Trichoderma harzianum Inoculation Reduces the Incidence of Clubroot Disease in Chinese Cabbage by Regulating the Rhizosphere Microbial Community


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Site
2.2. Experimental Design
2.3. Bioformulation
2.4. Sample Collection
2.5. Assessment of Clubroot Disease Incidence and Severity
2.6. Bacterial and Fungal Genomic DNA Extraction
2.7. PCR Amplification and High-Throughput Sequencing
2.8. qPCR Amplification
2.9. Processing of Bioinformatics and Data Analysis
2.9.1. Differential Analysis of Bacteria and Fungi
2.9.2. Differential Analysis of Fungi
3. Results
3.1. Field Test Results
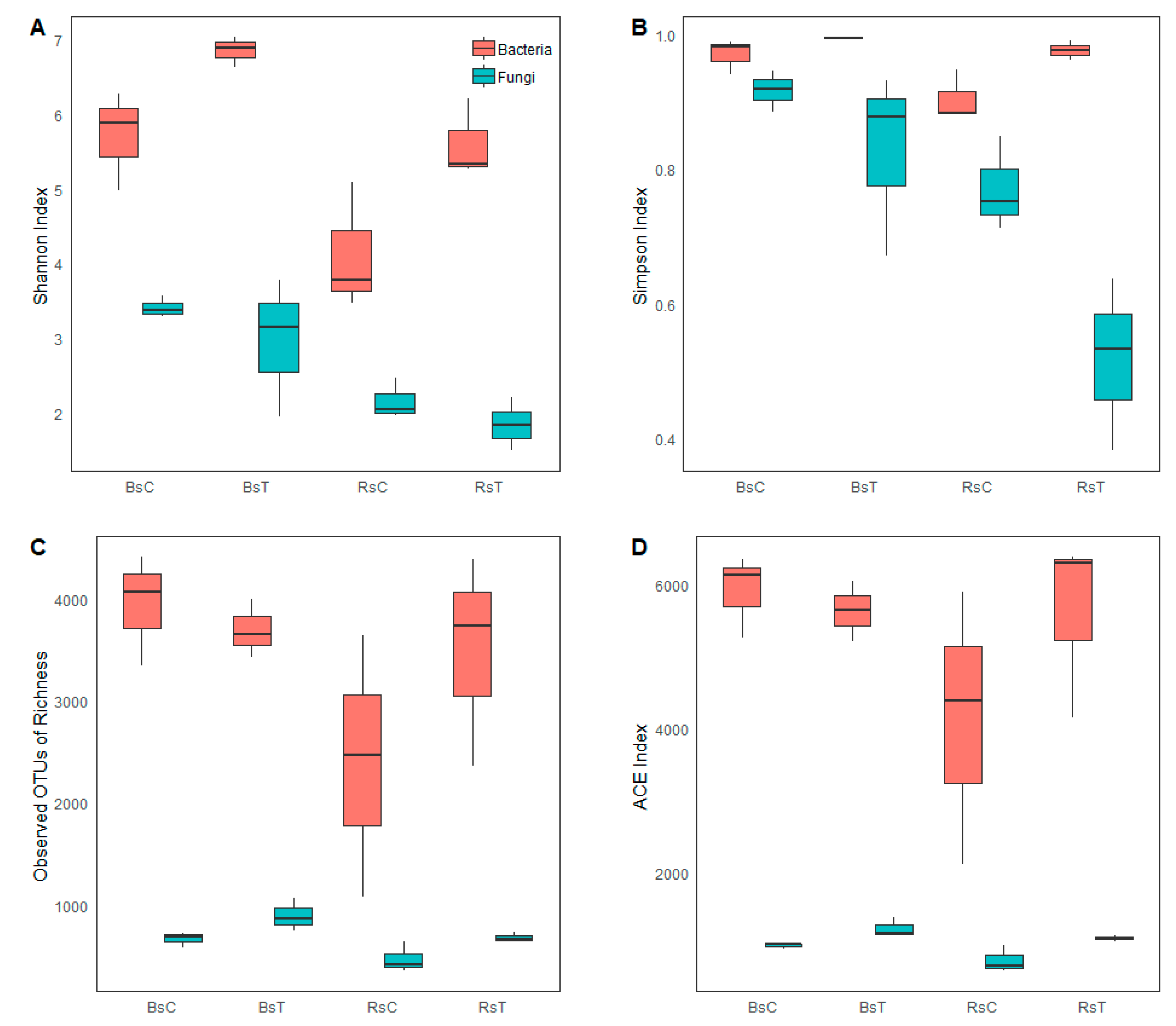
3.2. Alpha Diversity Analysis
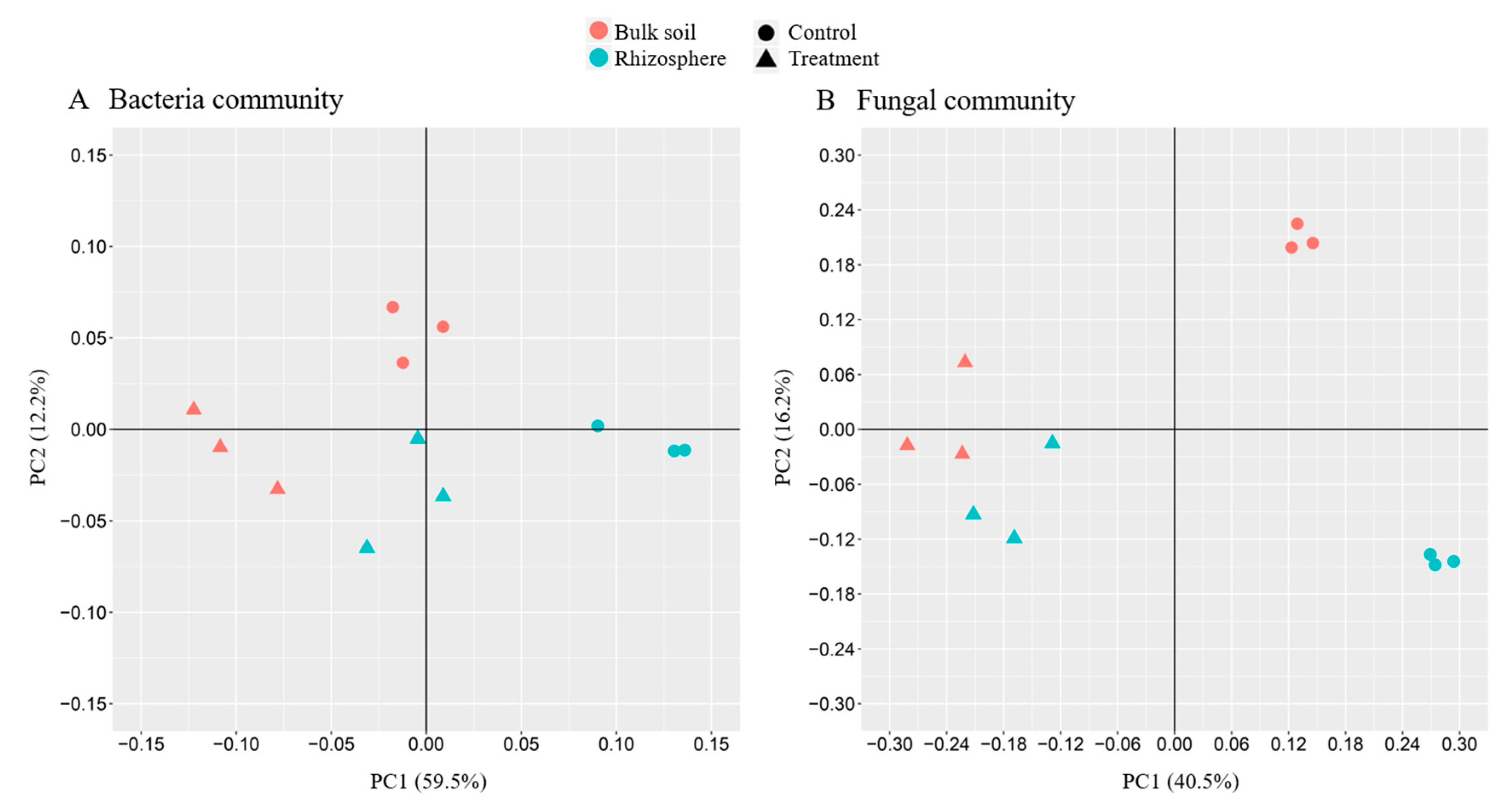
3.3. Beta Diversity Analysis
3.3.1. Principal Component Analysis
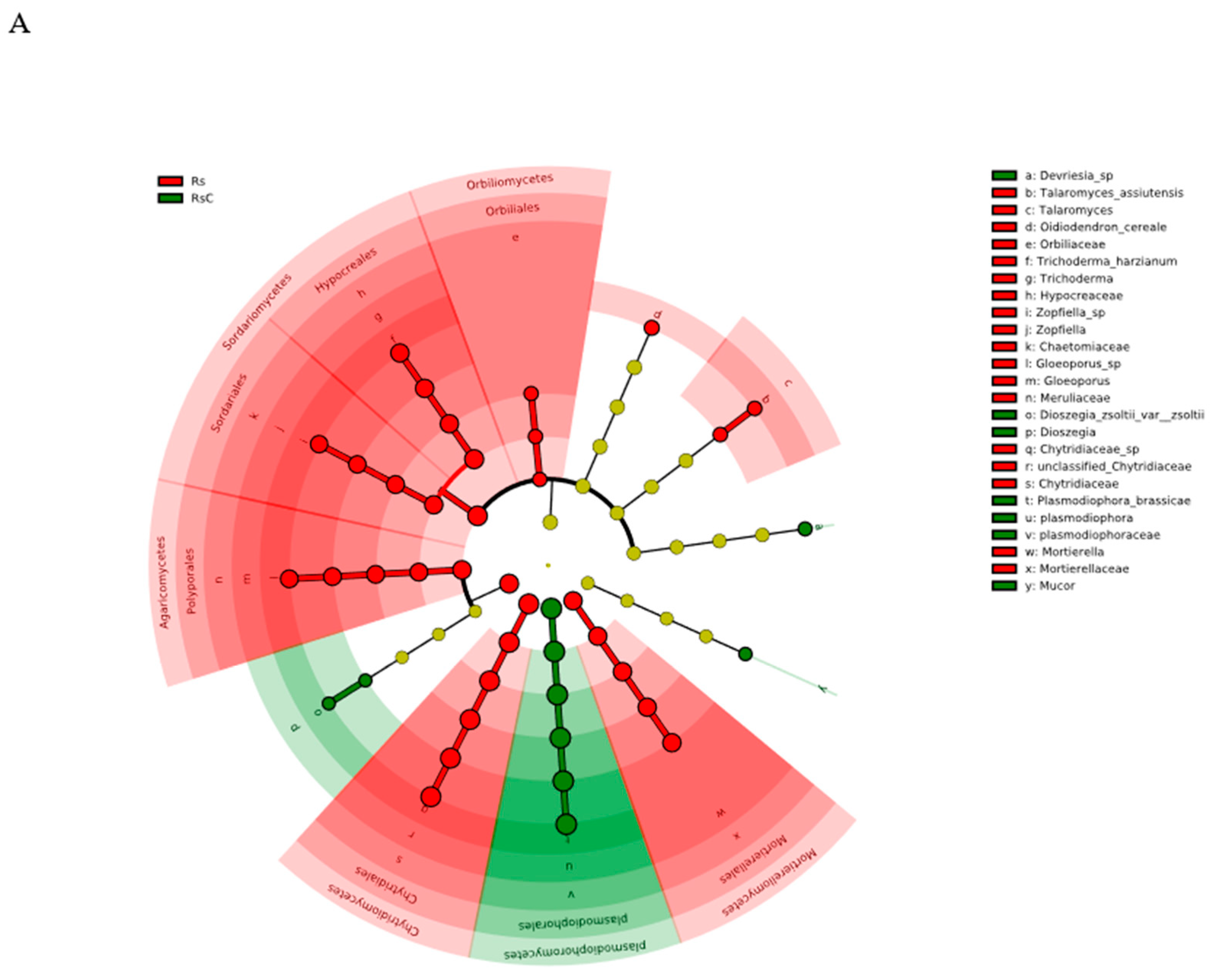
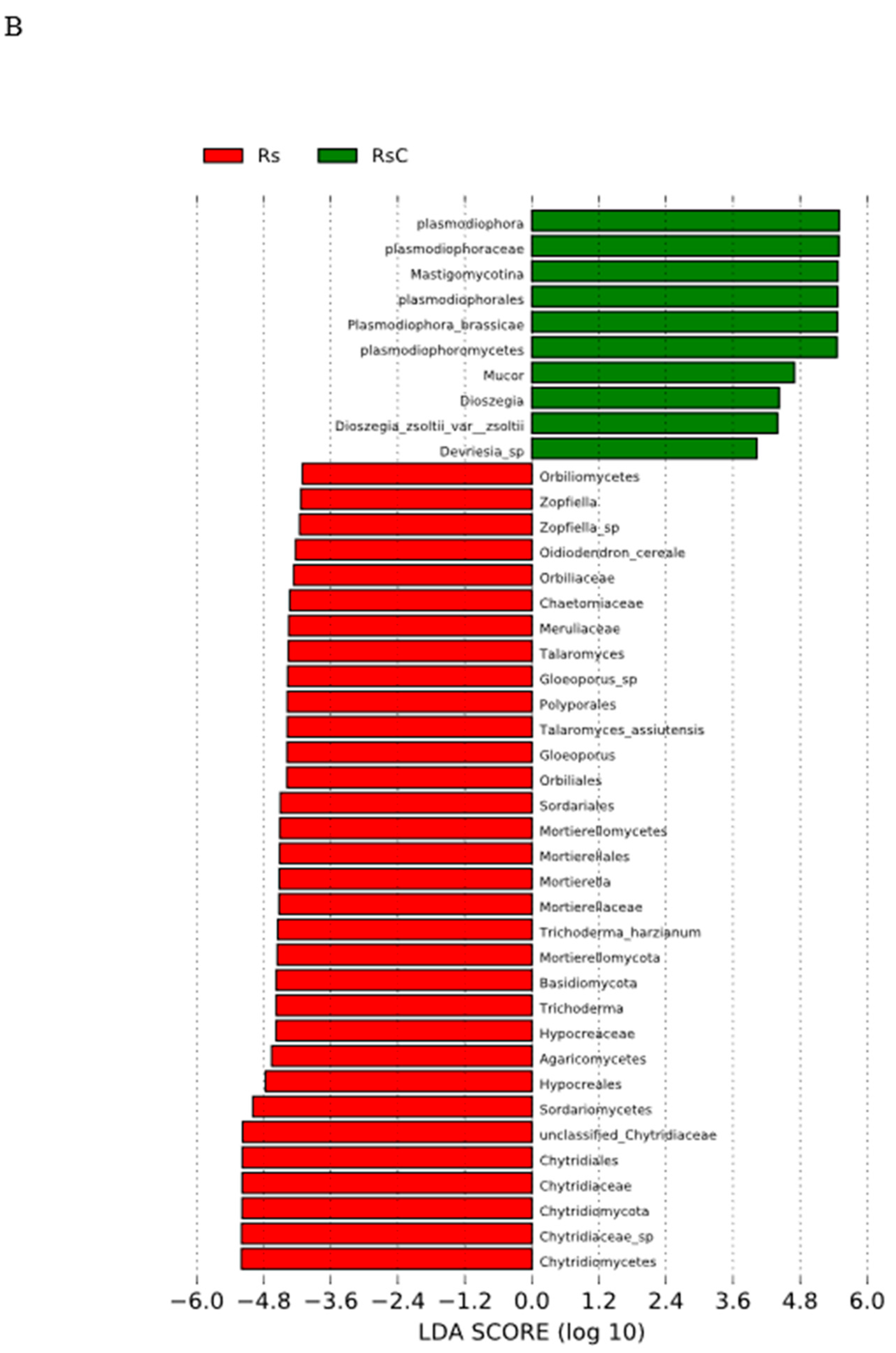
3.3.2. Linear Discriminant Analysis Effect Size (LEfSe)
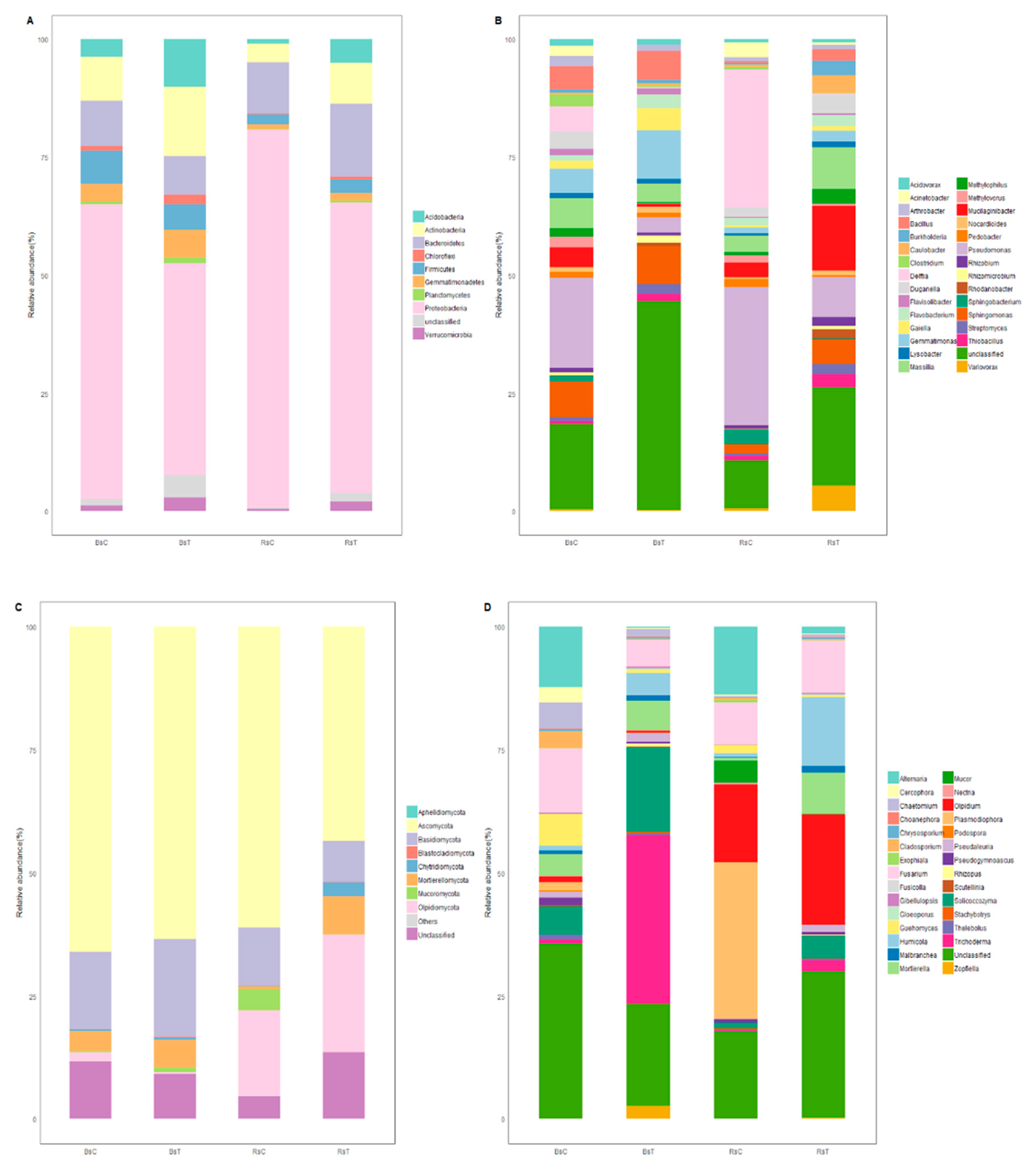
3.4. Community Analysis
3.4.1. Bacteria
3.4.2. Fungi
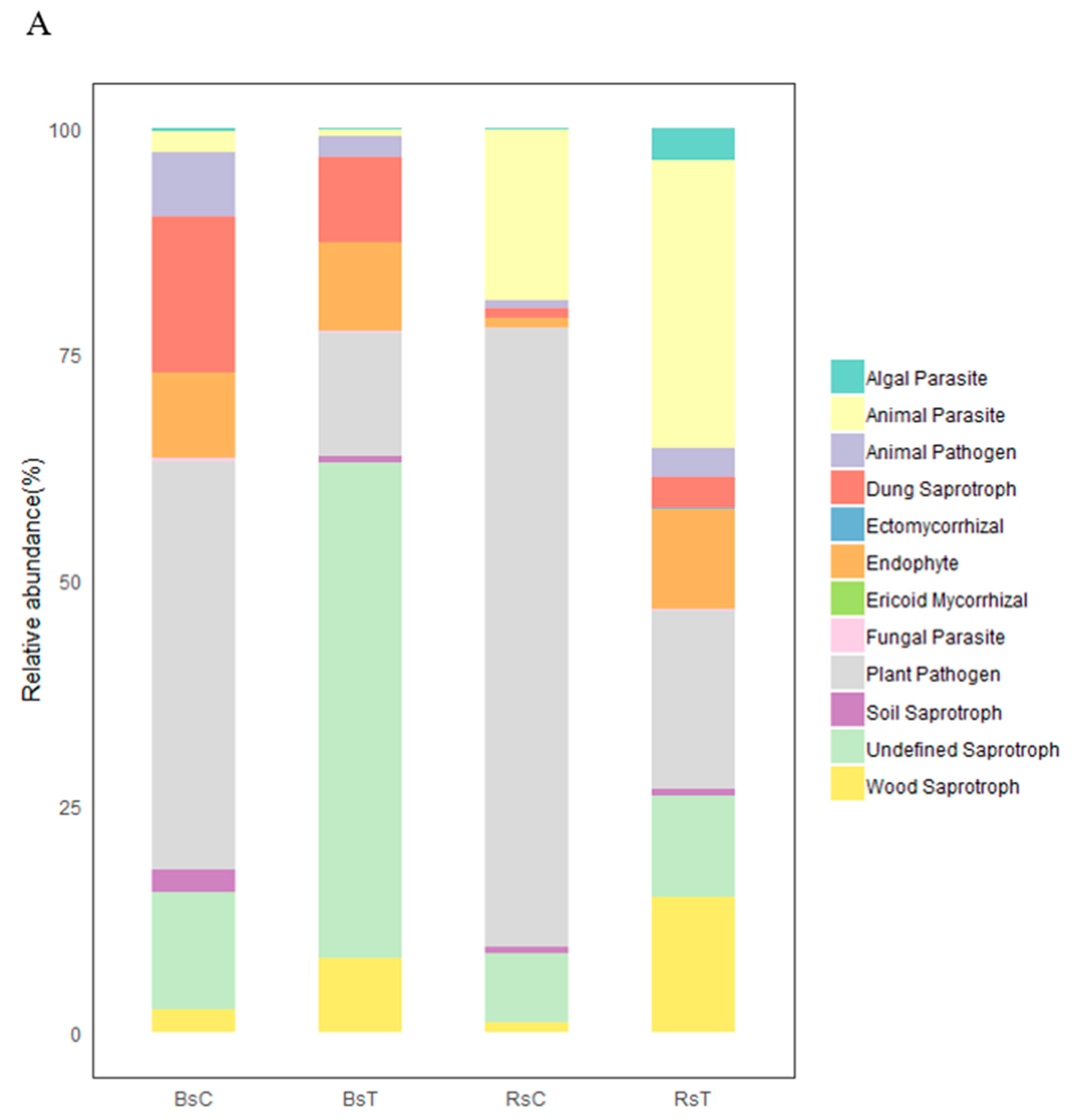
3.4.3. Flora Function, Prediction and Analysis
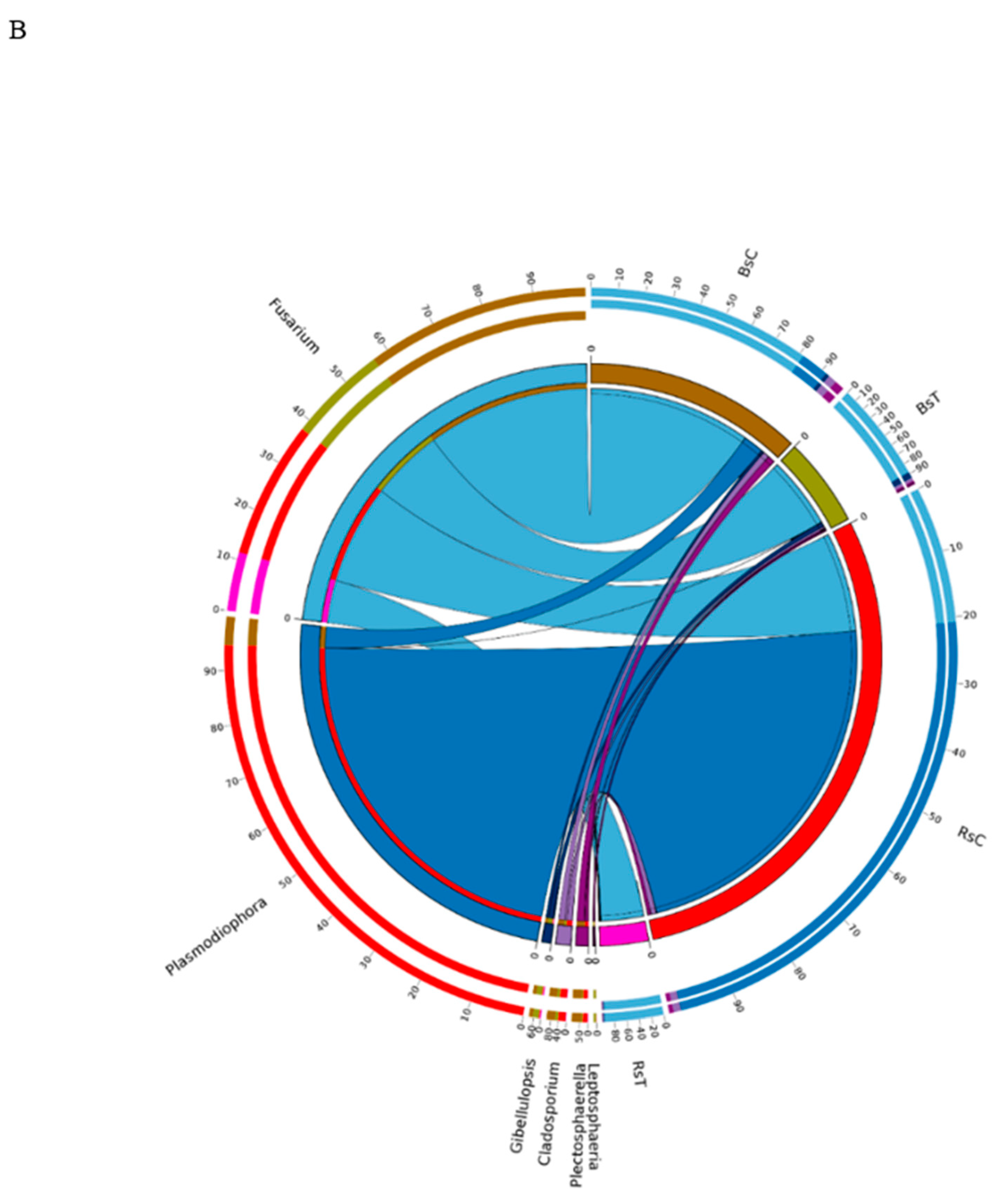
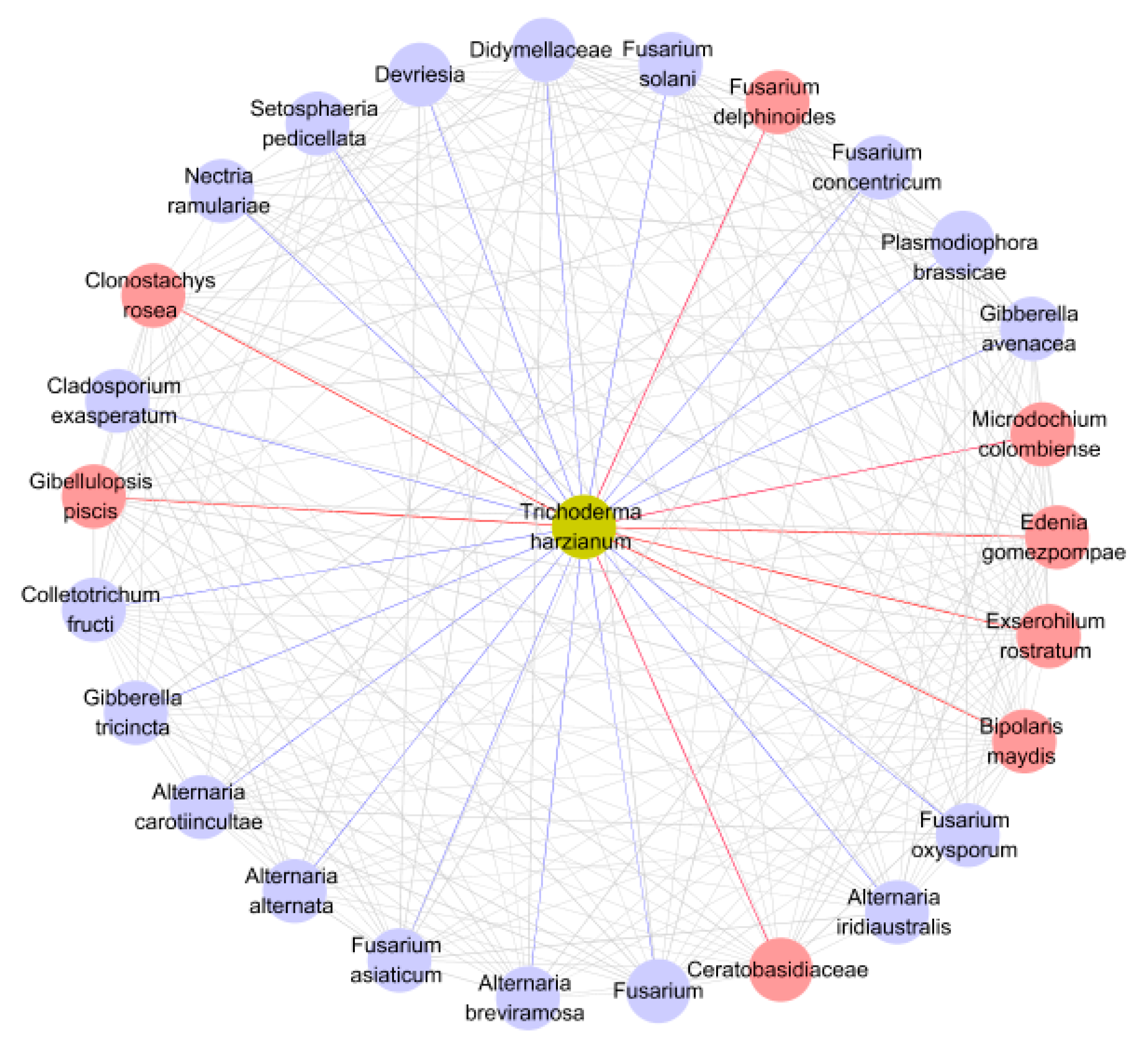
3.5. Co-Occurrence Network Analysis
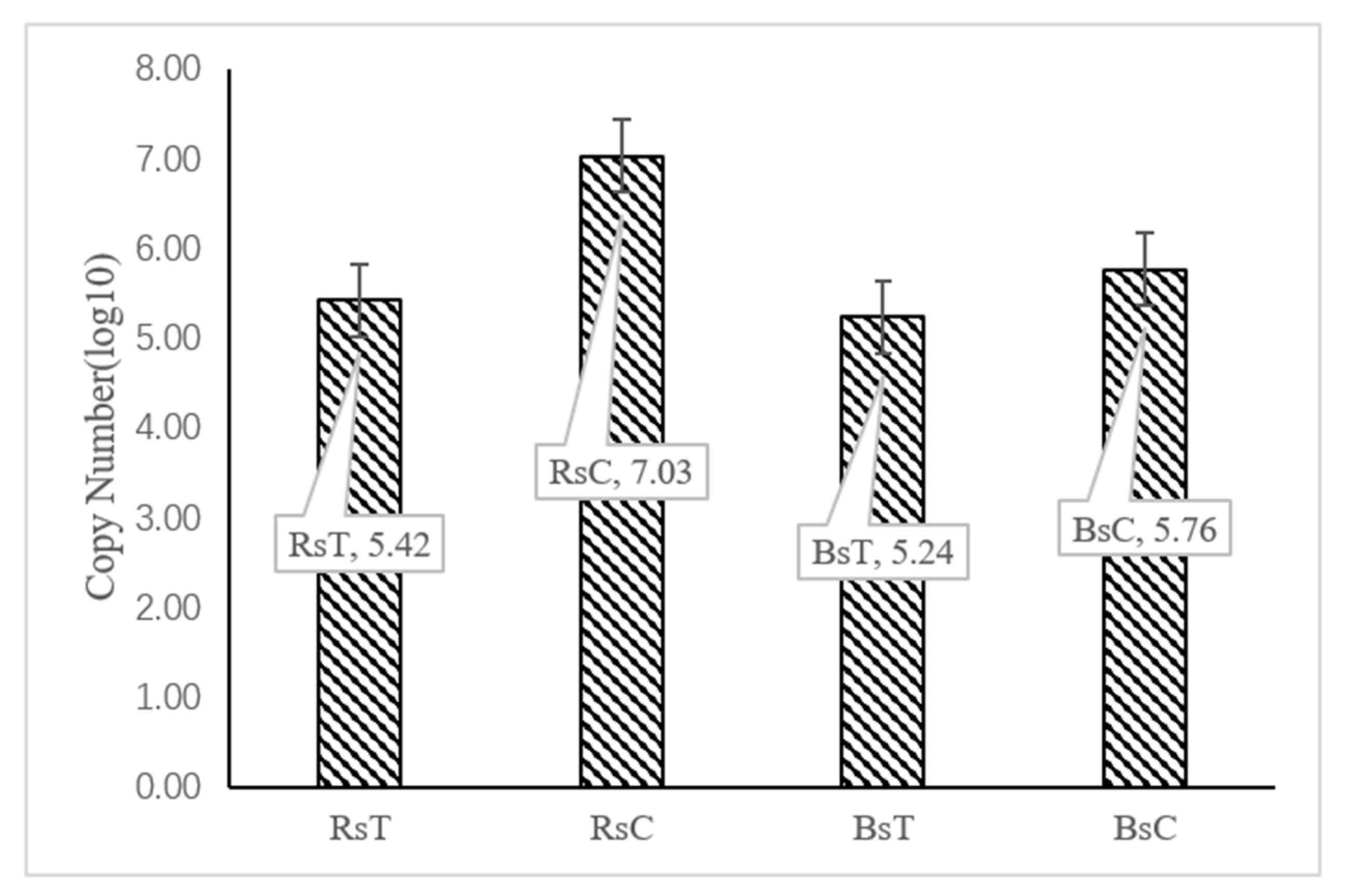
3.6. Analysis by qPCR
4. Discussion
4.1. Fungal Community
4.2. Bacterial Community
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Voorrips, R.E. Plasmodiophora brassicae: Aspects of pathogenesis and resistance in Brassica oleracea. Euphytica 1995, 83, 139–146. [Google Scholar] [CrossRef]
- Dixon, G.R. The occurrence and economic impact of Plasmodiophora brassicae and clubroot disease. J. Plant Growth Regul. 2009, 28, 194–202. [Google Scholar] [CrossRef]
- Howard, R.J.; Strelkov, S.E.; Harding, M.W. Clubroot of cruciferous crops–new perspectives on an old disease. Can. J. Plant Pathol. 2010, 32, 43–57. [Google Scholar] [CrossRef]
- Kageyama, K.; Asano, T. Life cycle of Plasmodiophora brassicae. J. Plant Growth Regul. 2009, 28, 203–211. [Google Scholar] [CrossRef]
- Ludwig- Müller, J.; Prinsen, E.; Rolfe, S.A.; Scholes, J.D. Metabolism and plant hormone action during clubroot disease. J. Plant Growth Regul. 2009, 28, 229–234. [Google Scholar] [CrossRef]
- Ludwig- Müller, J.; Schuller, A. What can we learn from clubroots: Alterations in host roots and hormone homeostasis caused by Plasmodiophora brassicae. Eur. J. Plant Pathol. 2008, 121, 291–302. [Google Scholar] [CrossRef]
- Malinowski, R.; Smith, J.A.; Fleming, A.J.; Scholes, J.D.; Rolfe, S.A. Gall formation in clubroot-infected Arabidopsis results from an increase in existing meristematic activities of the host but is not essential for the completion of the pathogen life cycle. Plant J. 2012, 71, 226–238. [Google Scholar] [CrossRef]
- Cranmer, T.J.; Al-Daoud, F.; Gossen, B.D.; Deora, A.; Hwang, S.F.; Mcdonald, M.R. Vertical distribution of resting spores of Plasmodiophora brassicae in soil. Eur. J. Plant Pathol. 2017, 149, 435–442. [Google Scholar] [CrossRef]
- Botero, A.; Garcia, C.; Gossen, B.D.; Strelkov, S.E.; Todd, C.D.; Bonhamsmith, P.C.; Perezlopez, E. Clubroot disease in Latin America: Distribution and management strategies. Plant Pathol. 2019, 68, 827–833. [Google Scholar] [CrossRef] [Green Version]
- Wallenhammar, A.C. Prevalence of Plasmodiophora brassicae in a spring oilseed rape growing area in central Sweden and factors influencing soil infestation levels. Plant Pathol. 1996, 45, 710–719. [Google Scholar] [CrossRef]
- Dixon, G.R. Managing clubroot disease (caused by Plasmodiophora brassicae Wor.) by exploiting the interactions between calcium cyanamide fertilizer and soil microorganisms. J. Agric. Sci. 2016, 155, 527–543. [Google Scholar] [CrossRef]
- Murakami, H.; Tsushima, S.; Kuroyanagi, Y.; Shishido, Y. Reduction of resting spore density of Plasmodiophora brassicae and clubroot disease severity by liming. Soil Sci. Plant Nutr. 2002, 48, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.; Kochi, S.; Kunita, H.; Ito, S.; Kameyaiwaki, M. Biological mode of action of the fungicide, flusulfamide, against Plasmodiophora brassicae (clubroot). Eur. J. Plant Pathol. 1999, 105, 577–584. [Google Scholar] [CrossRef]
- Kowata-Dresch, L.; May-De Mio, L. Clubroot management of highly infested soils. Crop. Prot. 2012, 35, 47–52. [Google Scholar] [CrossRef]
- Liu, C.; Yang, Z.; He, P.; Munir, S.; He, P.; Wu, Y.; Ho, H.; He, Y. Fluazinam positively affected the microbial communities in clubroot cabbage rhizosphere. Sci. Hortic. 2019, 256, 108519. [Google Scholar] [CrossRef]
- Sakkas, V.A.; Lambropoulou, D.A.; Albanis, T.A. Study of chlorothalonil photodegradation in natural waters and in the presence of humic substances. Chemosphere 2002, 48, 939–945. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.; Dong, L.; Li, X.; Asif, M.; Guo, Q.; Jiang, W.; Ma, P.; Zhang, L. Fengycin produced by Bacillus subtilis NCD-2 is involved in suppression of clubroot on Chinese cabbage. Biol. Control 2019, 136, 104001. [Google Scholar] [CrossRef]
- Lorito, M.; Woo, S. Trichoderma: A multi-purpose tool for integrated pest management. In Principles of Plant-Microbiome Interactions; Lutenberg, B., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 345–353. [Google Scholar]
- Harman, G.E. Overview of mechanisms and uses of Trichoderma spp. Phytopathology 2006, 96, 190–194. [Google Scholar] [CrossRef] [Green Version]
- Mahdizadehnaraghi, R.; Heydari, A.; Zamanizadeh, H.R.; Rezaee, S.; Nikan, J. Biological control of garlic (Allium) white rot disease using antagonistic fungi-based bioformulations. J. Plant Prot. Res. 2015, 55, 136–141. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, Y.; Chen, K.; Li, J.; Wei, Y.; Wang, Y.; Wu, Y.; Ryder, M.H.; Hetong, Y.; Denton, M.D. Large-scale Trichoderma diversity was associated with ecosystem, climate and geographic location. Environ. Microbiol. 2020. Accepted. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, Z.; Yang, X.; Ran, W.; Shen, Q. Trichoderma harzianum T-E5 significantly affects cucumber root exudates and fungal community in the cucumber rhizosphere. Appl. Soil Ecol. 2013, 72, 41–48. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Zhuang, L.; Yu, Y.; Liu, J.; Zhang, L.; Gao, Z.; Wu, Y.; Gao, W.; Ding, G.; et al. A Rhizosphere-Derived Consortium of Bacillus subtilis and Trichoderma harzianum Suppresses Common Scab of Potato and Increases Yield. Comput. Struct. Biotechnol. J. 2019, 17, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Yuan, J.; Yang, X.; Cui, Y.; Chen, L.; Ran, W.; Shen, Q. Putative Trichoderma harzianum mutant promotes cucumber growth by enhanced production of indole acetic acid and plant colonization. Plant Soil. 2013, 368, 433–444. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Horwitz, B.A.; Herrera-Estrella, A.; Schmoll, M.; Kenerley, C.M. Trichoderma research in the genome era. Annu. Rev. Phytopathol. 2013, 51, 105–129. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Shukla, N.; Kabadwal, B.C.; Tewari, A.K.; Kumar, J. Review on plant-Trichoderma-pathogen interaction. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2382–2397. [Google Scholar] [CrossRef]
- Hajiehgrari, B.; Torabi-Giglou, M.; Mohammadi, M.R.; Davari, M.M. Biological potential of some Iranian Trichoderma isolates in the control of soil borne plant pathogenic fungi. Afr. J. Biol. Control. 2008, 7, 967–972. [Google Scholar]
- Nihorimbere, V.; Cawoy, H.; Seyer, A.; Brunelle, A.; Thonart, P.; Ongena, M. Impact of rhizosphere factors on cyclic lipopeptide signature from the plant beneficial strain Bacillus amyloliquefaciens S499. FEMS Microbiol. Ecol. 2012, 79, 176–191. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Chen, K.; Wu, Y.; Hu, J.; Wei, Y.; Li, J.; Yang, H.; Ryder, M.; Denton, M.D. Near-complete genomes of two Trichoderma species: A resource for biological control of plant pathogens. Mol. Plant Microbe Interact. 2020. Accepted. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Zhao, Y.; Cheng, J.; Wang, W. Biocontrol effect of Trichoderma harzianum T4 on brassica clubroot and analysis of rhizosphere microbial communities based on T-RFLP. Biocontrol Sci. Technol. 2015, 25, 1493–1505. [Google Scholar] [CrossRef]
- Gao, G.; Yin, D.; Chen, S.; Xia, F.; Yang, J.; Li, Q.; Wang, W. Effect of biocontrol agent Pseudomonas fluorescens 2P24 on soil fungal community in cucumber rhizosphere using T-RFLP and DGGE. PLoS ONE 2012, 7, e31806. [Google Scholar] [CrossRef]
- Zhong, Y.; Yang, Y.; Liu, P.; Xu, R.; Rensing, C.; Fu, X.; Liao, H. Genotype and rhizobium inoculation modulate the assembly of soybean rhizobacterial communities. Plant Cell Environ. 2019, 42, 2028–2044. [Google Scholar] [CrossRef] [PubMed]
- Blaalid, R.; Carlsen, T.; Kumar, S.; Halvorsen, R.; Ugland, K.I.; Fontana, G.; Kauserud, H. Changes in the root-associated fungal communities along a primary succession gradient analysed by 454 pyrosequencing. Mol. Ecol. 2012, 21, 1897–1908. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, H.; Ge, X.; Wu, F. Effects of vanillin on the community structures and abundances of Fusarium and Trichoderma spp. in cucumber seedling rhizosphere. J. Plant Interact. 2018, 13, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Saravanakumar, K.; Li, Y.; Yu, C.; Wang, Q.; Wang, M.; Sun, J.; Gao, J.; Chen, J. Effect of Trichoderma harzianum on maize rhizosphere microbiome and biocontrol of Fusarium Stalk rot. Sci. Rep. 2017, 7, 1771. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Ioannidis, P.; Lengelle, J.; Cebron, A.; Morin, E.; Buee, M.; Martin, F. Functional assays and metagenomic analyses reveals differences between the microbial communities inhabiting the soil horizons of a Norway spruce plantation. PLoS ONE 2013, 8, e55929. [Google Scholar] [CrossRef]
- Rennie, D.C.; Manolii, V.P.; Cao, T.; Hwang, S.F.; Howard, R.J.; Strelkov, S.E. Direct evidence of surface infestation of seeds and tubers by Plasmodiophora brassicae and quantification of spore loads. Plant Pathol. 2011, 60, 811–819. [Google Scholar] [CrossRef]
- Wallenhammar, A.C.; Almquist, C.; Söderström, M.; Jonsson, A. In-field distribution of Plasmodiophora brassicae measured using quantitative real-time PCR. Plant Pathol. 2012, 61, 16–28. [Google Scholar] [CrossRef]
- Faggian, R.; Hanson, M.; Kennedy, R.; Petch, G.; Wakeham, A. Assessment of the response of Plasmodiophora brassicae in contaminated horticultural land, using lime-based fertilizer concentrations. Food Energy Secur. 2017, 6, e00122. [Google Scholar] [CrossRef]
- Al-Daoud, F.; Gossen, B.D.; Robson, J.; McDonald, M.R. Propidium Monoazide Improves Quantification of Resting Spores of Plasmodiophora brassicae with qPCR. Plant Dis. 2017, 3, 442–447. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, L.; Liu, Y.; Yang, J.; Ji, G. Detection of Plasmodiophora brassicae with Real-time Quantitative PCR in Yunnan Province. J. Yunnan Agric. Univ. 2016, 31, 43–48. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, PlatformIndependent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzywinski, M.; Schein, J.E.; Birol, I.; Connors, J.M.; Gascoyne, R.D.; Horsman, D.; Jones, S.J.M.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2015, 20, 241–248. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcıa, R.; Riera, R.; Zambrano, C.; Gutierrez, L. Desarrollo de un fungicida biologico a base de una cepa del hongo Trichodermaharzianum proveniente de la region andina venezolana. Fitosanidad 2006, 10, 115–121. [Google Scholar]
- Botero-Ramırez, A. Effect of Three Trichoderma Species on Clubroot Disease in Cabbage. Master’s Thesis, Universidad Nacional de Colombia, Bogot, Colombia, 2016. [Google Scholar]
- Santos, C.A.; Sobrinho, N.M.B.A.; Costa, E.S.P.; Diniz, C.S.; do Carmo, M.G.F. Liming and biofungicide for the control of clubroot in cauliflower. Pesqui. Agropec. Trop. 2017, 47, 303–311. [Google Scholar] [CrossRef]
- Debode, J.; de Tender, C.; Cremelie, P.; Lee, A.S.; Kyndt, T.; Muylle, H.; de Swaef, T.; Vandecasteele, B. Trichoderma-inoculated Miscanthus straw can replace peat in strawberry cultivation, with beneficial effects on disease control. Front. Plant Sci. 2018, 9, 213. [Google Scholar] [CrossRef]
- Seaman, A. Efficacy of OMRI-Approved Products for Tomato Foliar Disease Control; New York State IPM Program: New York, NY, USA, 2003; pp. 164–167. [Google Scholar]
- Fotoohiyan, Z.; Rezaee, S.; Bonjar, G.H.S.; Mohammadi, A.H.; Moradi, M. Biocontrol potential of Trichoderma harzianum in controlling wilt disease of pistachio caused by Verticillium dahliae. J. Plant Prot. Res. 2017, 57, 185–193. [Google Scholar] [CrossRef]
- Peng, G.; Mcgregor, L.; Lahlali, R.; Gossen, B.D.; Hwang, S.F.; Adhikari, K.K.C.; Strelkov, S.E.; Mcdonald, M.R. Potential biological control of clubroot on canola and crucifer vegetable crops. Plant Pathol. 2011, 60, 566–574. [Google Scholar] [CrossRef]
- Wang, J.; Huang, Y.; Lin, S.; Liu, F.; Song, Q.; Peng, Y.; Zhao, L. A strain of Streptomyces griseoruber isolated from rhizospheric soil of Chinese cabbage as antagonist to Plasmodiophora brassicae. Ann. Microbiol. 2012, 62, 247–253. [Google Scholar] [CrossRef]
- Tian, X.; Wang, D.; Mao, Z.; Pan, L.; Liao, J.; Cai, Z. Infection of Plasmodiophora brassicae changes the fungal endophyte community of tumourous stem mustard roots as revealed by high-throughput sequencing and culturedependent methods. PLoS ONE 2019, 14, e0214975. [Google Scholar] [CrossRef] [Green Version]
- Lori, G.; Edel-Hermann, V.; Gautheron, N.; Alabouvette, C. Genetic diversity of pathogenic and nonpathogenic populations of Fusarium oxysporum isolated from carnation fields in Argentina. Phytopathology 2004, 94, 661–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganuza, M.; Pastor, N.; Boccolini, M.; Erazo, J.; Palacios, S.; Oddino, C.; Reynoso, M.M.; Rovera, M.; Torres, A.M. Evaluating the impact of the biocontrol agent Trichoderma harzianum ITEM 3636 on indigenous microbial communities from field soils. J. Appl. Microbiol. 2019, 126, 608–623. [Google Scholar] [CrossRef] [PubMed]
- Munir, S.; Wu, Y.; Yang, Z.; Munir, S.; Wu, Y.; Ho, H.; He, Y. Deciphering the bacterial and fungal communities in clubroot-affected cabbage rhizosphere treated with Bacillus Subtilis XF-1. Agric. Ecosyst. Environ. 2018, 256, 12–22. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence |
|---|---|
| 341F | CCCTACACGACGCTCTTCCGAT CTGCCTACGGGNGGCWGCAG |
| 805R | GACTGGAGTTCCTTGGCACCCGAGAA TTCCAGACTACHVGGGTATCTAATCC |
| ITS3 | CCCTACACGACGCTCTTCCGATC TNTCCTCCGCTTATTGATATGC |
| ITS4 | GTGACTGGAGTTCCTTGGCACCCGAG AATTCCAGCATCGATGAAGAACGCAGC |
| Data | Disease Index | Disease Incidence | PDIDS | |
|---|---|---|---|---|
| Sample | ||||
| Inoculation | 23.2% | 51.3% | 63% | |
| Control | 62.5% | 96.7% | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Philp, J.; Li, J.; Wei, Y.; Li, H.; Yang, K.; Ryder, M.; Toh, R.; Zhou, Y.; Denton, M.D.; et al. Trichoderma harzianum Inoculation Reduces the Incidence of Clubroot Disease in Chinese Cabbage by Regulating the Rhizosphere Microbial Community. Microorganisms 2020, 8, 1325. https://doi.org/10.3390/microorganisms8091325
Li J, Philp J, Li J, Wei Y, Li H, Yang K, Ryder M, Toh R, Zhou Y, Denton MD, et al. Trichoderma harzianum Inoculation Reduces the Incidence of Clubroot Disease in Chinese Cabbage by Regulating the Rhizosphere Microbial Community. Microorganisms. 2020; 8(9):1325. https://doi.org/10.3390/microorganisms8091325
Chicago/Turabian StyleLi, Junhui, Joshua Philp, Jishun Li, Yanli Wei, Hongmei Li, Kai Yang, Maarten Ryder, Ruey Toh, Yi Zhou, Matthew D. Denton, and et al. 2020. "Trichoderma harzianum Inoculation Reduces the Incidence of Clubroot Disease in Chinese Cabbage by Regulating the Rhizosphere Microbial Community" Microorganisms 8, no. 9: 1325. https://doi.org/10.3390/microorganisms8091325