Human Metapneumovirus Establishes Persistent Infection in Lung Microvascular Endothelial Cells and Primes a Th2-Skewed Immune Response

 , , and
, , and 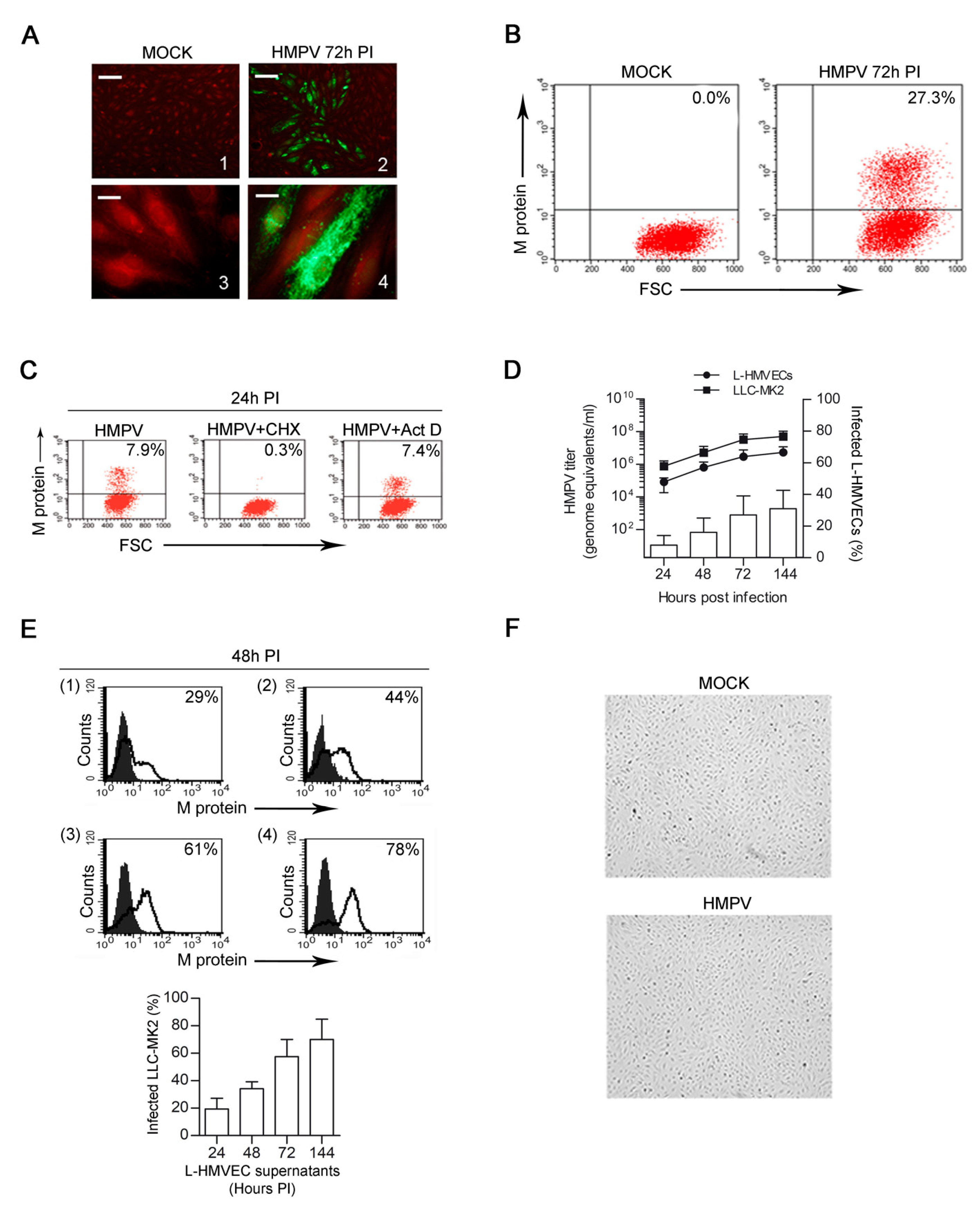{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. HMPV Production and Titration
2.2. Endothelial Cell Culture and Infection
2.3. Immunofluorescence Analysis
2.4. DC Generation
2.5. Coculture and Priming of Naïve T Cells
2.6. Flow Cytometry
2.7. Real-Time PCR
2.8. Thymic Stromal Lymphopoietin (TSLP) ELISA
2.9. Metagenomic Analysis of HMPV
2.10. Statistical Analysis
3. Results
3.1. HMPV Successfully Infects Human Lung Microvascular Endothelial Cells
3.2. HMPV Establishes a Persistent Productive Infection in L-HMVECs
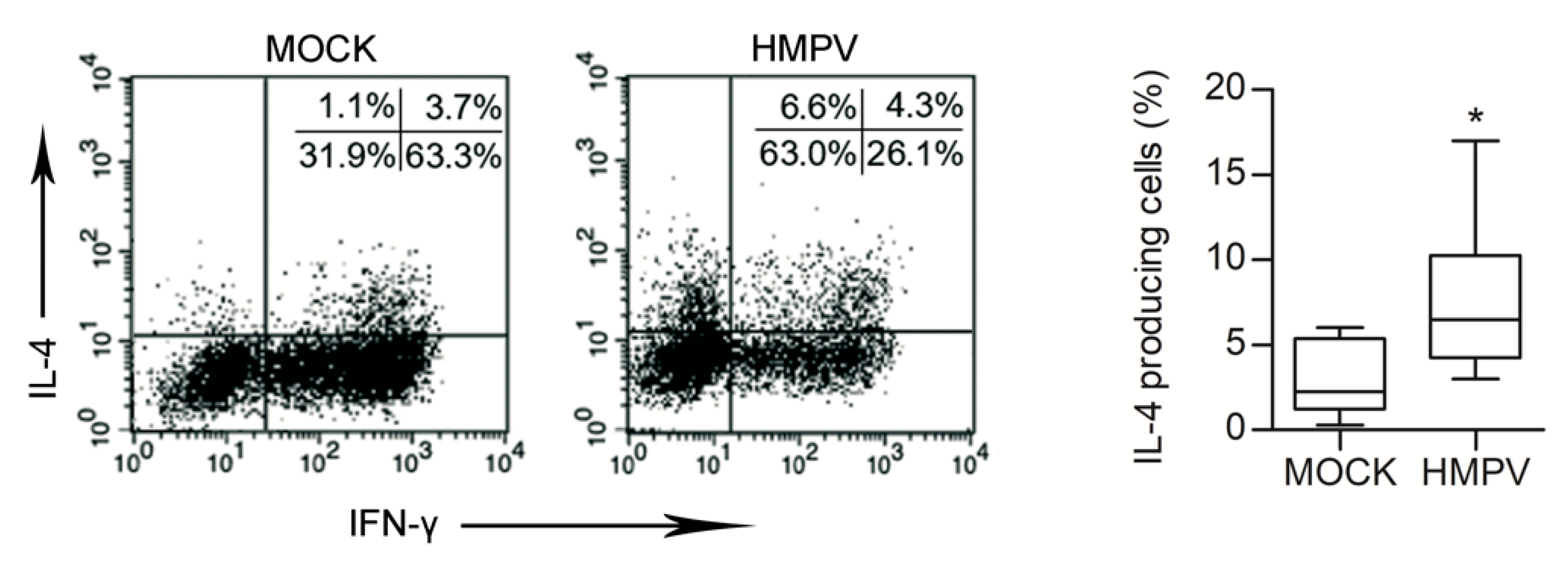
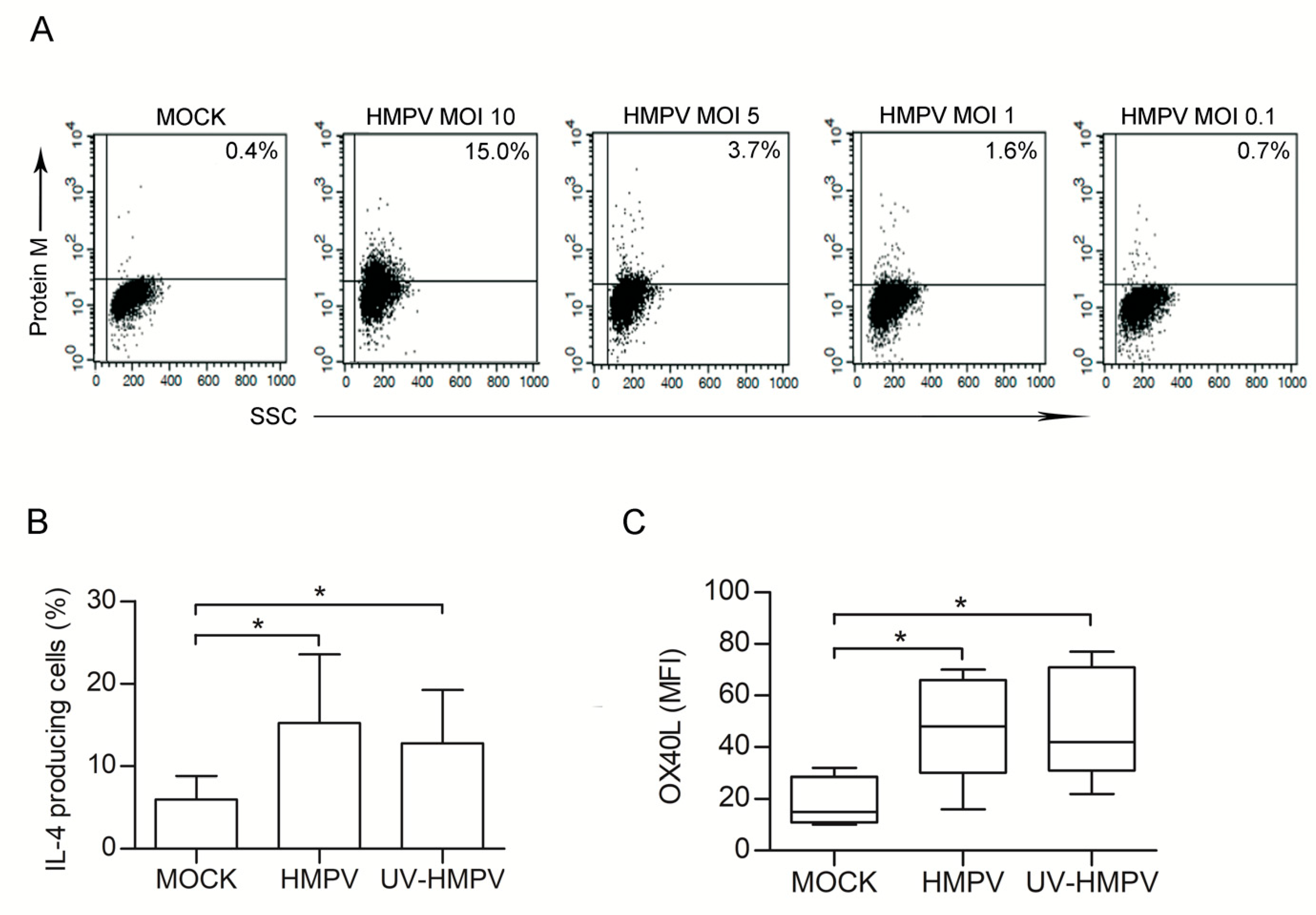
3.3. HMPV-Infected L-HMVEC Secretomes Induce DC to Promote a Th2 Phenotype
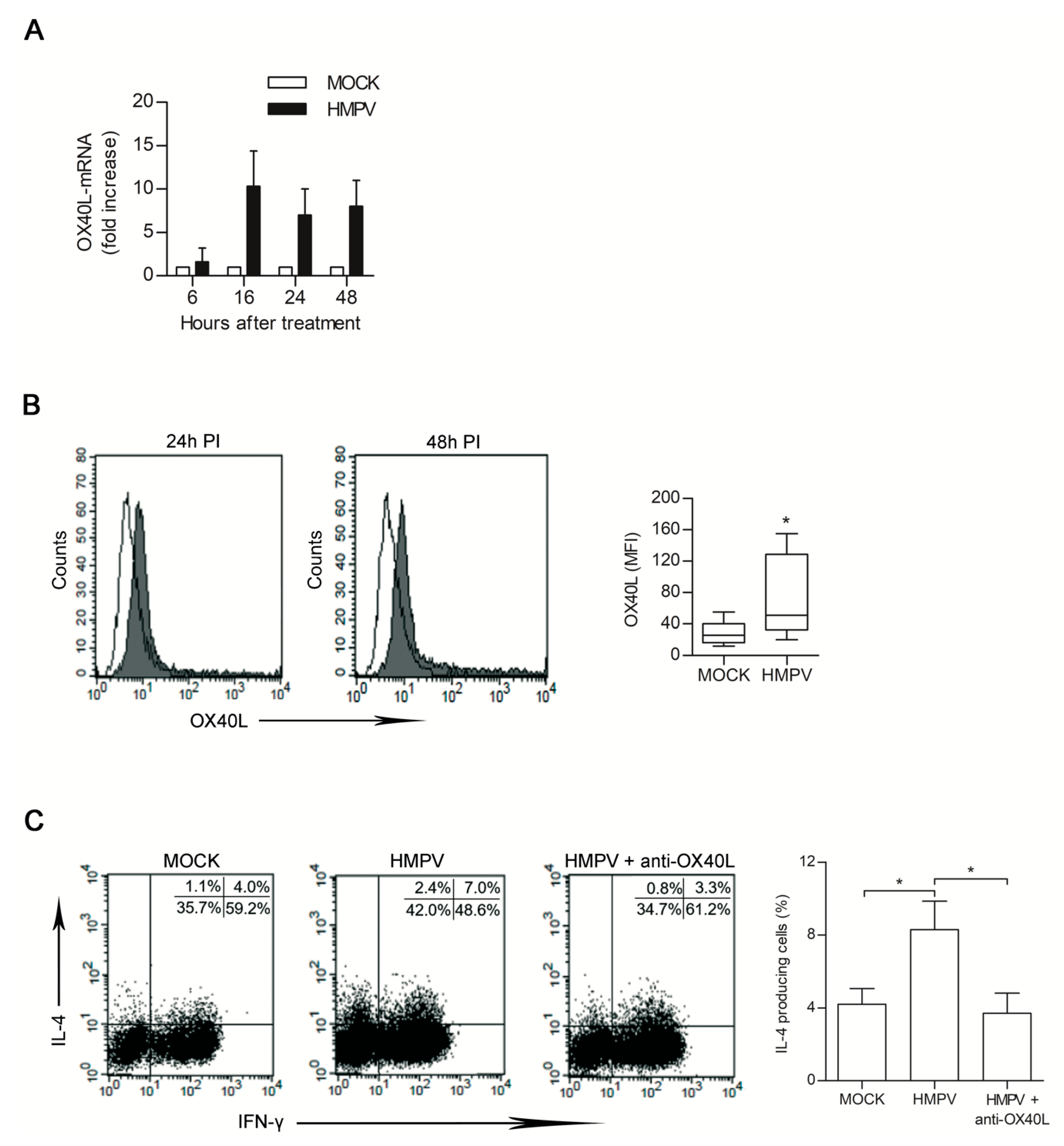
3.4. HMPV-Infected LHMVECs Secretome Induces OX40L Expression on DCs
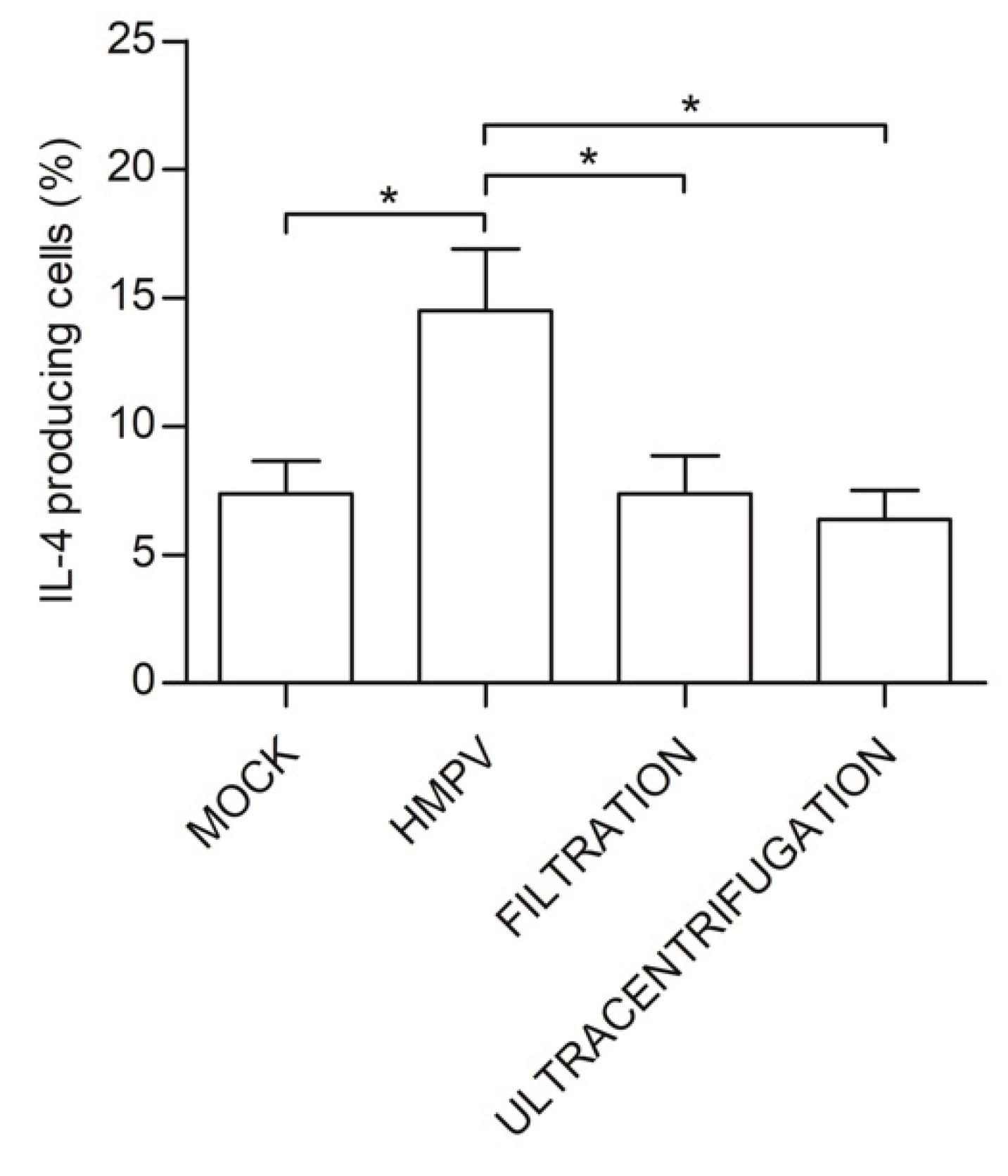
3.5. Th2 Polarization is Directly Induced by HMPV Viral Particles Contained in L-HMVEC Secretomes
3.6. HMPV Induces, per se, Th2 cell Polarization
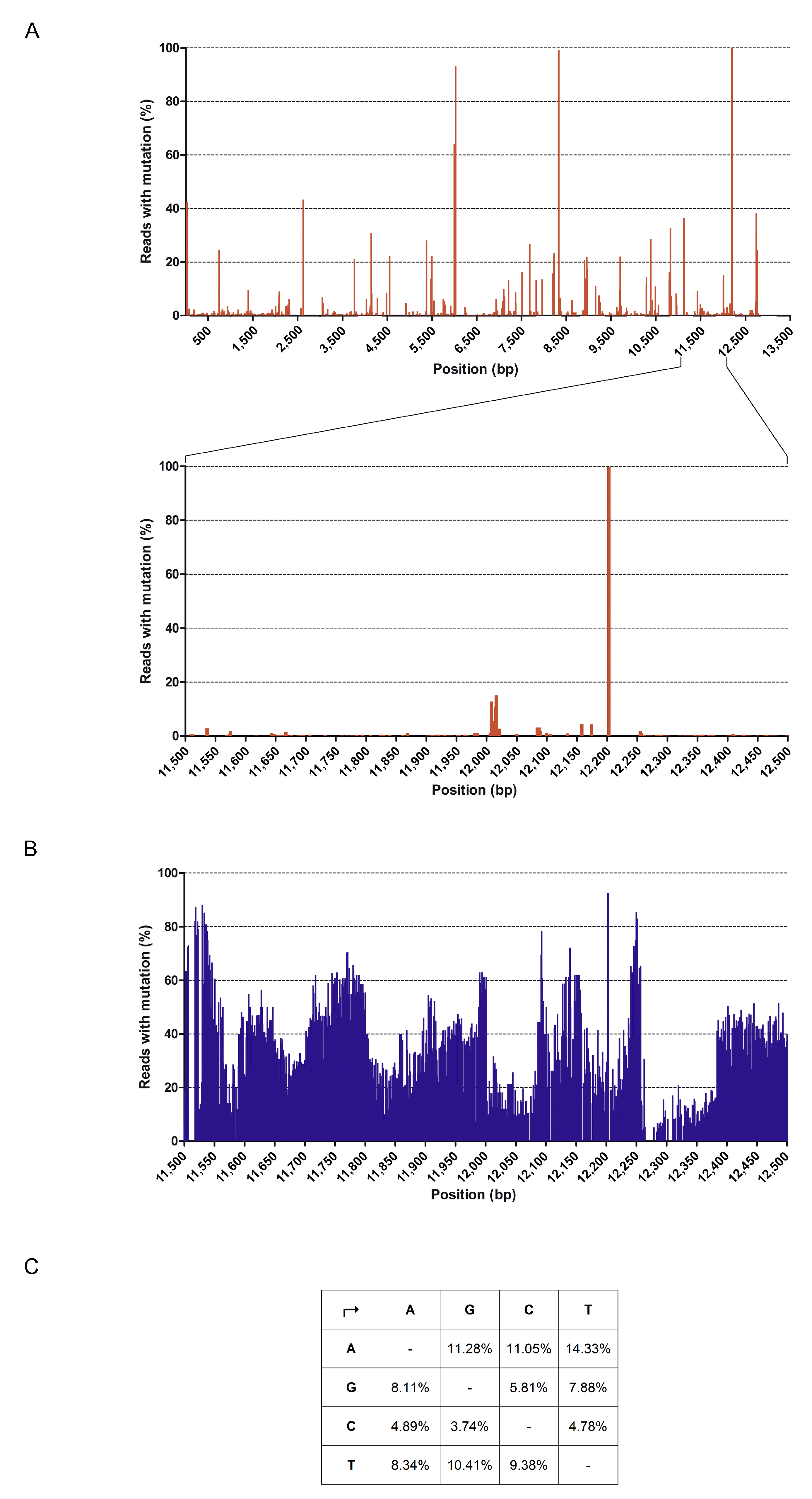
3.7. Full Genome Sequencing of HMPV
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Williams, J.V.; Wang, C.K.; Yang, C.F.; Tollefson, S.J.; House, F.S.; Heck, J.M.; Chu, M.; Brown, J.B.; Lintao, L.D.; Quinto, J.D.; et al. The role of human metapneumovirus in upper respiratory tract infections in children: A 20-year experience. J. Infect. Dis. 2006, 193, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Boivin, G.; Abed, Y.; Pelletier, G.; Ruel, L.; Moisan, D.; Cote, S.; Peret, T.C.; Erdman, D.D.; Anderson, L.J. Virological features and clinical manifestations associated with human metapneumovirus: A new paramyxovirus responsible for acute respiratory-tract infections in all age groups. J. Infect. Dis. 2002, 186, 1330–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkesmann, A.; Schildgen, O.; Eis-Hubinger, A.M.; Geikowski, T.; Glatzel, T.; Lentze, M.J.; Bode, U.; Simon, A. Human metapneumovirus infections cause similar symptoms and clinical severity as respiratory syncytial virus infections. Eur. J. Pediatr. 2006, 165, 467–475. [Google Scholar] [CrossRef]
- Caracciolo, S.; Minini, C.; Colombrita, D.; Rossi, D.; Miglietti, N.; Vettore, E.; Caruso, A.; Fiorentini, S. Human metapneumovirus infection in young children hospitalized with acute respiratory tract disease: Virologic and clinical features. Pediatr. Infect. Dis. J. 2008, 27, 406–412. [Google Scholar] [CrossRef]
- Apostoli, P.; Zicari, S.; Lo Presti, A.; Ciccozzi, M.; Ciotti, M.; Caruso, A.; Fiorentini, S. Human metapneumovirus-associated hospital admissions over five consecutive epidemic seasons: Evidence for alternating circulation of different genotypes. J. Med. Virol. 2012, 84, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Ebihara, T.; Endo, R.; Kikuta, H.; Ishiguro, N.; Yoshioka, M.; Ma, X.; Kobayashi, K. Seroprevalence of human metapneumovirus in Japan. J. Med. Virol. 2003, 70, 281–283. [Google Scholar] [CrossRef]
- Biacchesi, S.; Pham, Q.N.; Skiadopoulos, M.H.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Modification of the trypsin-dependent cleavage activation site of the human metapneumovirus fusion protein to be trypsin independent does not increase replication or spread in rodents or nonhuman primates. J. Virol. 2006, 80, 5798–5806. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, R.; Tripp, R.A. The immune response to human metapneumovirus is associated with aberrant immunity and impaired virus clearance in BALB/c mice. J. Virol. 2005, 79, 5971–5978. [Google Scholar] [CrossRef] [Green Version]
- Hammad, H.; Lambrecht, B.N. Dendritic cells and epithelial cells: Linking innate and adaptive immunity in asthma. Nat. Rev. Immunol. 2008, 8, 193–204. [Google Scholar] [CrossRef]
- García-García, M.L.; Calvo, C.; Casas, I.; Bracamonte, T.; Rellán, A.; Gozalo, F.; Tenorio, T.; Pérez-Breña, P. Human metapneumovirus bronchiolitis in infancy is an important risk factor for asthma at age 5. Pediatr. Pulmonol. 2007, 42, 458–464. [Google Scholar] [CrossRef]
- Melendi, G.A.; Laham, F.R.; Monsalvo, A.C.; Casellas, J.M.; Israele, V.; Polack, N.R.; Kleeberger, S.R.; Polack, F.P. Cytokine profiles in the respiratory tract during primary infection with human metapneumovirus, respiratory syncytial virus, or influenza virus in infants. Pediatrics 2007, 120, e410–e415. [Google Scholar] [CrossRef] [PubMed]
- Laham, F.R.; Israele, V.; Casellas, J.M.; Garcia, A.M.; Lac Prugent, C.M.; Hoffman, S.J.; Hauer, D.; Thumar, B.; Name, M.I.; Pascual, A.; et al. Differential production of inflammatory cytokines in primary infection with human metapneumovirus and with other common respiratory viruses of infancy. J. Infect. Dis. 2004, 189, 2047–2056. [Google Scholar] [CrossRef] [PubMed]
- Douville, R.N.; Bastien, N.; Li, Y.; Pochard, P.; Simons, F.E.; HayGlass, K.T. Human metapneumovirus elicits weak IFN-gamma memory responses compared with respiratory syncytial virus. J. Immunol. 2006, 176, 5848–5855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, A.E.; Lay, M.K.; Jara, E.L.; Espinoza, J.A.; Gómez, R.S.; Soto, J.; Rivera, C.A.; Abarca, K.; Bueno, S.M.; Riedel, C.A. Aberrant T cell immunity triggered by human Respiratory Syncytial Virus and human Metapneumovirus infection. Virulence 2017, 8, 685–704. [Google Scholar] [CrossRef]
- Hamelin, M.E.; Prince, G.A.; Gomez, A.M.; Kinkead, R.; Boivin, G. Human metapneumovirus infection induces long-term pulmonary inflammation associated with airway obstruction and hyperresponsiveness in mice. J. Infect. Dis. 2006, 193, 1634–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolli, D.; Bataki, E.L.; Spetch, L.; Guerrero-Plata, A.; Jewell, A.M.; Piedra, P.A.; Milligan, G.N.; Garofalo, R.P.; Casola, A. T lymphocytes contribute to antiviral immunity and pathogenesis in experimental human metapneumovirus infection. J. Virol. 2008, 82, 8560–8569. [Google Scholar] [CrossRef] [Green Version]
- Lay, M.K.; Céspedes, P.F.; Palavecino, C.E.; León, M.A.; Díaz, R.A.; Salazar, F.J.; Méndez, G.P.; Bueno, S.M.; Kalergis, A.M. Human metapneumovirus infection activates the TSLP pathway that drives excessive pulmonary inflammation and viral replication in mice. Eur. J. Immunol. 2015, 6, 1680–1695. [Google Scholar] [CrossRef]
- Hamelin, M.E.; Couture, C.; Sackett, M.K.; Boivin, G. Enhanced lung disease and Th2 response following human metapneumovirus infection in mice immunized with the inactivated virus. J. Gen. Virol. 2007, 88, 3391–3400. [Google Scholar] [CrossRef]
- Alvarez, R.; Harrod, K.S.; Shieh, W.J.; Zaki, S.; Tripp, R.A. Human metapneumovirus persists in BALB/c mice despite the presence of neutralizing antibodies. J. Virol. 2004, 78, 14003–14011. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Haas, D.L.; Poore, S.; Isakovic, S.; Gahan, M.; Mahalingam, S.; Fu, Z.F.; Tripp, R.A. Human metapneumovirus establishes persistent infection in the lungs of mice and is reactivated by glucocorticoid treatment. J. Virol. 2009, 83, 6837–6848. [Google Scholar] [CrossRef] [Green Version]
- Marsico, S.; Caccuri, F.; Mazzuca, P.; Apostoli, P.; Roversi, S.; Lorenzin, G.; Zani, A.; Fiorentini, S.; Giagulli, C.; Caruso, A. Human lung epithelial cells support human metapneumovirus persistence by overcoming apoptosis. Pathog. Dis. 2018, 76, fty013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, R.; König, W. Respiratory syncytial virus infection of human lung endothelial cells enhances selectively intercellular adhesion molecule-1 expression. J. Immunol. 2005, 174, 7359–7367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teijaro, J.R.; Walsh, K.B.; Cahalan, S.; Fremgen, D.M.; Roberts, E.; Scott, F.; Martinborough, E.; Peach, R.; Oldstone, M.B.; Rosen, H. Endothelial cells are central orchestrators of cytokine amplification during influenza virus infection. Cell 2011, 146, 980–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perelygina, L.; Zheng, Q.; Metcalfe, M.; Icenogle, J. Persistent infection of human fetal endothelial cells with rubella virus. PLoS ONE 2013, 8, e73014. [Google Scholar] [CrossRef] [Green Version]
- Mladinich, M.C.; Schwedes, J.; Mackowa, E.R. Zika virus persistently infects and is basolaterally released from primary human brain microvascular endotheliall cells. MBio 2017, 8, e00952. [Google Scholar] [CrossRef] [Green Version]
- An, P.; Robles, E.; Duray, A.M.; Cantalupo, P.G.; Pipas, J.M. Human polyomavirus BKV infection of endothelial cells results in interferon pathway induction and persistence. PLoS Pathog. 2019, 15, e1007505. [Google Scholar] [CrossRef]
- Biacchesi, S.; Skiadopoulos, M.H.; Yang, L.; Lamirande, E.W.; Tran, K.C.; Murphy, B.R.; Collins, P.L.; Buchholz, U.J. Recombinant human Metapneumovirus lacking the small hydrophobic SH and/or attachment G glycoprotein: Deletion of G yields a promising vaccine candidate. J. Virol. 2004, 78, 12877–12887. [Google Scholar] [CrossRef] [Green Version]
- Mazzuca, P.; Marsico, S.; Schulze, K.; Mitola, S.; Pils, M.C.; Giagulli, C.; Guzman, C.A.; Caruso, A.; Caccuri, F. Role of Autophagy in HIV-1 Matrix Protein p17-Driven Lymphangiogenesis. J. Virol. 2017, 91, e00801. [Google Scholar] [CrossRef] [Green Version]
- Seeger, P.; Bosisio, D.; Parolini, S.; Badolato, R.; Gismondi, A.; Santoni, A.; Sozzani, S. Activin A as a mediator of NK-dendritic cell functional interactions. J. Immunol. 2014, 192, 1241–1248. [Google Scholar] [CrossRef] [Green Version]
- Krause, P.; Bruckner, M.; Uermösi, C.; Singer, E.; Groettrup, M.; Legler, D.F. Prostaglandin E2 enhances T-cell proliferation by inducing the costimulatory molecules OX40L, CD70, and 4-1BBL on dendritic cells. Blood 2009, 113, 2451–2460. [Google Scholar] [CrossRef]
- Caccuri, F.; Bugatti, A.; Corbellini, S.; Roversi, S.; Zani, A.; Mazzuca, P.; Marsico, S.; Caruso, A.; Giagulli, C. The Synthetic Dipeptide Pidotimod Shows a Chemokine-Like Activity through CXC Chemokine Receptor 3 (CXCR3). Int. J. Mol. Sci. 2019, 20, 5287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 15, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmead, B.; Salzberg, S. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Van den Hoogen, B.G.; van Boheemen, S.; de Rijck, J.; van Nieuwkoop, S.; Smith, D.J.; Laksono, B.; Gultyaev, A.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Excessive production and extreme editing of human metapneumovirus defective interfering RNA is associated with type I IFN induction. J. Gen. Virol. 2014, 95, 1625–1633. [Google Scholar] [CrossRef] [Green Version]
- Debiaggi, M.; Canducci, F.; Sampaolo, M.; Marinozzi, M.C.; Parea, M.; Terulla, C.; Colombo, A.A.; Alessandrino, E.P.; Bragotti, L.Z.; Arghittu, M.; et al. Persistent symptomless human metapneumovirus infection in hematopoietic stem cell transplant recipients. J. Infect. Dis. 2006, 194, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Lambrecht, B.N.; Hammad, H. Taking our breath away: Dendritic cells in the pathogenesis of asthma. Nat. Rev. Immunol. 2003, 3, 994–1003. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H. Lung dendritic cells in respiratory viral infection and asthma: From protection to immunopathology. Annu. Rev. Immunol. 2012, 30, 243–270. [Google Scholar] [CrossRef]
- Soumelis, V.; Liu, Y.J. Human thymic stromal lymphopoietin: A novel epithelial cell-derived cytokine and a potential key player in the induction of allergic inflammation. Springer Semin. Immunopathol. 2004, 25, 325–333. [Google Scholar] [CrossRef]
- Sozzani, S.; Del Prete, A.; Bosisio, D. Dendritic cell recruitment and activation in autoimmunity. J. Autoimmun. 2017, 85, 126–140. [Google Scholar] [CrossRef]
- Kwon, J.M.; Shim, J.W.; Ki, D.S.; Jung, H.L.; Park, M.S.; Shim, J.Y. Prevalence of respiratory viral infection in children hospitalized for acute lower respiratory tract diseases, and association of rhinovirus and influenza virus with asthma exacerbations. Korean J. Pediatr. 2014, 57, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Rudd, P.A.; Thomas, B.J.; Zaid, A.; MacDonald, M.; Kan-O, K.; Rolph, M.S.; Soorneedi, A.R.; Bardin, P.G.; Mahalingam, S. Role of human metapneumovirus and respiratory syncytial virus in asthma exacerbations: Where are we now? Clin. Sci. 2017, 131, 1713–1721. [Google Scholar] [CrossRef] [PubMed]
- Inagaki-Katashiba, N.; Ito, T.; Inaba, M.; Azuma, Y.; Tanaka, A.; Phan, V.; Kibata, K.; Satake, A.; Nomura, S. Statins can suppress DC-mediated Th2 responses through the repression of OX40-ligand and CCL17 expression. Eur. J. Immunol. 2019, 49, 2051–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cespedes, P.F.; Gonzalez, P.A.; Kalergis, A.M. Human metapneumovirus keeps dendritic cells from priming antigen-specific naive T cells. Immunology 2013, 139, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Plata, A.; Casola, A.; Suarez, G.; Yu, X.; Spetch, L.; Peeples, M.E.; Garofalo, R.P. Differential response of dendritic cells to human metapneumovirus and respiratory syncytial virus. Am. J. Respir. Cell Mol. Biol. 2006, 34, 320–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.C.; Battini, L.; Tuyama, A.C.; Macip, S.; Melendi, G.A.; Horga, M.A.; Gusella, G.L. Characterization of human metapneumovirus infection of myeloid dendritic cells. Virology 2007, 357, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Le Nouen, C.; Munir, S.; Losq, S.; Winter, C.C.; McCarty, T.; Stephany, D.A.; Holmes, K.L.; Bukreyev, A.; Rabin, R.L.; Collins, P.L.; et al. Infection and maturation of monocyte-derived human dendritic cells by human respiratory syncytial virus, human metapneumovirus, and human parainfluenza virus type 3. Virology 2009, 385, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Liu, Y.J.; Arima, K. Cellular and molecular mechanisms of TSLP function in human allergic disorders-TSLP programs the “Th2 code” in dendritic cells. Allergol. Int. 2012, 61, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.V.; Crowe, J.E., Jr.; Enriquez, R.; Minton, P.; Peebles, R.S., Jr.; Hamilton, R.G.; Higgins, S.; Griffin, M.; Hartert, T.V. Human metapneumovirus infection plays an etiologic role in acute asthma exacerbations requiring hospitalization in adults. J. Infect. Dis. 2005, 192, 1149–1153. [Google Scholar] [CrossRef]
- Coverstone, A.M.; Wilson, B.; Burgdorf, D.; Schechtman, K.B.; Storch, G.A.; Holtzman, M.J.; Castro, M.; Bacharier, L.B.; Sumino, K. Recurrent wheezing in children following human metapneumovirus infection. J. Allergy Clin. Immunol. 2018, 142, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Arestides, R.S.; He, H.; Westlake, R.M.; Chen, A.I.; Sharpe, A.H.; Perkins, D.L.; Finn, P.W. Costimulatory molecule OX40L is critical for both Th1 and Th2 responses in allergic inflammation. Eur. J. Immunol. 2002, 32, 2874–2880. [Google Scholar] [CrossRef]
- Akiba, H.; Miyahira, Y.; Atsuta, M.; Takeda, K.; Nohara, C.; Futagawa, T.; Matsuda, H.; Aoki, T.; Yagita, H.; Okumura, K. Critical contribution of OX40 ligand to T helper cell type 2 differentiation in experimental leishmaniasis. J. Exp. Med. 2000, 191, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Kaur, D.; Brightling, C. OX40/OX40 ligand interactions in T-cell regulation and asthma. Chest 2012, 141, 494–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vignuzzi, M.; López, C.B. Defective viral genomes are key drivers of the virus-host interaction. Nat. Microbiol. 2019, 7, 1075–1087. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bugatti, A.; Marsico, S.; Fogli, M.; Roversi, S.; Messali, S.; Bosisio, D.; Giagulli, C.; Caruso, A.; Sozzani, S.; Fiorentini, S.; et al. Human Metapneumovirus Establishes Persistent Infection in Lung Microvascular Endothelial Cells and Primes a Th2-Skewed Immune Response. Microorganisms 2020, 8, 824. https://doi.org/10.3390/microorganisms8060824
Bugatti A, Marsico S, Fogli M, Roversi S, Messali S, Bosisio D, Giagulli C, Caruso A, Sozzani S, Fiorentini S, et al. Human Metapneumovirus Establishes Persistent Infection in Lung Microvascular Endothelial Cells and Primes a Th2-Skewed Immune Response. Microorganisms. 2020; 8(6):824. https://doi.org/10.3390/microorganisms8060824
Chicago/Turabian StyleBugatti, Antonella, Stefania Marsico, Manuela Fogli, Sara Roversi, Serena Messali, Daniela Bosisio, Cinzia Giagulli, Arnaldo Caruso, Silvano Sozzani, Simona Fiorentini, and et al. 2020. "Human Metapneumovirus Establishes Persistent Infection in Lung Microvascular Endothelial Cells and Primes a Th2-Skewed Immune Response" Microorganisms 8, no. 6: 824. https://doi.org/10.3390/microorganisms8060824