cohen, S.G.; Sapp, T.M. Phagocytosis of bacteria by eosinophils in infectious-related asthma. J. Allergy 1969, 44, 113–117. [Google Scholar] [CrossRef]
- Cohen, S.G.; Sapp, T.M.; Chiampi, P.N. Eosinophil leukocyte responses and hypersensitivity reactions in the Bordetella pertussis-treated mouse. J. Allergy 1970, 46, 205–215. [Google Scholar] [CrossRef]
- Baehner, R.L.; Johnston, R.B. Metabolic and bactericidal activities of human eosinophils. Br. J. Haematol. 1971, 20, 277–285. [Google Scholar] [CrossRef]
- Bujak, J.S.; Root, R.K. The role of peroxidase in the bactericidal activity of human blood eosinophils. Blood 1974, 43, 727–736. [Google Scholar] [CrossRef]
- Leyva Covian, F. Eosinophils. III. Phagocytosis. Rev. Clin. Esp. 1977, 144, 167–171. [Google Scholar] [PubMed]
- Baggiolini, M. Phagocytes use oxygen to kill bacteria. Experientia 1984, 40, 906–909. [Google Scholar] [CrossRef] [PubMed]
- Kernen, P.; Wymann, M.P.; von Tscharner, V.; Deranleau, D.A.; Tai, P.C.; Spry, C.J.; Dahinden, C.A.; Baggiolini, M. Shape changes, exocytosis, and cytosolic free calcium changes in stimulated human eosinophils. J. Clin. Investig. 1991, 87, 2012–2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueki, S.; Konno, Y.; Takeda, M.; Moritoki, Y.; Hirokawa, M.; Matsuwaki, Y.; Honda, K.; Ohta, N.; Yamamoto, S.; Takagi, Y.; et al. Eosinophil extracellular trap cell death-derived DNA traps: Their presence in secretions and functional attributes. J. Allergy Clin. Immunol 2016, 137, 258–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gevaert, E.; Zhang, N.; Krysko, O.; Lan, F.; Holtappels, G.; De Ruyck, N.; Nauwynck, H.; Yousefi, S.; Simon, H.U.; Bachert, C. Extracellular eosinophilic traps in association with Staphylococcus aureus at the site of epithelial barrier defects in patients with severe airway inflammation. J. Allergy Clin. Immunol. 2017, 139, 1849–1860.e1846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousefi, S.; Simon, D.; Stojkov, D.; Karsonova, A.; Karaulov, A.; Simon, H.U. In vivo evidence for extracellular DNA trap formation. Cell Death Dis. 2020, 11, 300. [Google Scholar] [CrossRef]
- Samarasinghe, A.E.; Melo, R.C.; Duan, S.; LeMessurier, K.S.; Liedmann, S.; Surman, S.L.; Lee, J.J.; Hurwitz, J.L.; Thomas, P.G.; McCullers, J.A. Eosinophils Promote Antiviral Immunity in Mice Infected with Influenza A Virus. J. Immunol. 2017, 198, 3214–3226. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, P.J. Immunity and immunopathology to respiratory syncytial virus. The mouse model. Am. J. Respir. Crit. Care Med. 1995, 152, S59–S62. [Google Scholar] [CrossRef]
- Ehlenfield, D.R.; Cameron, K.; Welliver, R.C. Eosinophilia at the time of respiratory syncytial virus bronchiolitis predicts childhood reactive airway disease. Pediatrics 2000, 105, 79–83. [Google Scholar] [CrossRef]
- Rosenberg, H.F.; Dyer, K.D.; Domachowske, J.B. Respiratory viruses and eosinophils: Exploring the connections. Antivir. Res. 2009, 83, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jesenak, M.; Schwarze, J. Lung Eosinophils—A Novel ‘Virus Sink’ that is Defective in Asthma? Allergy 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabogal Piñeros, Y.S.; Bal, S.M.; Dijkhuis, A.; Majoor, C.J.; Dierdorp, B.S.; Dekker, T.; Hoefsmit, E.P.; Bonta, P.I.; Picavet, D.; van der Wel, N.N.; et al. Eosinophils capture viruses, a capacity that is defective in asthma. Allergy 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Zou, Y.; Liu, J.; Jin, X.; Ma, C.; Li, J.; Guo, R.; Huang, B. Analysis of the predicting factors of recurrent wheezing in infants. Ital. J. Pediatr. 2019, 45, 19. [Google Scholar] [CrossRef]
- Gestal, M.C.; Howard, L.K.; Dewan, K.; Johnson, H.M.; Barbier, M.; Bryant, C.; Soumana, I.H.; Rivera, I.; Linz, B.; Blas-Machado, U.; et al. Enhancement of immune response against Bordetella spp. by disrupting immunomodulation. Sci. Rep. 2019, 9, 20261. [Google Scholar] [CrossRef] [Green Version]
- McGuirk, P.; McCann, C.; Mills, K.H. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic cells: A novel strategy for evasion of protective T helper type 1 responses by Bordetella pertussis. J. Exp. Med. 2002, 195, 221–231. [Google Scholar] [CrossRef]
- Andreasen, C.; Powell, D.A.; Carbonetti, N.H. Pertussis toxin stimulates IL-17 production in response to Bordetella pertussis infection in mice. PLoS ONE 2009, 4, e7079. [Google Scholar] [CrossRef]
- Skerry, C.M.; Mahon, B.P. A live, attenuated Bordetella pertussis vaccine provides long-term protection against virulent challenge in a murine model. Clin. Vaccine Immunol. 2011, 18, 187–193. [Google Scholar] [CrossRef]
- Zhang, X.; Hester, S.E.; Kennett, M.J.; Karanikas, A.T.; Bendor, L.; Place, D.E.; Harvill, E.T. Interleukin-1 receptor signaling is required to overcome the effects of pertussis toxin and for efficient infection- or vaccination-induced immunity against Bordetella pertussis. Infect. Immun. 2011, 79, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Warfel, J.M.; Merkel, T.J. Bordetella pertussis infection induces a mucosal IL-17 response and long-lived Th17 and Th1 immune memory cells in nonhuman primates. Mucosal Immunol. 2013, 6, 787–796. [Google Scholar] [CrossRef]
- Place, D.E.; Muse, S.J.; Kirimanjeswara, G.S.; Harvill, E.T. Caspase-1-independent interleukin-1β is required for clearance of Bordetella pertussis infections and whole-cell vaccine-mediated immunity. PLoS ONE 2014, 9, e107188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Silva Antunes, R.; Babor, M.; Carpenter, C.; Khalil, N.; Cortese, M.; Mentzer, A.J.; Seumois, G.; Petro, C.D.; Purcell, L.A.; Vijayanand, P.; et al. Th1/Th17 polarization persists following whole-cell pertussis vaccination despite repeated acellular boosters. J. Clin. Investig. 2018, 128, 3853–3865. [Google Scholar] [CrossRef] [Green Version]
- Solans, L.; Debrie, A.S.; Borkner, L.; Aguiló, N.; Thiriard, A.; Coutte, L.; Uranga, S.; Trottein, F.; Martín, C.; Mills, K.H.G.; et al. IL-17-dependent SIgA-mediated protection against nasal Bordetella pertussis infection by live attenuated BPZE1 vaccine. Mucosal Immunol. 2018, 11, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.C.; Wilk, M.M.; Misiak, A.; Borkner, L.; Murphy, D.; Mills, K.H.G. Sustained protective immunity against Bordetella pertussis nasal colonization by intranasal immunization with a vaccine-adjuvant combination that induces IL-17-secreting T. Mucosal Immunol. 2018, 11, 1763–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gestal, M.C.; Rivera, I.; Howard, L.K.; Dewan, K.K.; Soumana, I.H.; Dedloff, M.; Nicholson, T.L.; Linz, B.; Harvill, E.T. Blood or Serum Exposure Induce Global Transcriptional Changes, Altered Antigenic Profile, and Increased Cytotoxicity by Classical Bordetellae. Front. Microbiol. 2018, 9, 1969. [Google Scholar] [CrossRef]
- Yu, C.; Cantor, A.B.; Yang, H.; Browne, C.; Wells, R.A.; Fujiwara, Y.; Orkin, S.H. Targeted deletion of a high-affinity GATA-binding site in the GATA-1 promoter leads to selective loss of the eosinophil lineage in vivo. J. Exp. Med. 2002, 195, 1387–1395. [Google Scholar] [CrossRef] [Green Version]
- Blas-Machado, U.; Saliki, J.T.; Sánchez, S.; Brown, C.C.; Zhang, J.; Keys, D.; Woolums, A.; Harvey, S.B. Pathogenesis of a bovine enterovirus-1 isolate in experimentally infected calves. Vet. Pathol. 2011, 48, 1075–1084. [Google Scholar] [CrossRef]
- Wilk, M.M.; Misiak, A.; McManus, R.M.; Allen, A.C.; Lynch, M.A.; Mills, K.H.G. Lung CD4 Tissue-Resident Memory T Cells Mediate Adaptive Immunity Induced by Previous Infection of Mice with. J. Immunol. 2017, 199, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Akdis, M.; Andrä, I.; Annunziato, F.; Bacher, P.; Barnaba, V.; Battistini, L.; Bauer, W.M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies. Eur. J. Immunol. 2017, 47, 1584–1797. [Google Scholar] [CrossRef] [Green Version]
- Halim, T.Y.F.; Takei, F. Current Protocols in Immunology; Wiley Online library: Hoboken, NJ, USA, 2014; Volume 3.25, p. 13. [Google Scholar]
- Arnold, I.C.; Artola-Borán, M.; Tallón de Lara, P.; Kyburz, A.; Taube, C.; Ottemann, K.; van den Broek, M.; Yousefi, S.; Simon, H.U.; Müller, A. Eosinophils suppress Th1 responses and restrict bacterially induced gastrointestinal inflammation. J. Exp. Med. 2018, 215, 2055–2072. [Google Scholar] [CrossRef]
- Kavanagh, H.; Noone, C.; Cahill, E.; English, K.; Locht, C.; Mahon, B.P. Attenuated Bordetella pertussis vaccine strain BPZE1 modulates allergen-induced immunity and prevents allergic pulmonary pathology in a murine model. Clin. Exp. Allergy 2010, 40, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, T.; Dillon, M.B.; da Silva Antunes, R.; Paul, S.; Peters, B.; Crotty, S.; Lindestam Arlehamn, C.S.; Sette, A. Th1 versus Th2 T cell polarization by whole-cell and acellular childhood pertussis vaccines persists upon re-immunization in adolescence and adulthood. Cell Immunol. 2016, 304–305, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misiak, A.; Wilk, M.M.; Raverdeau, M.; Mills, K.H. IL-17-Producing Innate and Pathogen-Specific Tissue Resident Memory γδ T Cells Expand in the Lungs of Bordetella pertussis-Infected Mice. J. Immunol. 2017, 198, 363–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, T.W.; Doyle, A.D.; Lee, J.J.; Jelinek, D.F. Eosinophils regulate peripheral B cell numbers in both mice and humans. J. Immunol. 2014, 192, 3548–3558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, J.P.; Kiani, A.; Bozza, P.T.; Rao, A. Regulation of allergic inflammation and eosinophil recruitment in mice lacking the transcription factor NFAT1: Role of interleukin-4 (IL-4) and IL-5. Blood 1998, 91, 2223–2230. [Google Scholar] [CrossRef]
- Berek, C. Eosinophils: Important players in humoral immunity. Clin. Exp. Immunol. 2016, 183, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Ignacio, A.; McCoy, K.D.; Harris, N.L. The emerging roles of eosinophils in mucosal homeostasis. Mucosal Immunol. 2020. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.J.; Matossian, K.; Resnick, M.B.; Beil, W.J.; Wong, D.T.; Gordon, J.R.; Dvorak, A.M.; Weller, P.F.; Galli, S.J. Human eosinophils can express the cytokines tumor necrosis factor-alpha and macrophage inflammatory protein-1 alpha. J. Clin. Investig. 1993, 91, 2673–2684. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Yang, T.; Wang, J.; Zhao, T.; Wang, L.; Kang, Y.; Cheng, C.; Fan, Y. Elevated Plasma Chemokines for Eosinophils in Neuromyelitis Optica Spectrum Disorders during Remission. Front. Neurol. 2018, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Wong, C.K.; Lam, C.W. Activation of eosinophils by IL-12 family cytokine IL-27: Implications of the pleiotropic roles of IL-27 in allergic responses. Immunobiology 2011, 216, 54–65. [Google Scholar] [CrossRef]
- Legrand, F.; Driss, V.; Woerly, G.; Loiseau, S.; Hermann, E.; Fournié, J.J.; Héliot, L.; Mattot, V.; Soncin, F.; Gougeon, M.L.; et al. A functional gammadeltaTCR/CD3 complex distinct from gammadeltaT cells is expressed by human eosinophils. PLoS ONE 2009, 4, e5926. [Google Scholar] [CrossRef]
- Spencer, L.A.; Szela, C.T.; Perez, S.A.; Kirchhoffer, C.L.; Neves, J.S.; Radke, A.L.; Weller, P.F. Human eosinophils constitutively express multiple Th1, Th2, and immunoregulatory cytokines that are secreted rapidly and differentially. J. Leukoc. Biol. 2009, 85, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gounni, A.S.; Nutku, E.; Koussih, L.; Aris, F.; Louahed, J.; Levitt, R.C.; Nicolaides, N.C.; Hamid, Q. IL-9 expression by human eosinophils: Regulation by IL-1beta and TNF-alpha. J. Allergy Clin. Immunol. 2000, 106, 460–466. [Google Scholar] [CrossRef]
- Yoon, J.; Um, H.N.; Jang, J.; Bae, Y.A.; Park, W.J.; Kim, H.J.; Yoon, M.S.; Chung, I.Y.; Jung, Y. Eosinophil Activation by Toll-Like Receptor 4 Ligands Regulates Macrophage Polarization. Front. Cell Dev. Biol. 2019, 7, 329. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Okugawa, S.; Ota, Y.; Yamaguchi, M.; Tomizawa, H.; Matsushima, K.; Ohta, K.; Yamamoto, K.; Hirai, K. Expression and function of Toll-like receptors in eosinophils: Activation by Toll-like receptor 7 ligand. J. Immunol. 2003, 171, 3977–3982. [Google Scholar] [CrossRef] [Green Version]
- Mulder, D.J.; Lobo, D.; Mak, N.; Justinich, C.J. Expression of toll-like receptors 2 and 3 on esophageal epithelial cell lines and on eosinophils during esophagitis. Dig. Dis. Sci. 2012, 57, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Cheung, P.F.; Ip, W.K.; Lam, C.W. Intracellular signaling mechanisms regulating toll-like receptor-mediated activation of eosinophils. Am. J. Respir. Cell Mol. Biol 2007, 37, 85–96. [Google Scholar] [CrossRef]
- Kvarnhammar, A.M.; Cardell, L.O. Pattern-recognition receptors in human eosinophils. Immunology 2012, 136, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Xenakis, J.J.; Howard, E.D.; Smith, K.M.; Olbrich, C.L.; Huang, Y.; Anketell, D.; Maldonado, S.; Cornwell, E.W.; Spencer, L.A. Resident intestinal eosinophils constitutively express antigen presentation markers and include two phenotypically distinct subsets of eosinophils. Immunology 2018, 154, 298–308. [Google Scholar] [CrossRef] [Green Version]
- Akuthota, P.; Wang, H.; Weller, P.F. Eosinophils as antigen-presenting cells in allergic upper airway disease. Curr. Opin. Allergy Clin. Immunol 2010, 10, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Gestal, M.C.; Whitesides, L.T.; Harvill, E.T. Integrated Signaling Pathways Mediate Bordetella Immunomodulation, Persistence, and Transmission. Trends Microbiol. 2019, 27, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Hester, S.E.; Lui, M.; Nicholson, T.; Nowacki, D.; Harvill, E.T. Identification of a CO2 responsive regulon in Bordetella. PLoS ONE 2012, 7, e47635. [Google Scholar] [CrossRef] [Green Version]
- Brickman, T.J.; Suhadolc, R.J.; Armstrong, S.K. Interspecies variations in Bordetella catecholamine receptor gene regulation and function. Infect. Immun. 2015, 83, 4639–4652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, S.K.; Brickman, T.J.; Suhadolc, R.J. Involvement of multiple distinct Bordetella receptor proteins in the utilization of iron liberated from transferrin by host catecholamine stress hormones. Mol. Microbiol. 2012, 84, 446–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gestal, M.; Jonhson, H.; Harvill, E. Immunomodulation as a Novel Strategy for Prevention and Treatment of Bordetella spp. Infections. Front. Front. Immunol. 2019, in press. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilione, M.R.; Harvill, E.T. The Bordetella bronchiseptica type III secretion system inhibits gamma interferon production that is required for efficient antibody-mediated bacterial clearance. Infect. Immun. 2006, 74, 1043–1049. [Google Scholar] [CrossRef] [Green Version]
- Siciliano, N.A.; Skinner, J.A.; Yuk, M.H. Bordetella bronchiseptica modulates macrophage phenotype leading to the inhibition of CD4+ T cell proliferation and the initiation of a Th17 immune response. J. Immunol. 2006, 177, 7131–7138. [Google Scholar] [CrossRef] [Green Version]
- Gorgojo, J.; Scharrig, E.; Gómez, R.M.; Harvill, E.T.; Rodríguez, M.E. Bordetella parapertussis Circumvents Neutrophil Extracellular Bactericidal Mechanisms. PLoS ONE 2017, 12, e0169936. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, C.; Eby, J.; Gray, M.; Heath Damron, F.; Melvin, J.; Cotter, P.; Hewlett, E. Bordetella adenylate cyclase toxin interacts with filamentous haemagglutinin to inhibit biofilm formation in vitro. Mol. Microbiol. 2017, 103, 214–228. [Google Scholar] [CrossRef] [Green Version]
- Gonyar, L.A.; Gray, M.C.; Christianson, G.J.; Mehrad, B.; Hewlett, E.L. Albumin, in the Presence of Calcium, Elicits a Massive Increase in Extracellular Bordetella Adenylate Cyclase Toxin. Infect. Immun. 2017, 85. [Google Scholar] [CrossRef] [Green Version]
- Freyberg, Z.; Harvill, E.T. Pathogen manipulation of host metabolism: A common strategy for immune evasion. PLoS Pathog. 2017, 13, e1006669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirimanjeswara, G.S.; Agosto, L.M.; Kennett, M.J.; Bjornstad, O.N.; Harvill, E.T. Pertussis toxin inhibits neutrophil recruitment to delay antibody-mediated clearance of Bordetella pertussis. J. Clin. Investig. 2005, 115, 3594–3601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauchi, S.; Locht, C. Non-specific Effects of Live Attenuated Pertussis Vaccine Against Heterologous Infectious and Inflammatory Diseases. Front. Immunol. 2018, 9, 2872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.Y.; Chi, S.; Sun, Y. Modulation of Toll-like signal path of allergic asthma by CpG-ODNs from Bordetella pertussis. Yao Xue Xue Bao 2011, 46, 285–292. [Google Scholar] [PubMed]
- Carbonetti, N.H. Immunomodulation in the pathogenesis of Bordetella pertussis infection and disease. Curr. Opin. Pharmacol. 2007, 7, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Ennis, D.P.; Cassidy, J.P.; Mahon, B.P. Whole-cell pertussis vaccine protects against Bordetella pertussis exacerbation of allergic asthma. Immunol. Lett. 2005, 97, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ennis, D.P.; Cassidy, J.P.; Mahon, B.P. Prior Bordetella pertussis infection modulates allergen priming and the severity of airway pathology in a murine model of allergic asthma. Clin. Exp. Allergy 2004, 34, 1488–1497. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Kwon, K.S.; Kim, D.K.; Choi, I.W.; Lee, H.K. Inhibition of murine allergic airway disease by Bordetella pertussis. Immunology 2004, 112, 624–630. [Google Scholar] [CrossRef]
- Dong, W.; Selgrade, M.K.; Gilmour, M.I. Systemic administration of Bordetella pertussis enhances pulmonary sensitization to house dust mite in juvenile rats. Toxicol. Sci. 2003, 72, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Donato, L.; Gaugler, C.; Weiss, L.; Krieger, P.; Debry, C. Chronic cough in children: Signs of serious disease and investigations. Arch. Pediatr. 2001, 8 (Suppl. 3), 638–644. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flow Cytometry Antibodies | ||||
|---|---|---|---|---|
| Fluorochrome | Target | Clone | Vendor | Ref |
| APC-Cy7 | GR1 | RB6–8C5 | Biolegend | 108–424 |
| PE | CD11b | M1/70 | Tonbo | 50–0112–U100 |
| PerCP | F4/80 | BM8 | Biolegend | 123,126 |
| APC | SiglecF | S17007L | Biolegend | 155,508 |
| BV510 | CD193 | CCR3 | BD Biosciences | 747,820 |
| PerCP | CD19 | 6D5 | Biolegend | 115,532 |
| PE | CD90.2 | 53–2.1 | Biolegend | 140,308 |
| AF488 | CD4 | GK1.5 | Biolegend | 100,423 |
| BV510 | CD8 | 53–6.7 | BD Biosciences | 563,068 |
| + | + | + | |
|---|---|---|---|
| Eosinophils | CD11b | CD193 | SiglecF |
| B cells | CD11b | CD19 | |
| CD4 | CD11b | CD90.2 | CD4 |
| CD8 | CD11b | CD90.2 | CD8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gestal, M.C.; Blas-Machado, U.; Johnson, H.M.; Rubin, L.N.; Dewan, K.K.; Bryant, C.; Tiemeyer, M.; Harvill, E.T. Disrupting Bordetella Immunosuppression Reveals a Role for Eosinophils in Coordinating the Adaptive Immune Response in the Respiratory Tract. Microorganisms 2020, 8, 1808. https://doi.org/10.3390/microorganisms8111808
Gestal MC, Blas-Machado U, Johnson HM, Rubin LN, Dewan KK, Bryant C, Tiemeyer M, Harvill ET. Disrupting Bordetella Immunosuppression Reveals a Role for Eosinophils in Coordinating the Adaptive Immune Response in the Respiratory Tract. Microorganisms. 2020; 8(11):1808. https://doi.org/10.3390/microorganisms8111808
Chicago/Turabian StyleGestal, Monica C., Uriel Blas-Machado, Hannah M. Johnson, Lily N. Rubin, Kalyan K. Dewan, Claire Bryant, Michael Tiemeyer, and Eric T. Harvill. 2020. "Disrupting Bordetella Immunosuppression Reveals a Role for Eosinophils in Coordinating the Adaptive Immune Response in the Respiratory Tract" Microorganisms 8, no. 11: 1808. https://doi.org/10.3390/microorganisms8111808