Soy Metabolism by Gut Microbiota from Patients with Precancerous Intestinal Lesions

 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
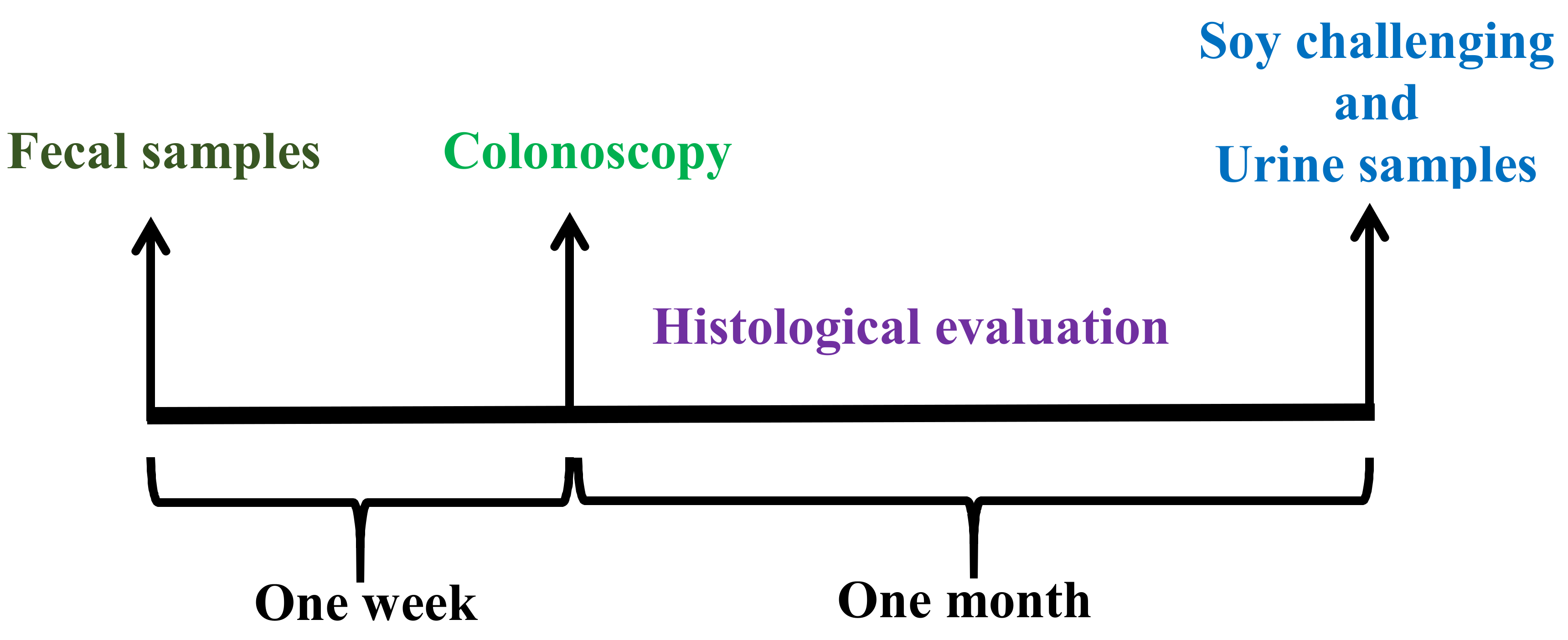
2.1. Patients Enrollment
2.2. Microbiological Study
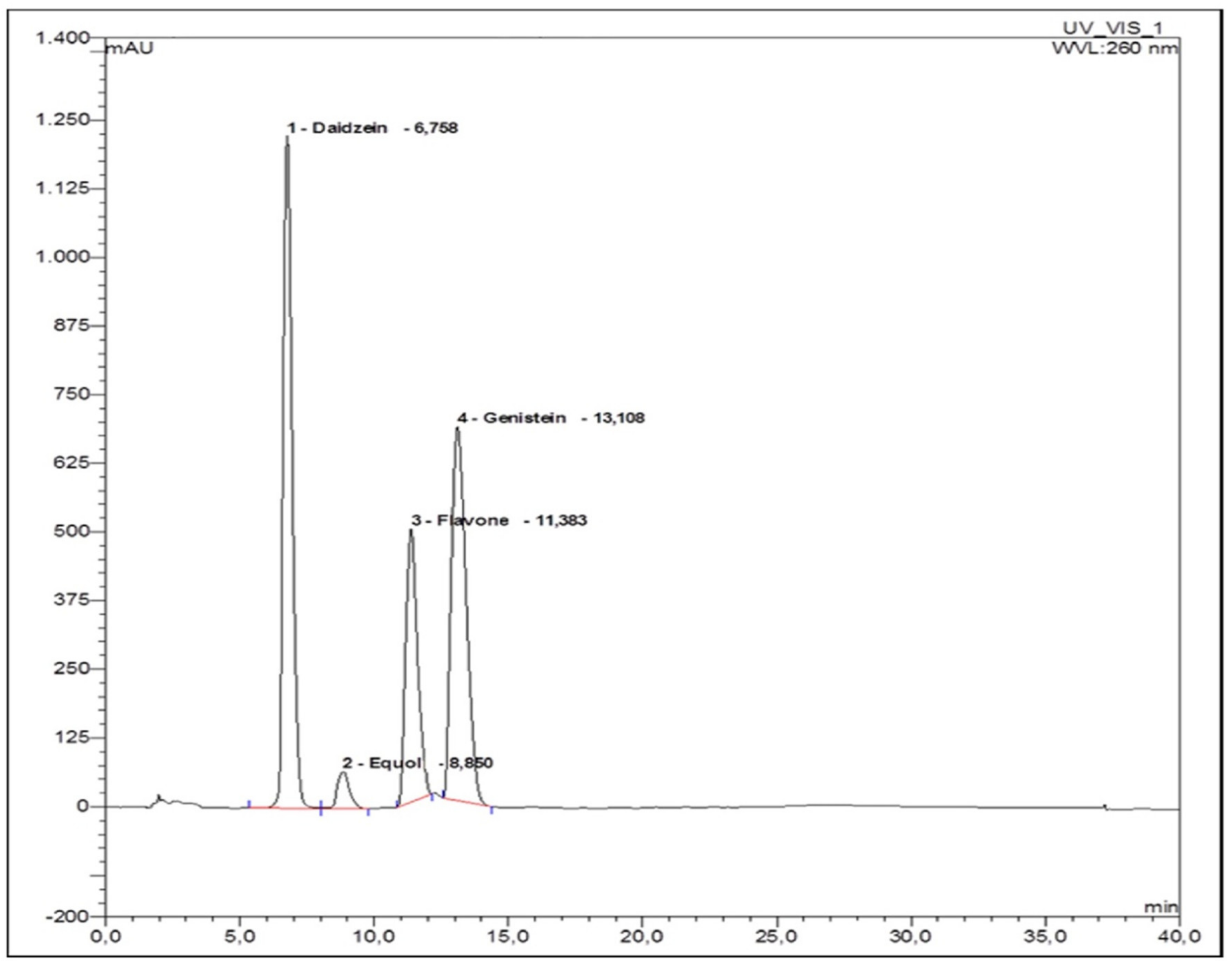
2.3. High Performance Liquid Chromatographic Assay of Isoflavonoids
2.3.1. Instrument
2.3.2. Chemicals
2.3.3. Standard Solutions and Calibration Curves
2.3.4. Chromatographic Study
2.3.5. Soy Challenging and Urine Collection
2.3.6. Extraction of Urinary Phytoestrogens
2.3.7. Enzyme Hydrolysis of Urine Phytoestrogens
2.3.8. Identification of Urinary Phytoestrogens
2.4. Statistical Analysis
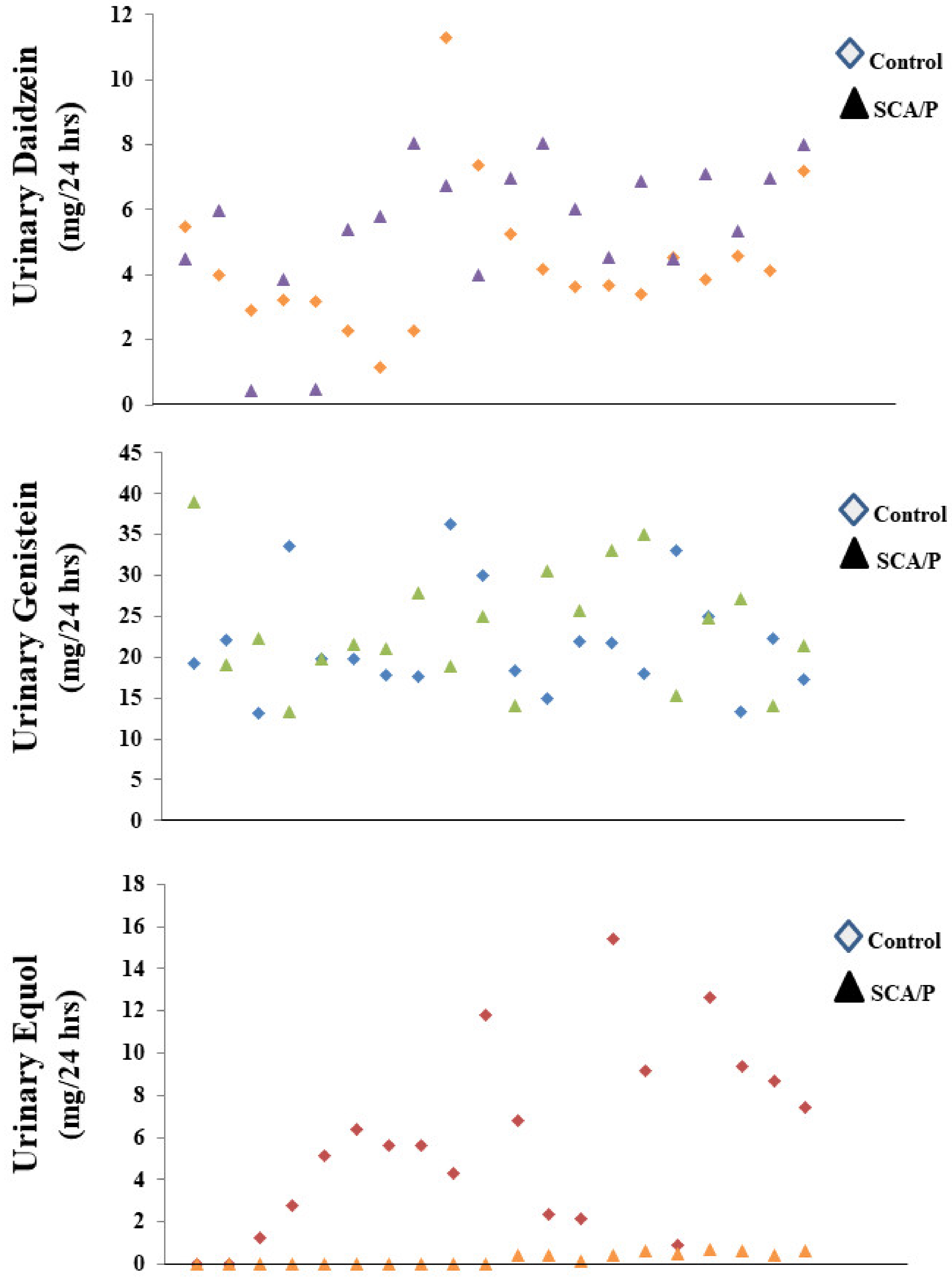
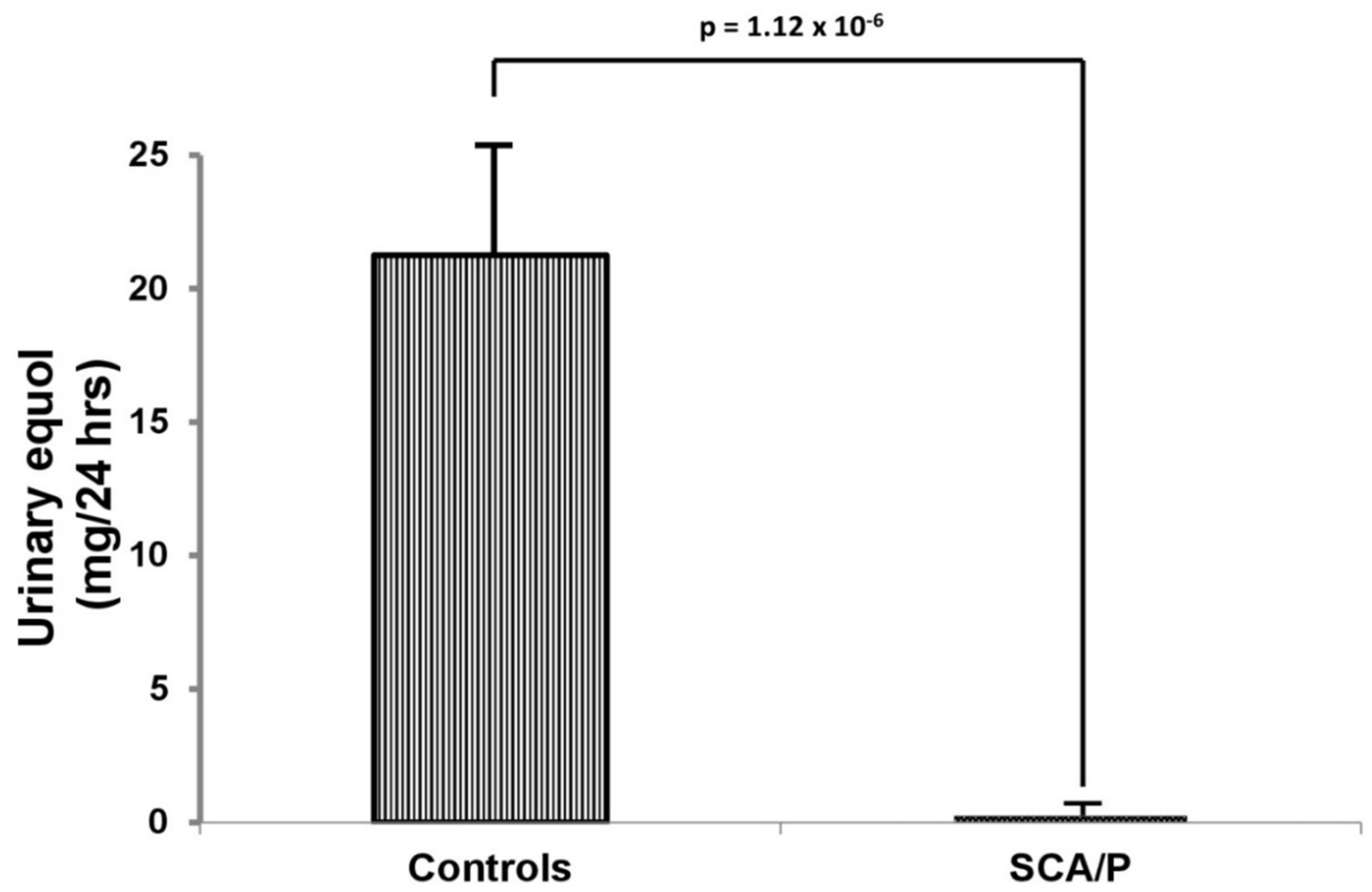
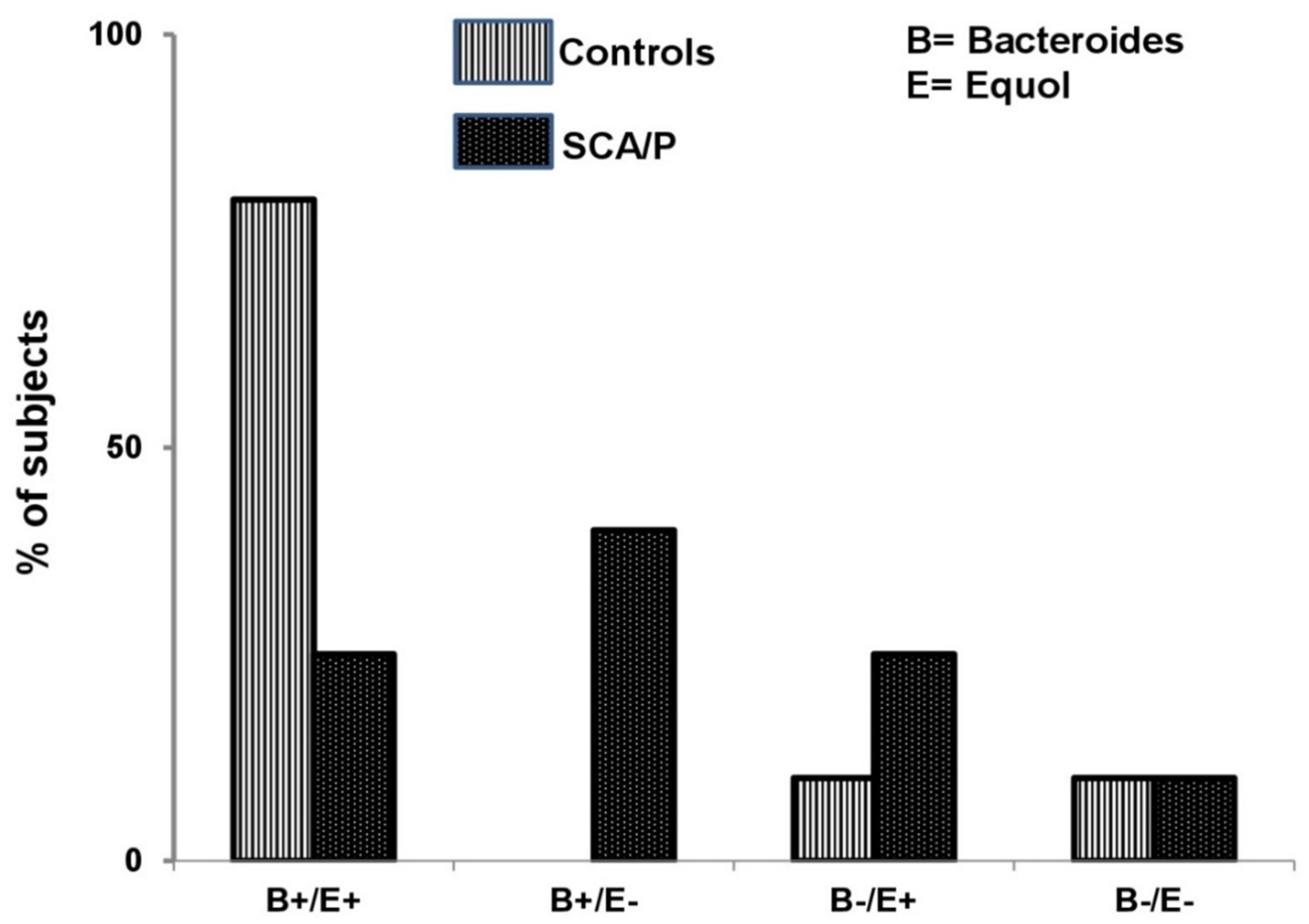
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wong, M.C.; Ding, H.; Wang, J.; Chan, P.S.; Huang, J. Prevalence and risk factors of colorectal cancer in Asia. Intest. Res. 2019, 17, 317–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barone, M.; Lofano, K.; De Tullio, N.; Licino, R.; Albano, F.; Di Leo, A. Dietary, Endocrine, and Metabolic Factors in the Development of Colorectal Cancer. J. Gastrointest. Cancer 2012, 43, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.; Tanzi, S.; Lofano, K.; Scavo, M.P.; Guido, R.; Demarinis, L.; Principi, M.B.; Bucci, A.; Di Leo, A. Estrogens, phytoestrogens and colorectal neoproliferative lesions. Genes Nutr. 2008, 3, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barone, M.; Scavo, M.P.; Papagni, S.; Piscitelli, D.; Guido, R.; Di Lena, M.; Comelli, M.C.; Di Leo, A. ERβ expression in normal, adenomatous and carcinomatous tissues of patients with familial adenomatous polyposis. Scand. J. Gastroenterol. 2010, 45, 1320–1328. [Google Scholar]
- Di Leo, A.; Barone, M.; Maiorano, E.; Tanzi, S.; Piscitelli, D.; Marangi, S.; Lofano, K.; Ierardi, E.; Principi, M.B.; Francavilla, A. ER-beta expression in large bowel adenomas: implications in colon carcinogenesis. Dig. Liver Dis. 2008, 40, 260–266. [Google Scholar] [CrossRef]
- Tlaskalova-Hogenova, H.; Vannucci, L.; Klimesova, K.; Stepankova, R.; Krizan, J.; Kverka, M. Microbiome and colorectal carcinoma: insights from germ-free and conventional animal models. Cancer J. 2014, 20, 217–224. [Google Scholar] [CrossRef]
- Akin, H.; Tözün, N. Diet, microbiota, and colorectal cancer. J. Clin. Gastroenterol. 2014, 48, S67–S69. [Google Scholar] [CrossRef]
- Garrett, W.S. Cancer and the microbiota. Science 2015, 348, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Hold, G.L. Western lifestyle: A ‘master’ manipulator of the intestinal microbiota? Gut 2014, 63, 5–6. [Google Scholar] [CrossRef]
- Nowak, A.; Libudzisz, Z. Influence of phenol, p-cresol and indole on growth and survival of intestinal lactic acid bacteria. Anaerobe 2006, 12, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Carbonero, F.; Zoetendal, E.G.; DeLany, J.P.; Wang, M.; Newton, K.; Gaskins, H.R.; O’Keefe, S.J. Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 2013, 98, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, W.R.; Duncan, S.H.; Scobbie, L.; Duncan, G.; Cantlay, L.; Calder, A.G.; Anderson, S.E.; Flint, H.J. Major phenylpropanoid-derived metabolites in the human gut can arise from microbial fermentation of protein. Mol. Nutr. Food Res. 2013, 57, 523–535. [Google Scholar] [CrossRef]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Gill, C.I.; Rowland, I.R. Diet and cancer: assessing the risk. Br. J. Nutr. 2002, 88, S73–S87. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Yu, Y.; Wang, M.; Li, Z.; Xu, H.; Tian, C.; Zhang, J.; Ye, X.; Li, X. Correlation between microbes and colorectal cancer: tumor apoptosis is induced by sitosterols through promoting gut microbiota to produce short-chain fatty acids. Apoptosis 2019, 24, 168–183. [Google Scholar] [CrossRef]
- Kandiah, M. Food intake and colorectal adenomas: a case-control study in Malaysia. Asian Pac. J. Cancer Prev. 2009, 10, 925–932. [Google Scholar]
- Sonoda, T.; Nagata, Y.; Mori, M.; Miyanaga, N.; Takaschima, N.; Okumura, K.; Goto, K.; Naito, S.; Fujimoto, K.; Hirao, Y.; et al. A case-control study of diet and prostate cancer in Japan: Possible prospective effect of traditional Japanese diet. Cancer Sci. 2004, 95, 238–242. [Google Scholar] [CrossRef]
- Khosravi, A.; Mazmanian, S.K. Disruption of the gut microbiome as a risk factor for microbial infections. Curr. Opin. Microbiol. 2013, 16, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Koh, G.Y.; Kane, A.; Lee, K.; Xu, Q.; Wu, X.; Roper, J.; Mason, J.B.; Crott, J.W. Parabacteroides distasonis attenuates toll-like receptor 4 signaling and Akt activation and blocks colon tumor formation in high-fat diet-fed azoxymethane-treated mice. Int. J. Cancer. 2018, 143, 1797–1805. [Google Scholar] [CrossRef] [Green Version]
- Bevins, C.L.; Salzman, N.H. Paneth cells, antimicrobial peptides and maintenance of intestinal homeostasis. Nat. Rev. Microbiol. 2011, 9, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Tuddenham, S.; Sears, C.L. The Intestinal Microbiome and Health. Curr. Opin. Infect. Dis. 2015, 28, 464–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, C.V.; de Camargo, M.R.; Russo, E.; Amedei, A. Role of diet and gut microbiota on colorectal cancer immunomodulation. World J. Gastroenterol. 2019, 25, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Lechner, D.; Kállay, E.; Cross, H.S. Phytoestrogens and colorectal cancer prevention. Vitam Horm. 2005, 70, 169–198. [Google Scholar]
- Bennink, M.R. Dietary soy reduces colon carcinogenesis in human and rats. Soy and colon cancer. Adv. Exp. Med. Biol. 2001, 492, 11–17. [Google Scholar]
- Murphy, N.; Achaintre, D.; Zamora-Ros, R.; Jenab, M.; Boutron-Ruault, M.C.; Carbonnel, F.; Savoye, I.; Kaaks, R.; Kuhn, T.; Boeing, H.; et al. A prospective evaluation of plasma polyphenol levels and colon cancer risk. Int. J. Cancer 2018, 143, 1620–1631. [Google Scholar] [CrossRef]
- Sarkar, F.H.; Li, Y. Soy isoflavones and cancer prevention. Cancer Invest. 2003, 21, 744–757. [Google Scholar] [CrossRef]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A Bacterial Metabolite from The Daidzein Isoflavone and Its Presumed Beneficial Health Effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef] [Green Version]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [Green Version]
- Polimeno, L.; Barone, M.; Mosca, A.; Viggiani, M.T.; Di Leo, A.; Debellis, L.; Troisi, M.; Daniele, A.; Santacroce, L. Gut Microbiota Imbalance is Related to Sporadic Colorectal Neoplasms. A Pilot Study. Appl. Sci. 2019, 9, 5491. [Google Scholar] [CrossRef] [Green Version]
- Schubert, S.; Kostrzewa, M. MALDI-TOF MS in the Microbiology Laboratory: Current Trends. Curr. Issues Mol. Biol. 2017, 23, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angeletti, S. Matrix assisted laser desorption time of flight mass spectrometry (MALDI-TOF MS) in clinical microbiology. J. Microbiol. Methods 2017, 138, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Franke, A.A.; Custer, L.J. High-performance liquid chromatographic assay of isoflavonoids and coumestrol from human urine. J. Chromatogr. B. Biomed. Appl. 1994, 662, 47–60. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, W.; Huang, S.; Yao, W. Microbial conversion of daidzein affects fecal equol concentration and bacterial composition of rats with or without ovariectomy. Wei Sheng Wu Xue Bao 2012, 52, 866–874. [Google Scholar]
- Agrawal, B. New therapeutic targets for cancer: the interplay between immune and metabolic checkpoints and gut microbiota. Clin. Transl. Med. 2019, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.F.; Zhang, H.M.; Niu, W.Y.; Zou, Y.Q.; Ma, D.F. Effects of equol on colon cancer cell proliferation. Beijing Da Xue Xue Bao Yi Xue Ban 2017, 49, 383–387. [Google Scholar] [PubMed]
- Barone, M.; Tanzi, S.; Lofano, K.; Scavo, M.P.; Pricci, M.; Demarinis, L.; Papagni, S.; Guido, R.; Maiorano, E.; Ingravallo, G.; et al. Dietary-induced ERbeta upregulation counteracts intestinal neoplasia development in intact male ApcMin/+ mice. Carcinogenesis 2010, 31, 269–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef] [Green Version]
- Costello, E.K.; Lauber, C.L.; Hamady, M.; Fierer, N.; Gordon, J.I.; Knight, R. Bacterial community variation in human body habitats across space and time. Science 2009, 326, 1694–1697. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-gut microbiota metabolic interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamida, K.; Tanaka, M.; Abe, A.; Sone, T.; Tomita, F.; Hara, H.; Asano, K. Production of equol from daidzein by gram-positive rod-shaped bacterium isolated from rat intestine. J. Biosci. Bioeng. 2006, 102, 247–250. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.L.; Hur, H.G.; Lee, J.H.; Kim, K.T.; Kim, S.I. Enantioselective synthesis of S-equol from dihydrodaidzein by a newly isolated anaerobic human intestinal bacterium. Appl. Environ. Microbiol. 2005, 71, 214–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.C.; Hevia, D.; Patchva, S.; Park, B.; Koh, W.; Aggarwal, B.B. Upsides and downsides of reactive oxygen species for cancer: the roles of reactive oxygen species in tumorigenesis, prevention, and therapy. Antioxid. Redox Signal. 2012, 16, 1295–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwong, T.N.Y.; Wang, X.; Nakatsu, G.; Chow, T.C.; Tipoe, T.; Dai, R.Z.W.; Tsoi, K.K.K.; Wong, M.C.S.; Tse, G.; Chan, M.T.V.; et al. Association Between Bacteremia From Specific Microbes and Subsequent Diagnosis of Colorectal Cancer. Gastroenterology 2018, 155, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Shiotani, A.; Fukushima, S.; Matsumoto, H. Carcinogenesis and Gut Microbiota. Gan. To Kagaku Ryoho. 2019, 46, 199–204. [Google Scholar] [PubMed]
- Topi, S.; Santacroce, L.; Bottalico, L.; Ballini, A.; Inchingolo, A.D.; Dipalma, G.; Charitos, I.A.; Inchingolo, F. Gastric Cancer in History: A Perspective Interdisciplinary Study. Cancers 2020, 12, 264. [Google Scholar] [CrossRef] [Green Version]
- Santacroce, L.; Cagiano, R.; Del Prete, R.; Bottalico, L.; Sabatini, R.; Carlaio, R.G.; Prejbeanu, R.; Vermesan, H.; Dragulescu, S.I.; Vermesan, D.; et al. Helicobacter pylori infection and gastric MALTomas: An up-to-date and therapy highlight. Clin. Ter. 2008, 159, 457–462. [Google Scholar]
- Ballini, A.; Cantore, S.; Farronato, D.; Cirulli, N.; Inchingolo, F.; Papa, F.; Malcangi, G.; Inchingolo, A.D.; Dipalma, G.; Sardaro, N.; et al. Periodontal disease and bone pathogenesis: the crosstalk between cytokines and porphyromonas gingivalis. J. Biol. Regul. Homeost. Agents. 2015, 29, 273–281. [Google Scholar]
- Di Serio, F.; Lovero, R.; D’Agostino, D.; Nisi, L.; Miragliotta, G.; Contino, R.; Man, A.; Ciccone, M.M.; Santacroce, L. Evaluation of procalcitonin, Vitamin D and C-reactive protein levels in septic patients with positive emocoltures. Our preliminary experience. Acta Med. Mediterr. 2016, 32, 1911–1914. [Google Scholar]
- Giudice, G.; Cutrignelli, D.A.; Sportelli, P.; Limongelli, L.; Tempesta, A.; Gioia, G.D.; Santacroce, L.; Maiorano, E.; Favia, G. Rhinocerebral Mucormycosis with Orosinusal Involvement: Diagnostic and Surgical Treatment Guidelines. Endocr. Metab. Immune. Disord. Drug. Targets. 2016, 16, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Inchingolo, F.; Dipalma, G.; Cirulli, N.; Cantore, S.; Saini, R.S.; Altini, V.; Santacroce, L.; Ballini, A.; Saini, R. Microbiological results of improvement in periodontal condition by administration of oral probiotics. J. Biol. Regul. Homeost. Agents. 2018, 32, 1323–1328. [Google Scholar] [PubMed]
- Man, A.; Santacroce, L.; Jacob, R.; Mare, A.; Man, L. Antimicrobial Activity of Six Essential Oils Against a Group of Human Pathogens: A Comparative Study. Pathogens 2019, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santacroce, L.; Charitos, I.A.; Bottalico, L. A successful history: probiotics and their potential as antimicrobials. Expert. Rev. Anti. Infect. Ther. 2019, 17, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Santacroce, L.; Cantore, S.; Bottalico, L.; Dipalma, G.; Topi, S.; Saini, R.; De Vito, D.; Inchingolo, F. Probiotics Efficacy on Oxidative Stress Values in Inflammatory Bowel Disease: A Randomized Double-Blinded Placebo-Controlled Pilot Study. Endocr. Metab. Immune. Disord. Drug. Targets. 2019, 19, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Gnoni, A.; De Vito, D.; Dipalma, G.; Cantore, S.; Gargiulo Isacco, C.; Saini, R.; Santacroce, L.; Topi, S.; Scarano, A.; et al. Effect of probiotics on the occurrence of nutrition absorption capacities in healthy children: a randomized double-blinded placebo-controlled pilot study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8645–8657. [Google Scholar] [PubMed]
- Shamekhi, S.; Lotfi, H.; Abdolalizadeh, J.; Bonabi, E.; Zarghami, N. An overview of yeast probiotics as cancer biotherapeutics: possible clinical application in colorectal cancer. Clin. Transl. Oncol. 2020. [Google Scholar] [CrossRef]
- Molska, M.; Reguła, J. Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer. Nutrients 2019, 11, 2453. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SCA/P | Controls | p-Value | |
|---|---|---|---|
| Age (yrs) | 63.4 ± 8 | 64.5 ± 4 | 0.56 |
| Sex ratio (M/F) | 14:6 | 11:9 | 0.61 |
| BMI | 29.8 ± 5.4 | 30.3 ± 7.2 | 0.76 |
| Weight (Kg) | 80.5 ± 12.3 | 77.3 ± 13.8 | 0.45 |
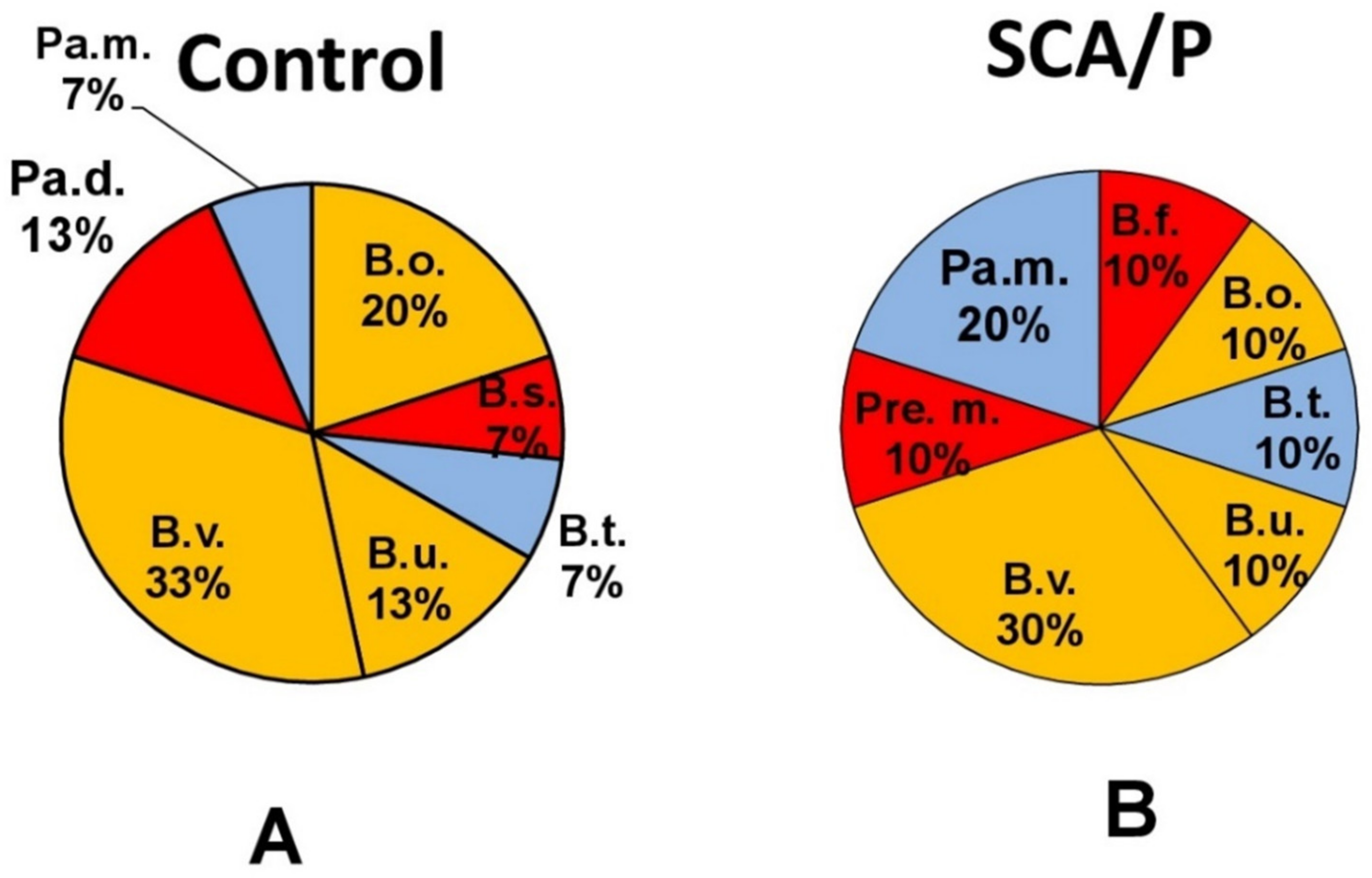
| Gram - bacteria | Gram + bacteria |
|---|---|
| 1. Bacteroides fragilis (B.f.) | 1. Bifidobacterium spp (Biff.) |
| 2. Bacteroides ovatus (B.o.) | 2. Clostridium clostridioforme (C.c.) |
| 3. Bacteroides stercoris (B.s.) | 3. Clostridium innocuum (C.i.) |
| 4. Bacteroides thetaiotaomicron (B.t.) | 4. Clostridium ramosum (C.r.) |
| 5. Bacteroides uniformis (B.u.) | 5. Clostridium sardelli (C.s.) |
| 6. Bacteroides vulgatus (B.v.) | 6. Corynebacterium xerosus (Cor.x.) |
| 7. Prevotella melaninigenica (Pre.m.) | 7. Propionibacterium avidum (Pr.a.) |
| 8. Parabacteroides distasonis (Pa.d.) | |
| 9. Parabacteroides merdae (Pa.m.) | |
| 10. Pediococcus pentasaceus (Pe.p.) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polimeno, L.; Barone, M.; Mosca, A.; Viggiani, M.T.; Joukar, F.; Mansour-Ghanaei, F.; Mavaddati, S.; Daniele, A.; Debellis, L.; Bilancia, M.; et al. Soy Metabolism by Gut Microbiota from Patients with Precancerous Intestinal Lesions. Microorganisms 2020, 8, 469. https://doi.org/10.3390/microorganisms8040469
Polimeno L, Barone M, Mosca A, Viggiani MT, Joukar F, Mansour-Ghanaei F, Mavaddati S, Daniele A, Debellis L, Bilancia M, et al. Soy Metabolism by Gut Microbiota from Patients with Precancerous Intestinal Lesions. Microorganisms. 2020; 8(4):469. https://doi.org/10.3390/microorganisms8040469
Chicago/Turabian StylePolimeno, Lorenzo, Michele Barone, Adriana Mosca, Maria Teresa Viggiani, Farahnaz Joukar, Fariborz Mansour-Ghanaei, Sara Mavaddati, Antonella Daniele, Lucantonio Debellis, Massimo Bilancia, and et al. 2020. "Soy Metabolism by Gut Microbiota from Patients with Precancerous Intestinal Lesions" Microorganisms 8, no. 4: 469. https://doi.org/10.3390/microorganisms8040469