Selection of Exopolysaccharide-Producing Lactobacillus Plantarum (Lactiplantibacillus Plantarum) Isolated from Algerian Fermented Foods for the Manufacture of Skim-Milk Fermented Products

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Strains
2.2. Genome Analysis
2.3. Milk Fermentations
2.4. Exopolysaccharide (EPS) Purification and Analysis
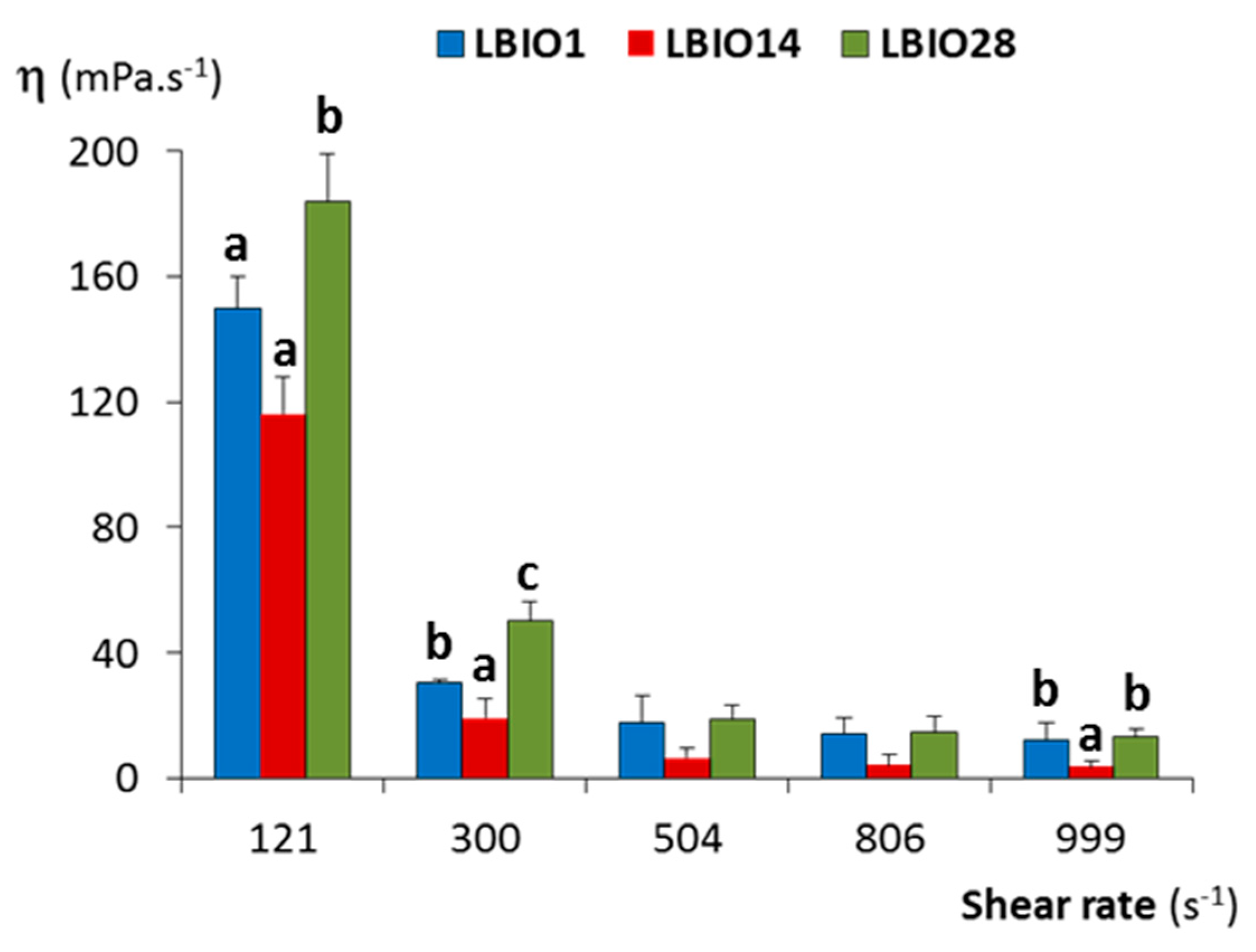
2.5. Viscosity of Fermented Milks
2.6. Macrostructure of Fermented Milks
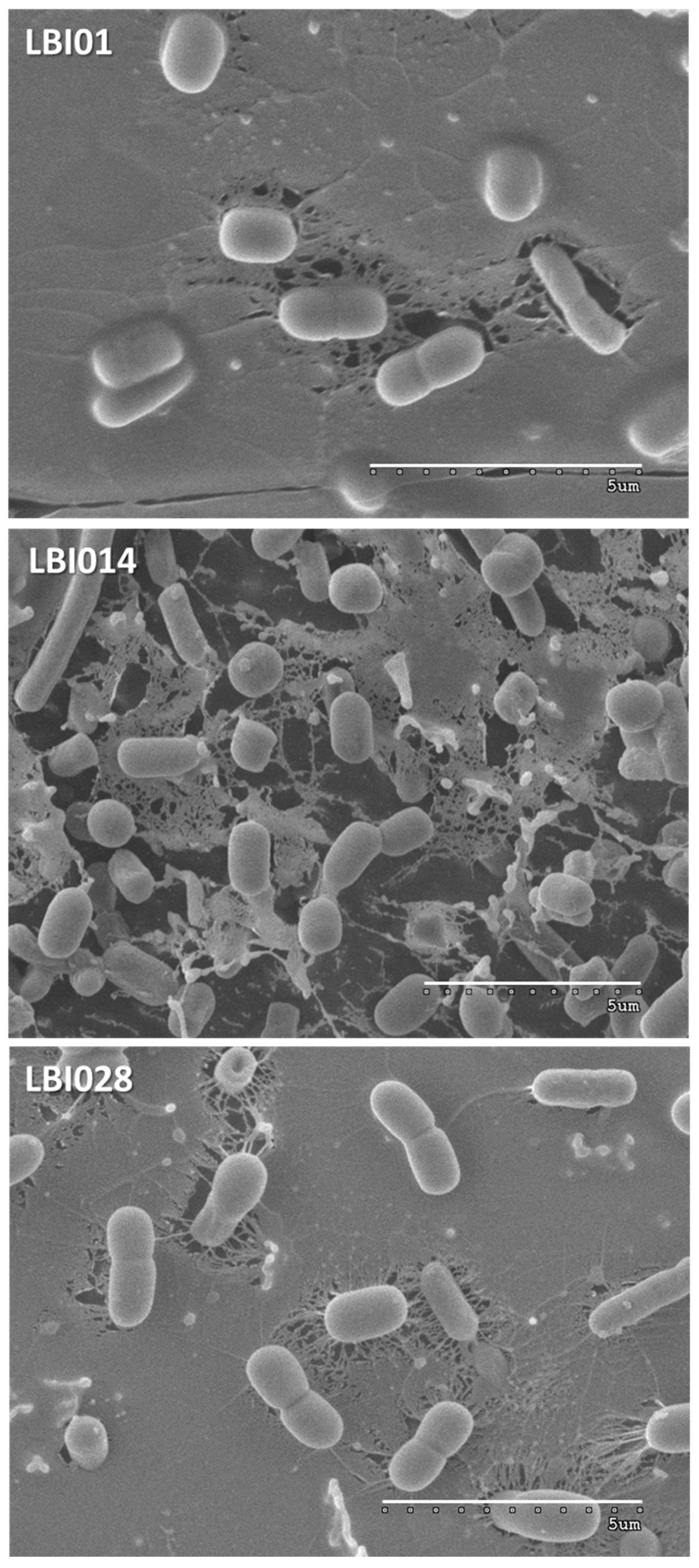
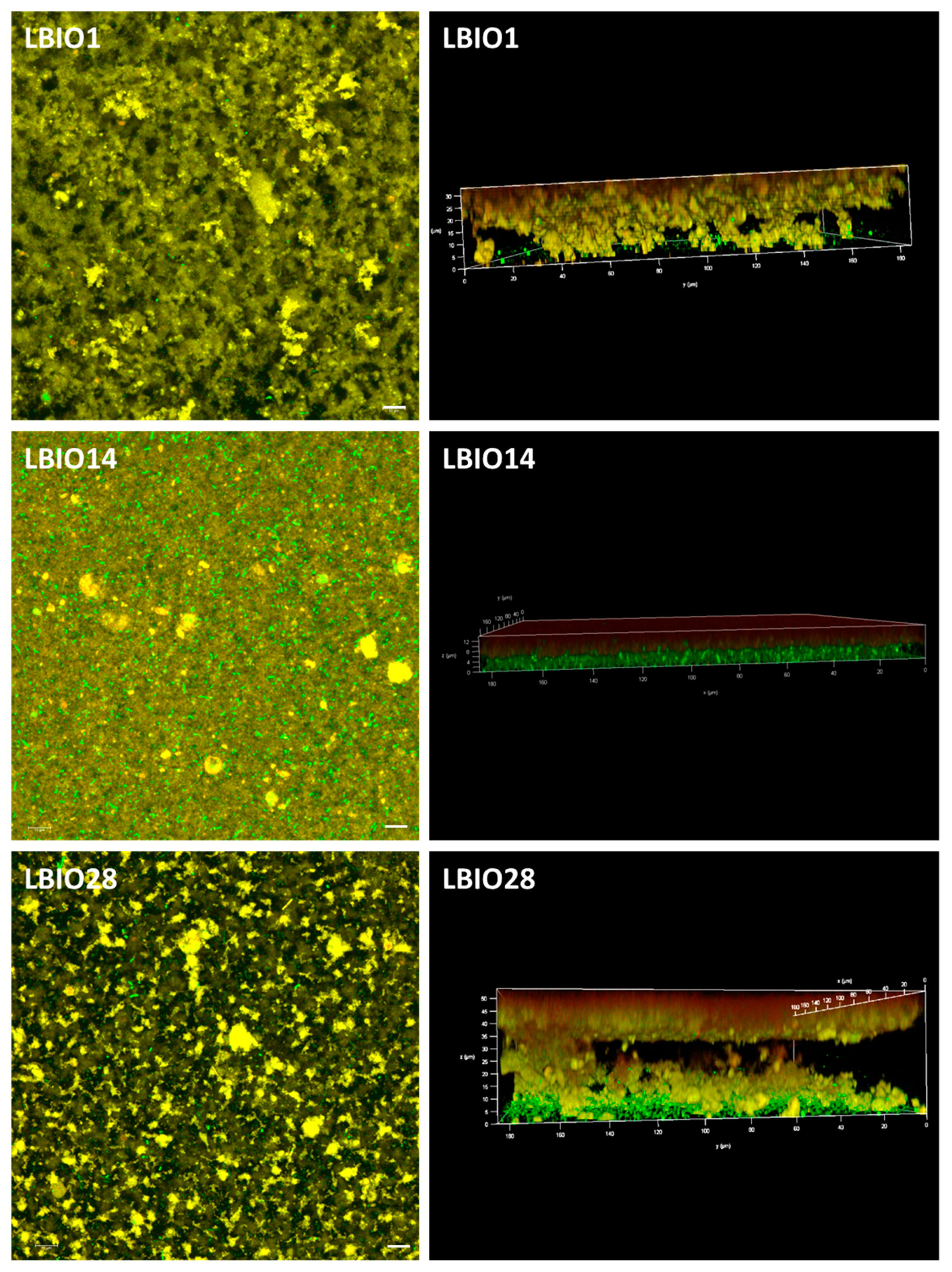
2.7. Microstructure of Fermented Milks
2.8. Statistical Analysis
3. Results and Discussion
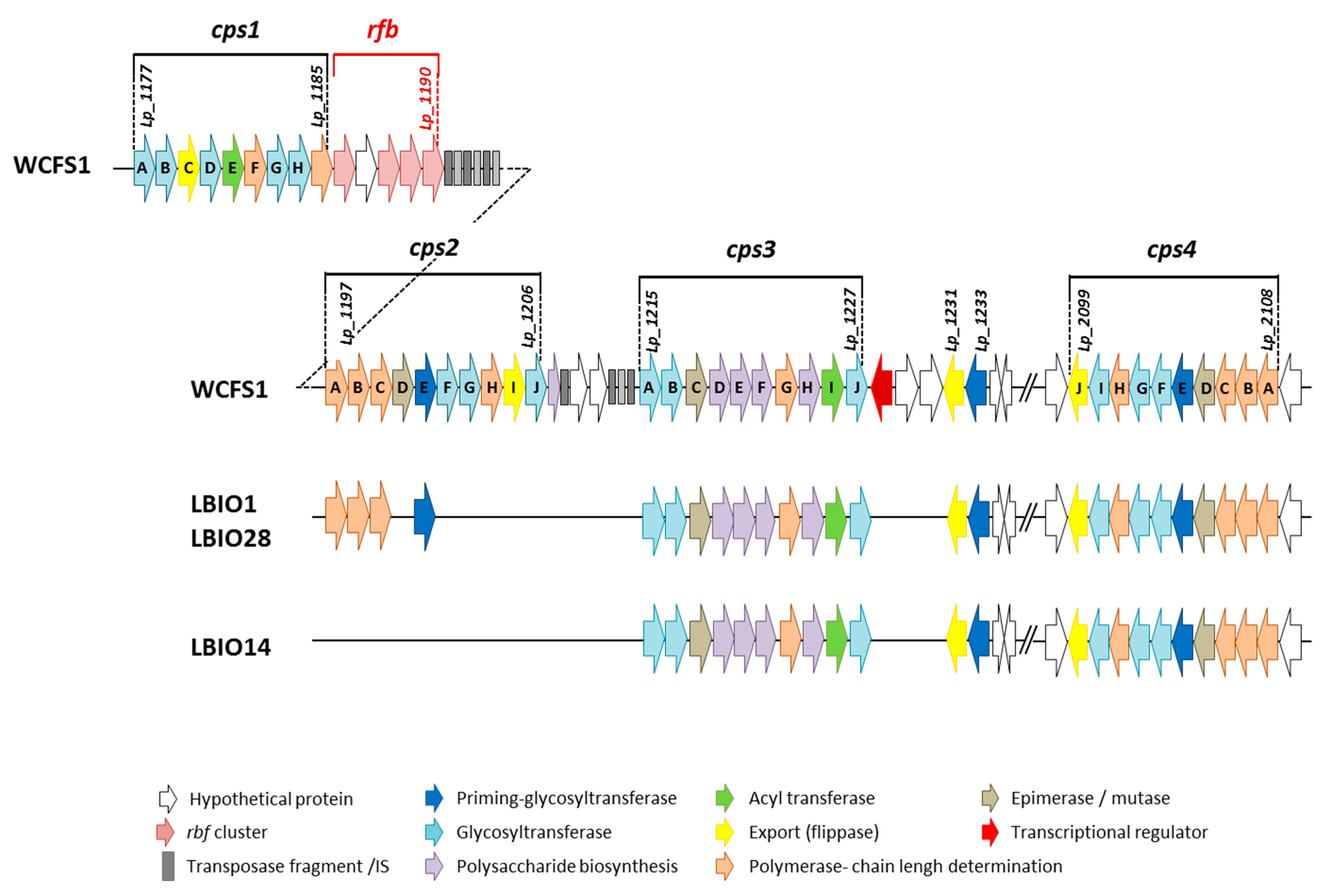
3.1. Insight into the Genomes of L. Plantarum Strains: Focus on eps Cluster Analysis
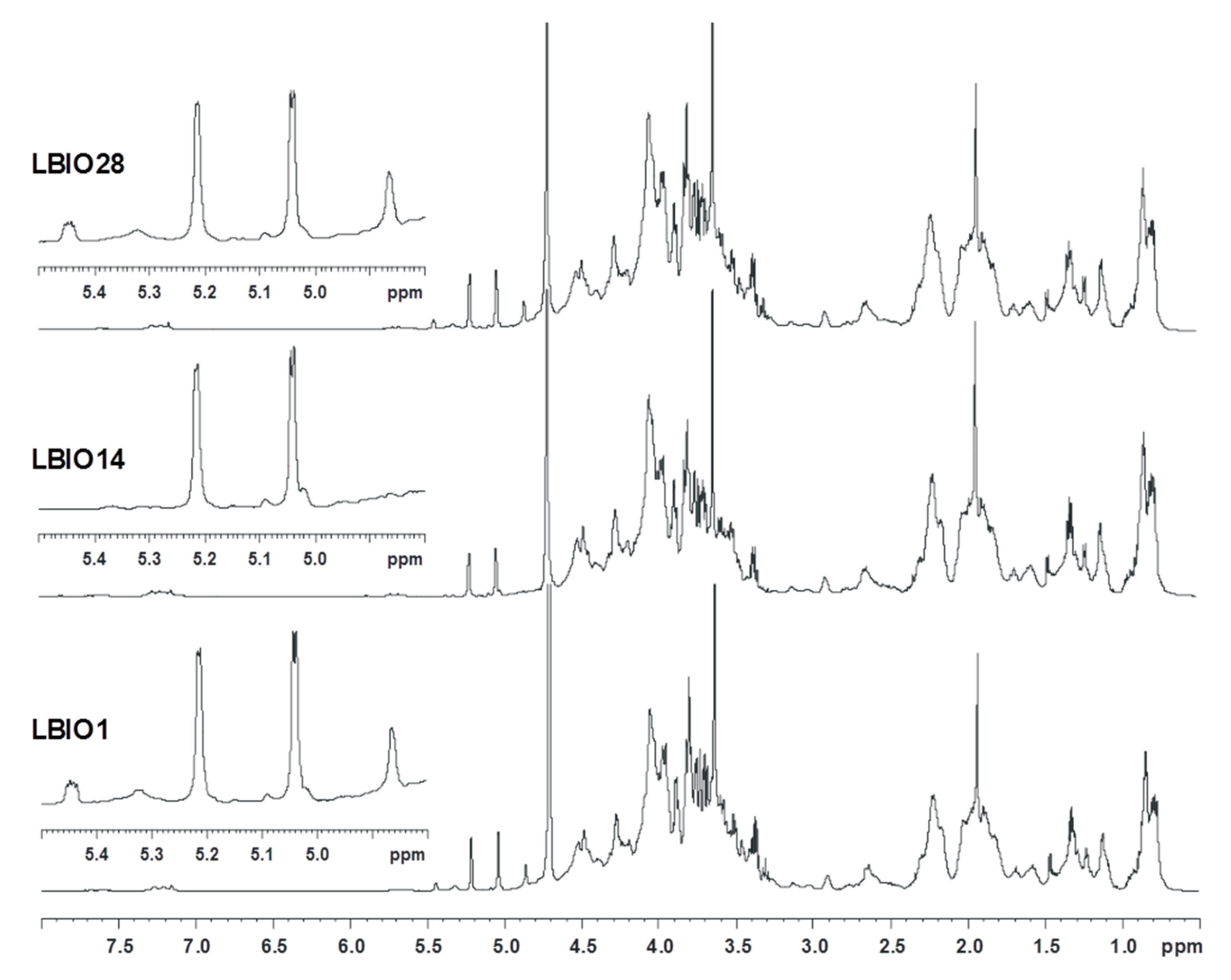
3.2. Characterization of EPS Purified from Fermented Milks
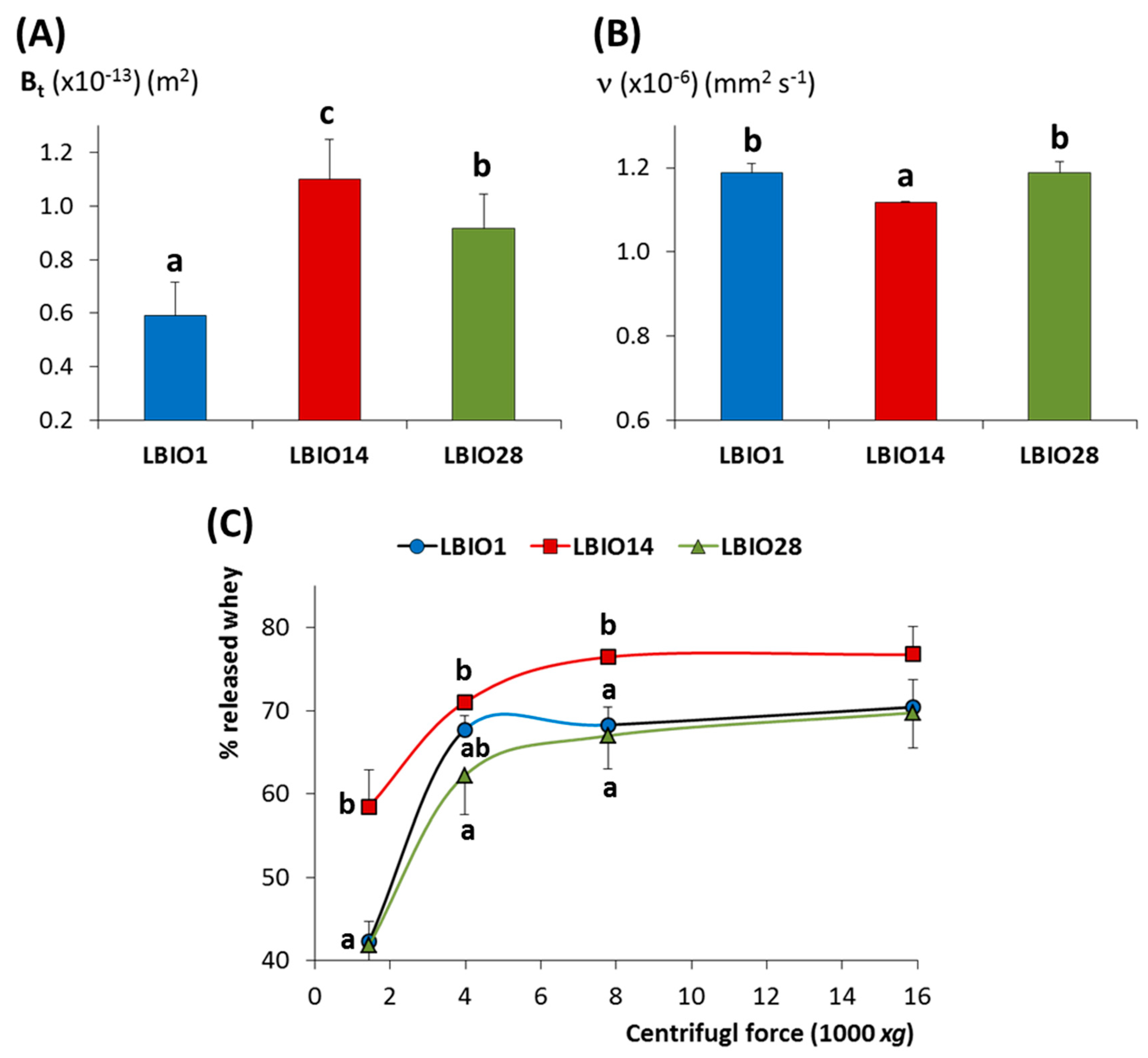
3.3. Characterization of Skim Milks Fermented with the EPS-producing L. Plantarum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carminati, D.; Meucci, A.; Tidona, F.; Zago, M.; Giraffa, G. Multifucntional lactic acid bacteria cultures to improve quality and nutritional benefits in dairy products. In Advances in Food Biotechnology; Ravishankar, R.V., Ed.; Wiley-Blackwell: Oxford, UK, 2016; Chapter 15; pp. 263–276. [Google Scholar]
- Gibbons, J.G.; Rinker, D.C. The genomics of microbial domestication in the fermented food environment. Curr. Opin. Gen. Dev. 2015, 35, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bron, P.A.; Marcelli, B.; Mulder, J.; van der Els, S.; Morawska, L.P.; Kuipers, O.P.; Kok, J.; Kleerebezem, M. Renaissance of traditional DNA transfer strategies for improvement of industrial lactic acid bacteria. Curr. Opin. Biotech. 2019, 56, 61–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leksir, C.; Boudalia, S.; Moujahed, N.; Chemmamm, M. Traditional dairy products in Algeria: Case of Klila cheese. J. Ethn. Foods 2019, 6, 7. [Google Scholar] [CrossRef] [Green Version]
- Zeidan, A.A.; Poulsen, V.K.; Janzen, T.; Buldo, P.; Derkx, P.M.F.; Øregaard, G.; Neves, A.R. Polysaccharide production by lactic acid bacteria: From genes to industrial applications. FEMS Microbiol. Rev. 2017, 41, S168–S200. [Google Scholar] [CrossRef] [Green Version]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic acid bacteria exopolysaccharides in foods and beverages: Isolation, properties, characterization, and health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef]
- Xu, X.; Cui, Y.; Yue, F.; Liu, L.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Exopolysaccharides produced by lactic acid bacteria and bifidobacteria: Structures, physiochemical functions and applications in the food industry. Food Hydrocol. 2019, 94, 475–499. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Sánchez, B.; Milani, C.; Ventura, M.; Margolles, A.; Ruas-Madiedo, P. Genomic overview and biological functions of exopolysaccharide biosynthesis in Bifidobacterium spp. Appl. Environ. Microbiol. 2014, 80, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Castro-Bravo, N.; Wells, J.M.; Margolles, A.; Ruas-Madiedo, P. Interactions of surface exopolysaccharides from Bifidobacterium and Lactobacillus within the intestinal environment. Front. Microbiol. 2018, 9, 2426. [Google Scholar] [CrossRef] [Green Version]
- van Hijum, S.A.F.T.; Kralj, S.; Ozimek, L.K.; Dijkhuizen, L.; van Geel-Schutten, I.G.H. Structure-function relationships of glucansucrase and fructansucrase enzymes from lactic acid bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 157–176. [Google Scholar] [CrossRef] [Green Version]
- Werning, M.L.; Ibarburu, I.; Dueñas, M.T.; Irastorza, A.; Navas, J.; López, P. Pediococcus parvulus gtf gene encoding the GTF glycosyltransferase and its application for specific PCR detection of β-D-glucan–producing bacteria in foods and beverages. J. Food Proct. 2006, 69, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; Hugenholtz, J.; Zoon, P. An overview of the functionality of exopolysaccharides produced by lactic acid bacteria. Int. Dairy J. 2002, 12, 163–171. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Sánchez, B.; Álvarez-Martín, P.; López, P.; Martínez-Álvarez, N.; Delley, M.; Martí, M.; Varela, E.; Suárez, A.; Antolín, M.; et al. A Single mutation in the gene responsible for the mucoid phenotype of Bifidobacterium animalis subsp. lactis confers surface and functional characteristics. Appl. Environ. Microbiol. 2015, 81, 7960–7968. [Google Scholar]
- Hassan, A.N. Possibilities and challenges of exopolysaccharide-producing lactic cultures in dairy foods. J. Dairy Sci. 2008, 91, 1282–1298. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Bachtarzi, N.; Kharroub, K.; Ruas-Madiedo, P. Exopolysaccharide-producing lactic acid bacteria isolated from traditional Algerian dairy products and their application for skim-milk fermentations. LWT Food Sci. Technol. 2019, 107, 117–124. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.; Yang, Z.; Busk, P.K.; Xu, Y.; Yin, Y. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, A.; van Hijum, S.A.F.T.; Bijlsma, J.J.E.; Kok, J.; Kuipers, O.P. BAGEL: A web-based bacteriocin genome mining tool. Nucleic Acids Res. 2006, 34, W273–W279. [Google Scholar] [CrossRef]
- Arndt, D.; Marcu, A.; Liang, Y.; Wishart, D.S. Review PHAST, PHASTER and PHASTEST: Tools for finding prophage in bacterial genomes. Brief. Bioinform. 2019, 20, 1560–1567. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Vielva, L.; de Toro, M.; Lanza, V.F.; de la Cruz, F. PLACNETw: A web-based tool for plasmid reconstruction from bacterial genomes. Bioinformatics 2017, 33, 3796–3798. [Google Scholar] [CrossRef]
- Burns, P.; Vinderola, G.; Reinheimer, J.; Cuesta, I.; de los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Technological characterization and survival of the exopolysaccharide-producing strain Lactobacillus delbrueckii subsp. lactis 193 and its bile-resistant derivative 193+ in simulated gastric and intestinal juices. J. Dairy Res. 2011, 78, 357–364. [Google Scholar] [CrossRef] [Green Version]
- De Castro, C.; Parrilli, M.; Holst, O.; Molinaro, A. Microbe-associated molecular patterns in innate immunity: Extraction and chemical analysis of gram-negative bacterial lipopolysaccharides. Method. Enzymol. 2010, 480, 89–115. [Google Scholar]
- Ruas-Madiedo, P.; Zoon, P. Effect of exopolysaccharide-producing Lactococcus lactis strains and temperature on the permeability of skim milk gels. Colloids Surf. A Physicochem. Eng. Asp. 2003, 213, 245–253. [Google Scholar] [CrossRef]
- Lee, W.J.; Lucey, J.A. Formation and physical properties of yogurt. Asian-Australas. J. Anim. Sci. 2010, 23, 1127–1136. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; de los Reyes-Gavilán, C.G. Methods for the screening, isolation, and characterization of exopolysaccharides produced by lactic acid bacteria. J. Dairy Sci. 2005, 88, 843–856. [Google Scholar] [CrossRef] [Green Version]
- Dohnalkova, A.C.; Marshall, M.J.; Arey, B.W.; Williams, K.H.; Buck, E.C.; Fredrickson, J.K. Imaging hydrated microbial extracellular polymers: Comparative analysis by electron microscopy. Appl. Environ. Microbiol. 2011, 77, 1254–1262. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.; Li, F.; Li, L.; Huang, L.; Li, Q. Genetic and biochemical characterization of an exopolysaccharide with in vitro antitumoral activity produced by Lactobacillus fermentum YL-11. Front. Microbiol. 2019, 10, 2898. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Niu, M.; Song, D.; Song, X.; Zhao, J.; Wu, Y.; Lu, B.; Niu, G. Preparation, partial characterization and biological activity of exopolysaccharides produced from Lactobacillus fermentum S1. J. Biosci. Bioeng. 2020, 129, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, L.; Giraffa, G. Rapid identification of dairy lactic acid bacteria by M13-generated, RAPD-PCR fingerprint databases. J. Microbiol. Methods 2005, 63, 135–144. [Google Scholar] [CrossRef]
- Choi, S.; Jin, G.D.; Park, J.; You, I.; Kim, E.B. Pan-Genomics of Lactobacillus plantarum revealed group-specific genomic profiles without habitat association. J. Microbiol. Biotechnol. 2018, 28, 1352–1359. [Google Scholar] [CrossRef]
- Evanovich, E.; Mendonça Mattos, P.J.S.; Guerreiro, J.F. Comparative Genomic analysis of Lactobacillus plantarum: An overview. Int. J. Genom. 2019, 4973214. [Google Scholar] [CrossRef] [Green Version]
- Jia, F.F.; Zhang, L.J.; Pang, X.H.; Gua, X.X.; Abdelazez, A.; Liang, Y.; Sun, S.R.; Meng, X.C. Complete genome sequence of bacteriocin-producing Lactobacillus plantarum KLDS1.0391, a probiotic strain with gastrointestinal tract resistance and adhesion to the intestinal epithelial cells. Genomics 2017, 109, 432–437. [Google Scholar] [CrossRef]
- Das, D.; Goyal, A. Characterization and biocompatibility of glucan: A safe food additive from probiotic Lactobacillus plantarum DM5. J. Sci. Food Agric. 2014, 94, 683–690. [Google Scholar] [CrossRef]
- Gangoiti, M.V.; Puertas, A.I.; Hamet, M.F.; Peruzzo, P.J.; Llamas, M.G.; Medrano, M.; Prieto, A.; Dueñas, M.T.; Abraham, A.G. Lactobacillus plantarum CIDCA 8327: An α-glucan producing-strainisolated from kefir grains. Carbohydr. Polym. 2017, 170, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Yang, Z. A functional and genetic overview of exopolysaccharides produced by Lactobacillus plantarum. J. Funct. Foods 2018, 47, 229–240. [Google Scholar] [CrossRef]
- Ryan, P.M.; Ross, R.P.; Fitzgerald, G.F.; Caplicee, N.M.; Stanton, C. Sugar-coated: Exopolysaccharide producing lactic acid bacteria for food and human health applications. Food Funct. 2015, 6, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Vastano, V.; Perrone, F.; Marasco, R.; Sacco, M.; Muscariello, L. Transcriptional analysis of exopolysaccharide biosynthesis gene clusters in Lactobacillus plantarum. Arch. Microbiol. 2016, 198, 295–300. [Google Scholar] [CrossRef]
- Lee, I.C.; Caggianiello, G.; van Swam, I.I.; Taverne, N.; Meijerink, M.; Bron, P.A.; Spano, G.; Kleerebezema, M. Strain-specific features of extracellular polysaccharides and their impact on Lactobacillus plantarum-host interactions. Appl. Environ. Microbiol. 2016, 82, 3959–3970. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, M.; Muto, M.; Yokokura, T.; Mutai, M. Structure of 6-deoxytalose-containing polysaccharide from the cell wall of Bifidobacterium adolescentis. J. Biochem. 1988, 103, 618–621. [Google Scholar] [CrossRef]
- Vinogradov, E.; Sadovskaya, I.; Grard, T.; Chapot-Chartier, M.P. Structural studies of the rhamnose-rich cell wall polysaccharide of Lactobacillus casei BL23. Carboh. Res. 2016, 435, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vello, P.; Sharma, G.; Speciale, I.; Molinaro, A.; Im, S.-H.; De Castro, C. Structural features and immunological perception of the cell surface glycans of Lactobacillus plantarum: A novel rhamnose-rich polysaccharide and teichoic acids. Carbohydr. Polym. 2020, 233, 115857. [Google Scholar] [CrossRef]
- Degeest, B.; Vaningelgem, F.; De Vuyst, L. Microbial physiology, fermentation kinetics, and process engineering of heteropolysaccharide production by lactic acid bacteria. Int. Dairy J. 2001, 11, 747–757. [Google Scholar] [CrossRef]
- Laws, A.P.; Leivers, S.; Chacon-Romero, M.; Chadha, M.J. Variation in the molecular mass of exopolysaccharides during the time course of extended fermentations of skimmed milk by lactic acid bacteria. Int. Dairy J. 2009, 19, 768–771. [Google Scholar] [CrossRef]
- Zivkovic, M.; Miljkovic, M.; Ruas-Madiedo, P.; Strahinic, I.; Tolinacki, M.; Golic, N.; Kojic, M. Exopolysaccharide production and ropy phenotype are determined by two gene clusters in putative probiotic strain Lactobacillus paraplantarum BGCG11. Appl. Environ. Microbiol. 2015, 81, 1387–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Casteren, W.H.M.; Dijkem, C.; Schols, H.A.; Beldman, G.; Voragen, A.G.J. Characterisation and modification of the exopolysaccharide produced by Lactococcus lactis subsp. cremoris B40. Carbohydr. Polym. 1998, 37, 123–130. [Google Scholar] [CrossRef]
- Uemura, J.; Itoh, T.; Kasneko, T.; Noda, K. Chemical characterization of extracellular polysaccharide from Lactobacillus belbrueckii subsp. bulgaricus OLL1073R-1. Milchwissenschaft 1998, 53, 443–446. [Google Scholar]
- Sakellaris, G.; Kolisis, F.N.; Evangelopoulos, A.E. Presence of sialic acids in Lactobacillus plantarum. Biochem. Biophys. Res. Commun. 1988, 155, 126–1132. [Google Scholar] [CrossRef]
- Senturk, D.Z.; Dertli, E.; Erten, H.; Simsek, O. Structural and technological characterization of ropy exopolysaccharides produced by Lactobacillus plantarum strains isolated from Tarhana. Food Sci. Biotechnol. 2020, 29, 121–129. [Google Scholar] [CrossRef]
- Tallon, R.; Bressollier, P.; Urdaci, M.C. Isolation and characterization of two exopolysaccharides produced by Lactobacillus plantarum EP56. Res. Microbiol. 2003, 154, 705–712. [Google Scholar] [CrossRef]
- Liu, T.; Zhou, K.; Yin, S.; Liu, S.; Zhu, Y.; Yang, Y.; Wang, C. Purification and characterization of an exopolysaccharide produced by Lactobacillus plantarum HY isolated from home-made Sichuan Pickle. Int. J. Biol. Macromol. 2019, 134, 516–526. [Google Scholar] [CrossRef]
- Mende, S.; Rohm, H.; Jaros, D. Influence of exopolysaccharides on the structure, texture, stability and sensory properties of yoghurt and related products. Int. Dairy J. 2016, 52, 57–71. [Google Scholar] [CrossRef]
- Vukic, D.V.; Vukic, V.R.; Milanovic, S.D.; Ilicic, M.D.; Kanuric, K.G. Modeling of rheological characteristics of the fermented dairy products obtained by novel and traditional starter cultures. J. Food Sci. Technol. 2018, 55, 2180–2188. [Google Scholar] [CrossRef]
- Bancalari, E.; Alinovi, M.; Bottari, B.; Caligiani, A.; Mucchetti, G.; Gatti, M. Ability of a wild Weissella strain to modify viscosity of fermented milk. Front. Microbiol. 2020, 10, 3086. [Google Scholar] [CrossRef] [PubMed]
- Radhouani, H.; Gonçalves, C.; Maia, F.R.; Oliveira, J.M.; Reis, R.L. Kefiran biopolymer: Evaluation of its physicochemical and biological properties. J. Bioact. Compat. Polym. 2018, 33, 461–478. [Google Scholar] [CrossRef]
- Hamet, M.F.; Piermaria, J.A.; Abraham, A.G. Selection of EPS-producing Lactobacillus strains isolated from kefir grains and rheological characterization of the fermented milks. LWT Food Sci. Technol. 2015, 63, 129–135. [Google Scholar] [CrossRef]
- Surber, G.; Mende, S.; Jaros, D.; Rohm, H. Clustering of Streptococcus thermophilus strains to establish a relation between exopolysaccharide characteristics and gel properties of acidified milk. Foods 2019, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentès, M.C.; St-Gelais, D.; Turgeon, S.L. Exopolysaccharide–milk protein interactions in a dairy model system simulating yoghurt conditions. Dairy Sci. Technol. 2013, 93, 255–271. [Google Scholar] [CrossRef] [Green Version]
- Lucey, J.A. Acid coagulation of milk. In Advanced Dairy Chemistry-1B: Proteins: Applied Aspects, 4th ed.; McSweeney, P.L.H., O’Mahony, J.A., Eds.; Springer International Publishing: New York, NY, USA, 2016; Chapter 12; pp. 309–328. [Google Scholar]
- Lluis-Arroyo, D.; Flores-Nájera, A.; Cruz-Guerrero, A.; Gallardo-Escamilla1, F.; Lobato-Calleros, C.; Jiménez-Guzmán, J.; García-Garibay, M. Effect of an exopolysaccharide-producing strain of Streptococcus thermophilus on the yield and texture of Mexican Manchego-type cheese. Int. J. Food Prop. 2014, 17, 1680–1693. [Google Scholar] [CrossRef]
- Girard, M.; Schaffer-Lequart, C. Gelation and resistance to shearing of fermented milk: Role of exopolysaccharides. Int. Dairy J. 2007, 17, 666–673. [Google Scholar] [CrossRef]
- Gentès, M.C.; St-Gelais, D.; Turgeon, S.L. Gel formation and rheological properties of fermented milk with in situ exopolysaccharide production by lactic acid bacteria. Dairy Sci. Technol. 2011, 91, 645–661. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Wu, K.; Wang, F.; Liang, X.; Liu, Q.; Li, G.; Li, Q. Effect of exopolysaccharides from lactic acid bacteria on the texture and microstructure of buffalo yoghurt. Int. Dairy J. 2014, 34, 252–256. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genome | ||||||
|---|---|---|---|---|---|---|
| Accession No. | Length (bp) | CDSs | %G + C | rRNA | tRNA | |
| LBIO1 (Ropy) | SAMN14671341 | 3,186,843 | 3113 | 44.32 | 9 | 66 |
| LBIO14 (non-ropy) | SAMN14671394 | 3,155,118 | 3072 | 44.44 | 10 | 68 |
| LBIO28 (Ropy) | SAMN14671401 | 3,187, 061 | 3108 | 44.32 | 10 | 66 |
| Mean ± Standard Deviation (SD) | ||||
|---|---|---|---|---|
| Strain | Parameters | Peak 1 | Peak 2 | Peak 3 |
| LBIO1 (Ropy) | Retention time (min) | 26.83 ± 0.01 | 37.61 ± 7.40 | 41.52 ± 0.01 |
| Mw (× 103 g/mol) | 1154.5 ± 16.26 | 19.9 ± 0.07 | 4.41 ± 0.47 | |
| Rw (nm) | 74.6 ± 2.26 | 34.8 ± 11.9 | 68.9 ± 1.34 | |
| % relative abundance | 15.8 ± 0.1 | 15.3 ± 0.2 | 68.2 ± 0.2 | |
| LBIO14 (No-ropy) | Retention time (min) | ND | 37.80 ± 0.01 | 41.54 ± 0.01 |
| Mw (× 103 g/mol) | 20.3 ± 1.57 | 8.81 ± 3.61 | ||
| Rw (nm) | 39.0 ± 8.9 | 137.1 ± 42.0 | ||
| % relative abundance | 14.6 ± 0.0 | 85.0 ± 0.1 | ||
| LBIO28 (Ropy) | Retention time (min) | 26.73 ± 0.01 | 37.63 ± 0.01 | 41.53 ± 0.01 |
| Mw (× 103 g/mol) | 1183.5 ± 43.13 | 22.4 ± 0.08 | 9.90 ± 0.11 | |
| Rw (nm) | 74.7 ± 0.50 | 45.9 ± 0.7 | 143.1 ± 0.56 | |
| % relative abundance | 15.9 ± 0.5 | 15.2 ± 0.2 | 68.0 ± 0.3 | |
| Monomer 1 Ratio/(%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| EPSs | Glycerol | Ribitol | Man | Gal | Glc | GalN | GlcN | NeuA |
| LBIO1 | 0.06 (3.2) | 0.19 (10.1) | 0.07 (3.7) | 0.38 (20.1) | 1.00 (52.9) | 0.05 (2.6) | 0.07 (3.7) | 0.07 (3.7) |
| LBIO14 | 0.12 (5.4) | 0.31 (13.8) | 0.14 (6.3) | 0.33 (14.7) | 1.00 (44.6) | 0.09 (4.0) | 0.12 (5.4) | 0.13 (5.8) |
| LBIO28 | 0.06 (3.4) | 0.16 (8.9) | 0.08 (4.5) | 0.36 (20.1) | 1.00 (55.9) | 0.03 (1.7) | 0.05 (2.8) | 0.05 (2.8) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachtarzi, N.; Speciale, I.; Kharroub, K.; De Castro, C.; Ruiz, L.; Ruas-Madiedo, P. Selection of Exopolysaccharide-Producing Lactobacillus Plantarum (Lactiplantibacillus Plantarum) Isolated from Algerian Fermented Foods for the Manufacture of Skim-Milk Fermented Products. Microorganisms 2020, 8, 1101. https://doi.org/10.3390/microorganisms8081101
Bachtarzi N, Speciale I, Kharroub K, De Castro C, Ruiz L, Ruas-Madiedo P. Selection of Exopolysaccharide-Producing Lactobacillus Plantarum (Lactiplantibacillus Plantarum) Isolated from Algerian Fermented Foods for the Manufacture of Skim-Milk Fermented Products. Microorganisms. 2020; 8(8):1101. https://doi.org/10.3390/microorganisms8081101
Chicago/Turabian StyleBachtarzi, Nadia, Immacolata Speciale, Karima Kharroub, Cristina De Castro, Lorena Ruiz, and Patricia Ruas-Madiedo. 2020. "Selection of Exopolysaccharide-Producing Lactobacillus Plantarum (Lactiplantibacillus Plantarum) Isolated from Algerian Fermented Foods for the Manufacture of Skim-Milk Fermented Products" Microorganisms 8, no. 8: 1101. https://doi.org/10.3390/microorganisms8081101