1. Introduction
The gastrointestinal microbiota largely relies on the host animal for the supply of nutrients. Depending on the bacterial substrate preferences, the microbiota compete with the host for easily digestible nutrients (e.g., starch and protein), whereas by fermenting dietary fiber the microbes convert a non-usable dietary component into absorbable nutrients in the form of short-chain fatty acids (SCFA) [
1]. Besides macronutrients, the gut microbiota rely on mineral supply originating from the diet or intestinal secretions and largely compete with the host if the provision becomes low [
2]. Especially phosphorus (P) is an essential macroelement for the gut microbiota; bacteria will downregulate cell replication and metabolism, if this mineral becomes deficient [
3]. For this reason, sufficient supply of the rumen microbiota with P has long been recognized as crucial in order not to compromise rumen fiber fermentation and SCFA production [
3]. Evidence is accumulating that the gastrointestinal P availability affects the gut microbiome composition in monogastric livestock species as well, thereby affecting the intestinal abundance of opportunistic pathogens [
4,
5]. While mammals cannot utilize phytate-P, the major storage form of P in plants, gut bacteria comprise the necessary enzymatic capacities [
6]. However, gut microbes only turn on this machinery when P in their surroundings becomes very low [
6]. Due to ecological and economic reasons, dietary levels of inorganic P have been reduced in pig production [
7] and different feed technological measures are taken to improve the availability of phytate-P for monogastric farm animals, modifying the microbial availability of P along the gastrointestinal tract [
8,
9]. The most common strategy is the addition of microbial phytase to diets for pigs, for which we could recently show to have a large impact on the ileal and fecal bacterial communities in weaned pigs, affecting the most abundant taxa (i.e.,
Clostridiaceae and
Lactobacillaceae), as well as pathobionts, like
Helicobacter,
Campylobacter and
Fusobacterium [
9,
10].
It is often assumed that the maturational changes in the gastrointestinal microbiome become less pronounced the more time elapses after weaning and that the microbiome composition is more affected by the diet. Nevertheless, age-related changes are still present [
11] possibly influencing the response of the intestinal microbiota to dietary changes. Dietary responses of the gut microbiota in growing pigs are often only assessed at one time point, typically at the termination of the experiment. If the gut microbiota progressively changes throughout the fattening period, it can be assumed that the microbial responses to diets may change over time. Since intestinal digesta and feces are a potential reservoir of food-borne pathogens for pork contamination during the slaughtering process [
12,
13], age-related differences in dietary effects on the gut bacterial composition are valid to investigate in order to optimize dietary formulations and reduce pathogen transmission. We therefore hypothesized that maturational changes in the microbiota composition would alter the bacterial response to a diet supplemented with phytase and hence to changes in the intestinally available P when comparing the microbiota composition in the early, mid and late fattening period. Furthermore, reports on different ‘community types’ in pig populations [
14] also led us assume that inter-individual differences in the gut microbiota composition may alter the microbial responses towards the applied phytase supplementation and intestinal P availability. Although feces may not be the suitable material in all situations, they allow multiple samplings in one animal over time, thereby reducing the inter-animal variability. In this study, we used phytase supplementation to alter the P availability in the hindgut and investigated the dietary response of the fecal bacterial microbiome from the early to late fattening period.
4. Discussion
Albeit maturational changes in the porcine gut microbiome become smaller the more time passes after weaning [
11,
27], present results demonstrate significant fluctuations in the fecal microbiota composition and diversity from the early to late fattening period. Corresponding to our findings in young growing pigs [
9], phytase affected, positively or negatively, all major bacterial genera in the present study, representing approximately 50% of all reads. This strongly supports the importance of P for bacterial metabolism [
28]. By assuming a similar situation in the large intestine, this may have impacted the degradation of feed residuals and SCFA production as well as may have altered the innate immune response in the distal large intestine [
29]. Our results also showed that the phytase-associated alterations in the fecal microbiome composition differed over time, indicating that the observed maturational changes modified the response of the fecal microbiome to the dietary manipulation.
Although the representativeness of the fecal community for the porcine gut microbiota is often questioned, monitoring the bacterial community over time and thus multiple samplings on the same (intact) animal can only be achieved when using feces as the gut material of choice [
27]. The bacterial microbiome composition is qualitatively similar to those in the distal parts of the large intestine [
11]. Hence, fecal microbiome data add valid information to our understanding of nutrient–microbiota interactions. Our data also provide useful information in that pigs are housed in a closed environment (pens), where they take up feces while playing or deliberately, which contributes to bacterial (pathogen) transmission between animals. Likewise, fecal smears on the skin of the pig are a potential source for carcass contamination during the slaughtering process [
13].
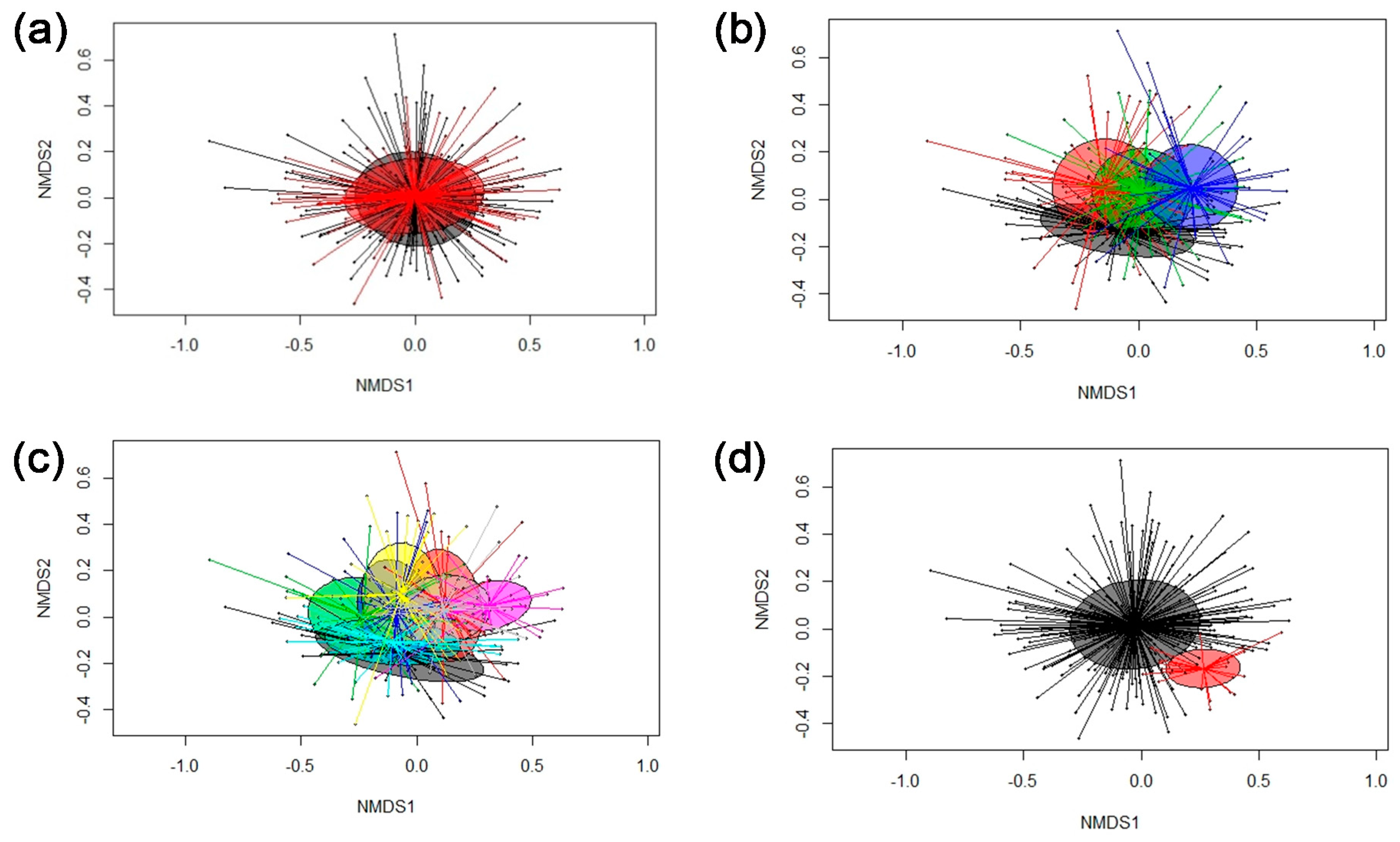
Inter-individual differences in the gut microbiome can reduce the chance to distinguish dietary effects on the gut community composition in a pig population [
30]. Certain pen and hence litter-associated differences were noticeable in the fecal microbiomes throughout the course of the experiment according to the PERMANOVA, which may be related, for instance, to a maternal effect or differences in the micro-environment ‘pen’ [
31]. Contrarily, the two identified ‘community types’ were not litter related. More interestingly, these differences in the bacterial community composition mainly existed at the start of the experiment and disappeared thereafter, supporting the strong impact of the diet on the gut microbiome composition [
19,
20,
32,
33]. Since the fecal microbiome composition on the second experimental day (week 0) was mainly affected by the conditions prior to the experiment, the cause for these differences remains difficult to trace back as pigs were similarly housed and treated (including feeding) from birth and kept together in litter groups postweaning. Potential reasons for the observed differences in the fecal microbiome structure might be pigs’ intake of colostrum, sow milk or solid feed pre- and postweaning. As one ‘community type’ comprised more
Prevotella,
Lactobacillus,
Ruminococcus and
Megasphaera, whereas the other was enriched in
Faecalibacterium and
Escherichia, differences in gut microbial–host networking can be assumed. Intestinal production of butyrate due to its multiple effects upon the host as energy source and upon apoptosis, inflammation, and oxidative stress is generally considered to be beneficial to gut health [
1]. Both ‘community types’ were enriched in butyrate producers; however,
Megasphaera and
Faecalibacterium rely on different substrates, supporting different microbial interactions. By including lactate utilizers, the genus
Megasphaera may have profited from cross-feeding relationships with
Lactobacillus [
1], whereas
Faecalibacterium may have utilized pectin-rich dietary residuals or host-derived
N-acetylglucosamine as substrate [
34].
Information is available about maturational changes in the porcine gut microbiome for the fattening compared to the pre- and early postweaning phase [
27,
35]. The bacterial diversity is commonly used as a measure for the stability of the gut microbiota, with a higher diversity representing a greater stability [
36]. Here, we found a decline in bacterial species richness (Chao1) and diversity in feces from week 11 of age (week 0 of the experiment) to week 14 of age (week 3 of the experiment), after which it remained at this level until week 21 of age (week 5 of the experiment). This was in contrast to previous observations where no changes [
37] or an increase in fecal diversity between 3 and 6 months of age [
11,
27] were reported. Since pigs did not show signs of enteric disease and had a high growth intensity in the present study, we can assume that this loss in species richness and evenness was in the physiological range, not affecting the stability of the present fecal microbiomes. At taxonomic level, especially the changes in the predominant bacterial genera within the very versatile families
Prevotellaceae,
Clostridiaceae and
Ruminococcaceae from week 0 to 10 point towards alterations in metabolic capabilities to degrade complex fibrous materials and protein with increasing age. Age-related shifts in the bacterial community likely changed cross-feeding relationships among bacteria [
1]. Notably,
Megasphaera abundances coincided with
Prevotella abundances in the present study, suggesting metabolite dependencies between these two taxa. Other taxa showed fluctuations over time, such as
Lactobacillus and
Coprococcus, which may have been related to competition for substrate and mutual promotion or inhibition by other bacteria. Some age-related alterations in the fecal community may have been linked to the higher feed intake with increasing BW of the pig, which was supported by relevance networking. However, more than 50% of the variation need to be explained by other factors.
Phytase improves the P availability and absorption in the upper digestive tract, leaving less P available for microbial metabolism in the large intestine [
8]. Although the present fecal microbiome composition differed from our previous study in the actual abundances [
9], similar taxa were affected, showing that genera within
Clostridiaceae and
Ruminococcaceae largely benefited from the lower P availability in the distal large intestine due to the phytase supplementation after 3 and 5 weeks on the diets. Many of these genera, except
SMB53 within
Clostridiaceae, were not assigned at genus or species level, making it difficult to deduce metabolic features for these genera and consequences for the host. From rumen research, evidence exists that especially taxa with cellulolytic and hemicellulolytic capabilities but less amylolytic bacteria reduce growth and metabolic activity with lower concentrations of P in their environment [
3]. Both families,
Clostridiaceae and
Ruminococcaceae, comprise amylolytic and pullulanolytic species [
20,
38], which may have tolerated the lower P availability in their environment better than other fibrolytic bacteria. By being a metabolically extremely versatile genus per se,
Prevotella is typically associated with a plant-rich diet and a microbiome enriched in this taxon has an increased potential to ferment complex polysaccharides [
39]. Due to the
Prevotella dominance in the present microbiomes, their phytase-related decline may have a stronger impact on degradation of hemicellulose and cellulose and microbe-to microbe and microbial–host interactions than lower abundant taxa. If a taxa with higher P requirement became less abundant due to maturation, more P may have been available for other taxa. This may help explaining that some taxa (e.g.,
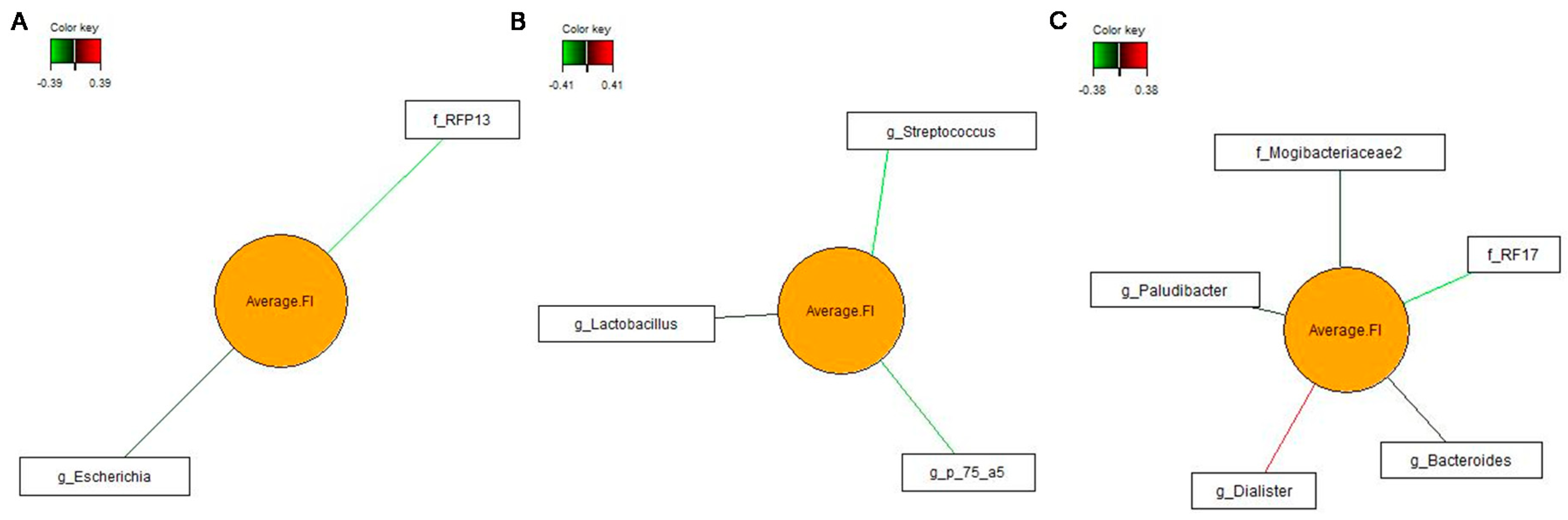
Lactobacillus, Streptococcus and
Turicibacter) were only affected at certain sampling time points but not at all. Also, maturational changes in host physiology and digestion during the fattening period may have contributed to the age-related responses of the fecal bacteria to the phytase. The same fattening diet was fed over the whole experimental period. Unfortunately, the apparent total tract digestibility was not determined in this study. The present results for the maturational changes in the fecal microbiome emphasize the need to include this measurement in future gut microbiome studies. However, it can be generally assumed that pigs received less nutrients than required in the early and more nutrients than required in the later stages of the fattening period [
16]. As a consequence, more P was absorbed in the small intestine, intensifying the P depletion in large intestinal digesta in the early compared to the later stages of the fattening period. Simultaneously, changes in other nutrients in digesta from the early fattening period to the later stages possibly altered the nutrient profile for microbial metabolism in the large intestine over time. This may explain, at least in part, the varying abundances of
Lactobacillus,
Streptococcus,
Dialister,
Turicibacter, Oscillospira, Blautia and
Christensenellaceae, which rely on different simple and complex carbohydrates, from week 3 to 10. Besides, phytase supplementation can increase the digestibility of cations, such as calcium [
40] and amino acids [
41], potentially improving their digestibility. Free Ca-ions bind to cell wall components of Gram-positive bacteria, including proteins, exopolysaccharides, and lipoteichoic acid [
42,
43], and hence can modify adhesive interactions with protein and polysaccharide adhesion molecules at the cell surface [
44]. Especially
Lactobacillus may have a certain requirement of free Ca ions for their attachment to the intestinal mucosa [
5,
45]. Therefore, lower intestinal Ca availability due to increased intestinal absorption of Ca with the phytase may help explaining, similar to our previous study [
9], the lower abundance of
Lactobacillus in feces of these pigs. Calcium and P, by forming an insoluble Ca-P complex at pH values above 5 [
46], as well as peptides contribute to buffering of the intestinal lumen. Since bacteria are susceptible to changes in the environmental pH, this may be another rationale having contributed to the phytase-associated alterations in the bacterial community. In a similar manner, the lower abundance of
Treponema, a genus with proteolytic abilities [
47], with phytase may link to a reduced large intestinal protein flow. Although a recent study could not find an effect of phytase on small intestinal mucin secretion [
48], the increased abundance of proteolytic and mucolytic taxa, such as
Clostridium and
Escherichia [
49], may hint at an increased glycoprotein utilization. Since genera like
Clostridium and
Escherichia comprise important porcine pathobionts, it seems valid to examine the effect of phytase on expression of virulence factors in functional approaches, which could not be covered by targeting the 16S rRNA gene sequencing approach.
In conclusion, the present results provide evidence that, during the fattening phase, bacterial responses to a dietary treatment change over time, emphasizing the need to investigate dietary effects in the short and long term. Whilst the bacterial phytase responses differed from the early to late fattening period, results largely support the importance of P for bacterial proliferation in the large intestine of pigs as the phytase supplementation affected the fecal abundances of all major genera in the present study. Moreover, cluster analysis identified two bacterial ‘community types’ at the start of the experiment, indicating the importance of screening for inter-individual differences in the gut microbiome when investigating bacterial dietary responses.





{kind=link}
{kind=link}