Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps

 , , ,
, , , 
Abstract
:1. Introduction
2. Bacterial Pathogens
3. Bacterial Resistance to Antimicrobials
Mechanisms of Bacterial Resistance to Antimicrobial Agents
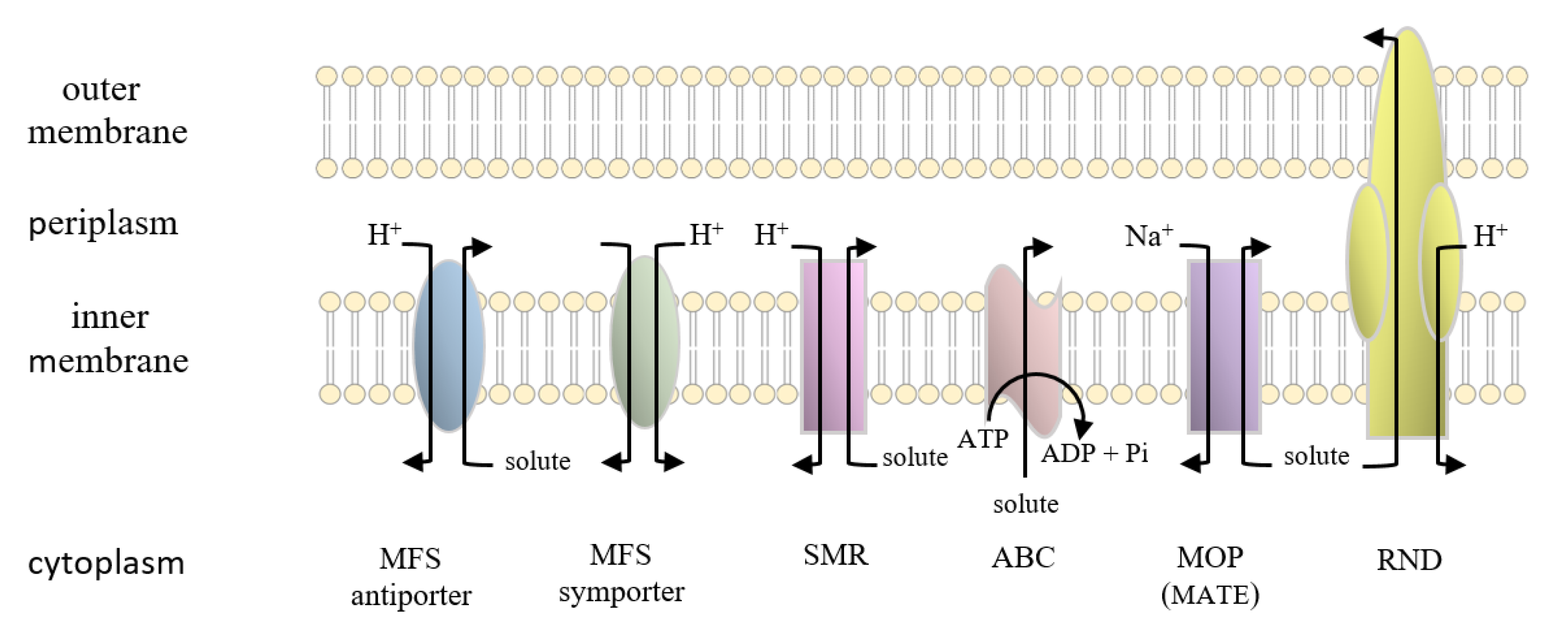
4. Transporter Superfamilies
5. The Major Facilitator Superfamily
6. Structure-Function Studies of Bacterial Multidrug Efflux Pumps from the Major Facilitator Superfamily
7. Modulation of Multidrug Efflux Pumps of the Major Facilitator Superfamily
8. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Peterson, J.W. Bacterial pathogenesis. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch: Galveston, TX, USA, 1996. [Google Scholar]
- Andersen, J.L.; He, G.X.; Kakarla, P.; KC, R.; Kumar, S.; Lakra, W.S.; Mukherjee, M.M.; Ranaweera, I.; Shrestha, U.; Tran, T.; et al. Multidrug efflux pumps from Enterobacteriaceae, Vibrio cholerae and Staphylococcus aureus bacterial food pathogens. Int. J. Environ. Res. Public Health 2015, 12, 1487–1547. [Google Scholar] [CrossRef] [Green Version]
- Lekshmi, M.; Ammini, P.; Kumar, S.; Varela, M.F. The food production environment and the development of antimicrobial resistance in human pathogens of animal origin. Microorganisms 2017, 5, 11. [Google Scholar] [CrossRef]
- Friedman, N.D.; Temkin, E.; Carmeli, Y. The negative impact of antibiotic resistance. Clin. Microbiol. Infect. 2016, 22, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Varela, M.F. Molecular mechanisms of bacterial resistance to antimicrobial agents. In Microbial Pathogens and Strategies for Combating Them: Science, Technology and Education; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2013; pp. 522–534. [Google Scholar]
- Floyd, J.T.; Kumar, S.; Mukherjee, M.M.; He, G.; Varela, M.F. A review of the molecular mechanisms of drug efflux in pathogenic bacteria: A structure-function perspective. Recent Res. Dev. Membr. Biol. 2013, 3, 15–66. [Google Scholar]
- Varela, M.F. Antimicrobial efflux pumps. In Antibiotic Drug Resistance; Capelo-Martinez, J.L., Igrejas, G., Eds.; Wiley: Hoboken, NJ, USA, 2019; pp. 167–179. [Google Scholar]
- Saier, M.H., Jr. A functional-phylogenetic system for the classification of transport proteins. J. Cell. Biochem. 1999, 32, 84–94. [Google Scholar] [CrossRef]
- Griffith, J.K.; Baker, M.E.; Rouch, D.A.; Page, M.G.; Skurray, R.A.; Paulsen, I.T.; Chater, K.F.; Baldwin, S.A.; Henderson, P.J. Membrane transport proteins: Implications of sequence comparisons. Curr. Opin. Cell. Biol. 1992, 4, 684–695. [Google Scholar] [CrossRef]
- Kakarla, P.; Ranjana, K.; Shrestha, U.; Ranaweera, I.; Mukherjee, M.M.; Willmon, T.M.; Hernandez, A.J.; Barr, S.R.; Varela, M.F. Functional roles of highly conserved amino acid sequence motifs A and C in solute transporters of the major facilitator superfamily. In Drug Resistance in Bacteria, Fungi, Malaria, and Cancer; Springer: Berlin/Heidelberg, Germany, 2017; pp. 111–140. [Google Scholar]
- Pao, S.S.; Paulsen, I.T.; Saier, M.H., Jr. Major facilitator superfamily. Microbiol. Mol. Biol. Rev. 1998, 62, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Law, C.J.; Maloney, P.C.; Wang, D.N. Ins and outs of major facilitator superfamily antiporters. Annu. Rev. Microbiol. 2008, 62, 289–305. [Google Scholar] [CrossRef] [Green Version]
- Reddy, V.S.; Shlykov, M.A.; Castillo, R.; Sun, E.I.; Saier, M.H., Jr. The major facilitator superfamily (MFS) revisited. FEBS J. 2012, 279, 2022–2035. [Google Scholar] [CrossRef]
- Pasqua, M.; Grossi, M.; Zennaro, A.; Fanelli, G.; Micheli, G.; Barras, F.; Colonna, B.; Prosseda, G. The varied role of efflux pumps of the MFS family in the interplay of bacteria with animal and plant cells. Microorganisms 2019, 7, 285. [Google Scholar] [CrossRef] [Green Version]
- Djeussi, D.E.; Noumedem, J.A.; Seukep, J.A.; Fankam, A.G.; Voukeng, I.K.; Tankeo, S.B.; Nkuete, A.H.; Kuete, V. Antibacterial activities of selected edible plants extracts against multidrug-resistant Gram-negative bacteria. BMC Complement. Altern. Med. 2013, 13, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, K.K.; Bertozzi, S.; Bloom, B.R.; Jha, P.; Gelband, H.; DeMaria, L.M.; Horton, S. Major infectious diseases: Key messages from disease control priorities. In Major Infectious Diseases, 3rd ed.; World Bank Publications: Washington, DC, USA, 2017. [Google Scholar]
- Conly, J. Antimicrobial resistance: Revisiting the tragedy of the commons. Bull World Health Organ 2010, 88, 805–806. [Google Scholar]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef] [PubMed]
- Stanley, I.J.; Kajumbula, H.; Bazira, J.; Kansiime, C.; Rwego, I.B.; Asiimwe, B.B. Multidrug resistance among Escherichia coli and Klebsiella pneumoniae carried in the gut of out-patients from pastoralist communities of Kasese district, Uganda. PLoS ONE 2018, 13, e0200093. [Google Scholar] [CrossRef] [PubMed]
- Chong, Y.; Shimoda, S.; Shimono, N. Current epidemiology, genetic evolution and clinical impact of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect. Genet. Evol. 2018, 61, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Gygli, S.M.; Borrell, S.; Trauner, A.; Gagneux, S. Antimicrobial resistance in Mycobacterium tuberculosis: Mechanistic and evolutionary perspectives. FEMS Microbiol. Rev. 2017, 41, 354–373. [Google Scholar] [CrossRef] [Green Version]
- Zignol, M.; Hosseini, M.S.; Wright, A.; Weezenbeek, C.L.; Nunn, P.; Watt, C.J.; Williams, B.G.; Dye, C. Global incidence of multidrug-resistant tuberculosis. J. Infect. Dis. 2006, 194, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.S.; Viveiros, M.; Amaral, L.; Couto, I. Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 2013, 7, 59. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.N.; Feazel, L.M.; Bessesen, M.T.; Price, C.S.; Janoff, E.N.; Pace, N.R. The human nasal microbiota and Staphylococcus aureus carriage. PLoS ONE 2010, 5, e10598. [Google Scholar] [CrossRef]
- Shrestha, U.; Ranaweera, I.; Kumar, S.; KC, R.; Kakarla, P.; Lakra, W.S.; He, G.; Andersen, J.; Varela, M.F. Multidrug resistance efflux pumps of Salmonella enterica. In Salmonella: Prevalence, Risk Factors and Treatment Options; Nova Science Publishers, Inc.: New York, NY, USA, 2015; pp. 1–30. [Google Scholar]
- Smith, K.P.; George, J.; Cadle, K.M.; Kumar, S.; Aragon, S.J.; Hernandez, R.L.; Jones, S.E.; Floyd, J.L.; Varela, M.F. Elucidation of antimicrobial susceptibility profiles and genotyping of Salmonella enterica isolates from clinical cases of salmonellosis in New Mexico in 2008. World J. Microbiol. Biotechnol. 2010, 26, 1025–1031. [Google Scholar] [CrossRef]
- Riedel, S.; Vijayakumar, D.; Berg, G.; Kang, A.D.; Smith, K.P.; Kirby, J.E. Evaluation of apramycin against spectinomycin-resistant and -susceptible strains of Neisseria gonorrhoeae. J. Antimicrob. Chemother. 2019, 74, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Klugman KP Madhi, S.A. Emergence of drug resistance: Impact on bacterial meningitis. Infect. Dis. Clin. N. Am. 1999, 13, 637–646. [Google Scholar] [CrossRef]
- Di Pietro, M.; Filardo, S.; De Santis, F.; Mastromarino, P.; Sessa, R. Chlamydia pneumoniae and oxidative stress in cardiovascular disease: State of the art and prevention strategies. Int. J. Mol. Sci. 2014, 16, 724–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neu, H.C. The crisis in antibiotic resistance. Science 1992, 257, 1064–1073. [Google Scholar] [CrossRef] [Green Version]
- Blair, J.M.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10 (Suppl. 12), S122–S129. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug resistance in bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Zaman, S.B.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A review on antibiotic resistance: Alarm bells are ringing. Cureus 2017, 9, e1403. [Google Scholar] [CrossRef] [Green Version]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Varela MF Kumar, S. Strategies for discovery of new molecular targets for anti-infective drugs. Curr. Opin. Pharmacol. 2019, 48, 57–68. [Google Scholar] [CrossRef]
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; He, G.; Kakarla, P.; Shrestha, U.; Ranjana, K.C.; Ranaweera, I.; Willmon, T.M.; Barr, S.R.; Hernandez, A.J.; Varela, M.F. Bacterial multidrug efflux pumps of the major facilitator superfamily as targets for modulation. Infect. Disord. Drug Targets 2016, 16, 28–43. [Google Scholar] [CrossRef] [PubMed]
- McDermott, P.F.; Walker, R.D.; White, D.G. Antimicrobials: Modes of action and mechanisms of resistance. Int. J. Toxicol. 2003, 22, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Spratt, B.G. Resistance to antibiotics mediated by target alterations. Science 1994, 264, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Davies, J.; Wright, G.D. Bacterial resistance to aminoglycoside antibiotics. Trends Microbiol. 1997, 5, 234–240. [Google Scholar] [CrossRef]
- Wright, G.D. Aminoglycoside-modifying enzymes. Curr. Opin. Microbiol. 1999, 2, 499–503. [Google Scholar] [CrossRef]
- Vu, H.; Nikaido, H. Role of β-lactam hydrolysis in the mechanism of resistance of a β-lactamase-constitutive Enterobacter cloacae strain to expanded-spectrum β-lactams. Antimicrob. Agents Chemother. 1985, 27, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Levy, S.B. Active efflux, a common mechanism for biocide and antibiotic resistance. Symp. Ser. Soc. Appl. Microbiol. 2002, 31, 65S–71S. [Google Scholar] [CrossRef]
- Nikaido, H. Prevention of drug access to bacterial targets: Permeability barriers and active efflux. Science 1994, 264, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Henderson, P.J. Proton-linked sugar transport systems in bacteria. J. Bioenerg. Biomembr. 1990, 22, 525–569. [Google Scholar] [CrossRef]
- Nelson, M.L.; Levy, S.B. The history of the tetracyclines. Ann. N. Y. Acad. Sci. 2011, 1241, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Varela, M.F.; Andersen, J.L.; Ranjana, K.; Kumar, S.; Sanford, L.M.; Hernandez, A.J. Bacterial resistance mechanisms and inhibitors of multidrug efflux pumps belonging to the major facilitator superfamily of solute transport systems. In Frontiers in Anti-Infective Drug Discovery; Bentham Science Publishers: Sharjah, UAE, 2017; pp. 109–131. [Google Scholar]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Principles of membrane transport. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Saier, M.H., Jr. A functional-phylogenetic classification system for transmembrane solute transporters. Microbiol. Mol. Biol. Rev. 2000, 64, 354–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, I.C. Ligand conduction and the gated-pore mechanism of transmembrane transport. Biochim. Biophys. Acta 1997, 1331, 213–234. [Google Scholar] [CrossRef]
- Nakae, T. Outer membrane of Salmonella. Isolation of protein complex that produces transmembrane channels. J. Biol. Chem. 1976, 251, 2176–2178. [Google Scholar] [PubMed]
- Wiener, M.C.; Horanyi, P.S. How hydrophobic molecules traverse the outer membranes of gram-negative bacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 10929–10930. [Google Scholar] [CrossRef] [Green Version]
- Todt, J.C.; Rocque, W.J.; Groarty, E.J. Effects of pH on bacterial porin function. Biochemistry 1992, 31, 10471–10478. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Carrier proteins and active membrane transport. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Dills, S.S.; Apperson, A.; Schmidt, M.R.; Saier, M.H., Jr. Carbohydrate transport in bacteria. Microbiol. Rev. 1980, 44, 385–418. [Google Scholar] [CrossRef]
- Varela, M.F.; Wilson, T.H. Molecular biology of the lactose carrier of Escherichia coli. Biochim. Biophys. Acta 1996, 1276, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism. Nature 1961, 191, 144–148. [Google Scholar] [CrossRef]
- West, I.C.; Mitchell, P. Stoicheiometry of lactose-H+ symport across the plasma membrane of Escherichia coli. Biochem. J. 1973, 132, 587–592. [Google Scholar] [CrossRef] [PubMed]
- West, I.C. Energy coupling in secondary active transport. Biochim. Biophys. Acta. 1980, 604, 91–126. [Google Scholar] [CrossRef]
- Wilson, T.H.; Ding, P.Z. Sodium-substrate cotransport in bacteria. Biochim. Biophys. Acta. 2001, 1505, 121–130. [Google Scholar] [CrossRef] [Green Version]
- Botfield, M.C.; Wilson, D.M.; Wilson, T.H. The melibiose carrier of Escherichia coli. Res. Microbiol. 1990, 141, 328–331. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Harada, S.; Tokuyama, S. Sodium-glucose transporter as a novel therapeutic target in disease. Eur. J. Pharmacol. 2018, 822, 25–31. [Google Scholar] [CrossRef]
- Deng, D.; Yan, N. GLUT, SGLT, and SWEET: Structural and mechanistic investigations of the glucose transporters. Protein. Sci. 2016, 25, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, S.B.; Fenton, R.A.; Rieg, T. Sodium-glucose cotransport. Curr. Opin. Nephrol. Hypertens. 2015, 24, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.M.; Loo, D.D.; Hirayama, B.A. Biology of human sodium glucose transporters. Physiol. Rev. 2011, 91, 733–794. [Google Scholar] [CrossRef] [Green Version]
- Lodish, H.; Berk, A.; Zipursky, S.L.; Matsudaira, P.; Baltimore, D.; Darnell, J. Cotransport by symporters and antiporters. In Molecular Cell Biology, 4th ed.; W.H. Freeman & Company: New York, NY, USA, 2000. [Google Scholar]
- Kumar, A.; Schweizer, H.P. Bacterial resistance to antibiotics: Active efflux and reduced uptake. Adv. Drug Deliv. Rev. 2005, 57, 1486–1513. [Google Scholar] [CrossRef]
- Kuroda, T.; Tsuchiya, T. Multidrug efflux transporters in the MATE family. Biochim. Biophys. Acta. 2009, 1794, 763–768. [Google Scholar] [CrossRef]
- Busch, W.; Saier, M.H., Jr. The IUBMB-endorsed transporter classification system. Mol. Biotechnol. 2004, 27, 253–262. [Google Scholar] [CrossRef]
- Chang, A.B.; Lin, R.; Studley, W.K.; Tran, C.V.; Saier, M.H., Jr. Phylogeny as a guide to structure and function of membrane transport proteins. Mol. Membr. Biol. 2004, 21, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Saier, M.H., Jr.; Tran, C.V.; Barabote, R.D. TCDB: The Transporter Classification Database for membrane transport protein analyses and information. Nucleic Acids Res. 2006, 34, D181–D186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saier, M.H., Jr.; Reddy, V.S.; Tsu, B.V.; Ahmed, M.S.; Li, C.; Moreno-Hagelsieb, G. The transporter classification database (TCDB): Recent advances. Nucleic Acids Res. 2016, 44, D372–D379. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H. RND transporters in the living world. Res. Microbiol. 2018, 169, 363–371. [Google Scholar] [CrossRef]
- Hvorup, R.N.; Winnen, B.; Chang, A.B.; Jiang, Y.; Zhou, X.F.; Saier, M.H., Jr. The multidrug/oligosaccharidyl-lipid/polysaccharide (MOP) exporter superfamily. Eur. J. Biochem. 2003, 270, 799–813. [Google Scholar] [CrossRef]
- He, G.X.; Thorpe, C.; Walsh, D.; Crow, R.; Chen, H.; Kumar, S.; Varela, M.F. EmmdR, a new member of the MATE family of multidrug transporters, extrudes quinolones from Enterobacter cloacae. Arch. Microbiol. 2011, 193, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Jack, D.L.; Yang, N.; Saier, M.H., Jr. The drug/metabolite transporter superfamily. Eur. J. Biochem. 2001, 268, 3620–3639. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Skurray, R.A.; Tam, R.; Saier, M.H., Jr.; Turner, R.J.; Weiner, J.H.; Goldberg, E.B.; Grinius, L.L. The SMR family: A novel family of multidrug efflux proteins involved with the efflux of lipophilic drugs. Mol. Microbiol. 1996, 19, 1167–1175. [Google Scholar] [CrossRef]
- He, G.X.; Zhang, C.; Crow, R.R.; Thorpe, C.; Chen, H.; Kumar, S.; Tsuchiya, T.; Varela, M.F. SugE, a new member of the SMR family of transporters, contributes to antimicrobial resistance in Enterobacter cloacae. Antimicrob. Agents Chemother. 2011, 55, 3954–3957. [Google Scholar] [CrossRef] [Green Version]
- Henderson, P.J. Sugar transport proteins. Curr. Opin. Struct. Biol. 1991, 1, 590–601. [Google Scholar] [CrossRef]
- Henderson, P.J. The 12-transmembrane helix transporters. Curr. Opin. Cell. Biol. 1993, 5, 708–721. [Google Scholar] [CrossRef]
- Saier, M.H., Jr.; Beatty, J.T.; Goffeau, A.; Harley, K.T.; Heijne, W.H.; Huang, S.C.; Jack, D.L.; Jahn, P.S.; Lew, K.; Liu, J.; et al. The major facilitator superfamily. J. Mol. Microbiol. Biotechnol. 1999, 1, 257–279. [Google Scholar] [PubMed]
- Maiden, M.C.; Davis, E.O.; Baldwin, S.A.; Moore, D.C.; Henderson, P.J. Mammalian and bacterial sugar transport proteins are homologous. Nature 1987, 325, 641–643. [Google Scholar] [CrossRef] [PubMed]
- Henderson, P.J.; Roberts, P.E.; Martin, G.E.; Seamon, K.B.; Walmsley, A.R.; Rutherford, N.G.; Varela, M.F.; Griffith, J.K. Homologous sugar-transport proteins in microbes and man. Biochem. Soc. Trans. 1993, 21, 1002–1006. [Google Scholar] [CrossRef]
- Saier, M.H., Jr. Families of transmembrane sugar transport proteins. Mol. Microbiol. 2000, 35, 699–710. [Google Scholar] [CrossRef]
- Kaback, H.R.; Guan, L. It takes two to tango: The dance of the permease. J. Gen. Physiol. 2019, 151, 878–886. [Google Scholar] [CrossRef] [Green Version]
- Levy, S.B. Active efflux mechanisms for antimicrobial resistance. Antimicrob. Agents Chemother. 1992, 36, 695–703. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Mukherjee, M.M.; Varela, M.F. Modulation of bacterial multidrug resistance efflux pumps of the major facilitator superfamily. Int. J. Bacteriol. 2013, 2013, 204141. [Google Scholar] [CrossRef] [Green Version]
- Ranaweera, I.; Shrestha, U.; Ranjana, K.C.; Kakarla, P.; Willmon, T.M.; Hernandez, A.J.; Mukherjee, M.M.; Barr, S.R.; Varela, M.F. Structural comparison of bacterial multidrug efflux pumps of the major facilitator superfamily. Trends Cell. Mol. Biol. 2015, 10, 131–140. [Google Scholar]
- Quistgaard, E.M.; Low, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell. Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Yan, N. Structural advances for the major facilitator superfamily (MFS) transporters. Trends Biochem. Sci. 2013, 38, 151–159. [Google Scholar] [CrossRef]
- Yin, Y.; He, X.; Szewczyk, P.; Nguyen, T.; Chang, G. Structure of the multidrug transporter EmrD from Escherichia coli. Science 2006, 312, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Henderson, P.J. Studies of translocation catalysis. Biosci. Rep. 1991, 11, 477–538. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Wright, S.H.; Tama, F. Simulations of substrate transport in the multidrug transporter EmrD. Proteins 2012, 80, 1620–1632. [Google Scholar] [CrossRef] [Green Version]
- Dang, S.; Sun, L.; Huang, Y.; Lu, F.; Liu, Y.; Gong, H.; Wang, J.; Yan, N. Structure of a fucose transporter in an outward-open conformation. Nature 2010, 467, 734–738. [Google Scholar] [CrossRef]
- Kumar, S.; Ranjana, K.; Sanford, L.M.; Hernandez, A.J.; Kakarla, P.; Varela, M.F. Structural and functional roles of two evolutionarily conserved amino acid sequence motifs within solute transporters of the major facilitator superfamily. Trends Cell. Mol. Biol. 2016, 11, 41–53. [Google Scholar]
- Rouch, D.A.; Cram, D.S.; DiBerardino, D.; Littlejohn, T.G.; Skurray, R.A. Efflux-mediated antiseptic resistance gene qacA from Staphylococcus aureus: Common ancestry with tetracycline- and sugar-transport proteins. Mol. Microbiol. 1990, 4, 2051–2062. [Google Scholar] [CrossRef]
- McMurry, L.; Petrucci, R.E.; Levy, S.B., Jr. Active efflux of tetracycline encoded by four genetically different tetracycline resistance determinants in Escherichia coli. Proc. Natl. Acad. Sci. USA 1980, 77, 3974–3977. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, A.; Ono, N.; Akasaka, T.; Noumi, T.; Sawai, T. Metal-tetracycline/H+ antiporter of Escherichia coli encoded by a transposon, Tn10. The role of the conserved dipeptide, Ser65-Asp66, in tetracycline transport. J. Biol. Chem. 1990, 265, 15525–15530. [Google Scholar]
- Kimura, T.; Shiina, Y.; Sawai, T.; Yamaguchi, A. Cysteine-scanning mutagenesis around transmembrane segment III of Tn10-encoded metal-tetracycline/H+ antiporter. J. Biol. Chem. 1998, 273, 5243–5247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, A.; Akasaka, T.; Kimura, T.; Sakai, T.; Adachi, Y.; Sawai, T. Role of the conserved quartets of residues located in the N- and C-terminal halves of the transposon Tn10-encoded metal-tetracycline/H+ antiporter of Escherichia coli. Biochemistry 1993, 32, 5698–5704. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Nakatani, M.; Kawabe, T.; Yamaguchi, A. Roles of conserved arginine residues in the metal-tetracycline/H+ antiporter of Escherichia coli. Biochemistry 1998, 37, 5475–5480. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Zhao, Y.; Wang, X.; Fan, J.; Heng, J.; Liu, X.; Feng, W.; Kang, X.; Huang, B.; Liu, J.; et al. Structure of the YajR transporter suggests a transport mechanism based on the conserved motif A. Proc. Natl. Acad. Sci. USA 2013, 110, 14664–14669. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.I.; Varela, M.F.; Wilson, T.H. Physiological evidence for an interaction between Glu-325 and His-322 in the lactose carrier of Escherichia coli. Biochim. Biophys. Acta 1996, 1278, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.I.; Hwang, P.P.; Hansen, C.; Wilson, T.H. Possible salt bridges between transmembrane alpha-helices of the lactose carrier of Escherichia coli. J. Biol. Chem. 1992, 267, 20758–20764. [Google Scholar]
- Masureel, M.; Martens, C.; Stein, R.A.; Mishra, S.; Ruysschaert, J.M.; McHaourab, H.S.; Govaerts, C. Protonation drives the conformational switch in the multidrug transporter LmrP. Nat. Chem. Biol. 2014, 10, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Bolhuis, H.; Poelarends, G.; van Veen, H.W.; Poolman, B.; Driessen, A.J.; Konings, W.N. The Lactococcal lmrP gene encodes a proton motive force-dependent drug transporter. J. Biol. Chem. 1995, 270, 26092–26098. [Google Scholar] [CrossRef] [Green Version]
- Marger, M.D.; Saier, M.H., Jr. A major superfamily of transmembrane facilitators that catalyse uniport, symport and antiport. Trends Biochem. Sci. 1993, 18, 13–20. [Google Scholar] [CrossRef]
- Varela, M.F.; Sansom, C.E.; Griffith, J.K. Mutational analysis and molecular modelling of an amino acid sequence motif conserved in antiporters but not symporters in a transporter superfamily. Mol. Membr. Biol. 1995, 12, 313–319. [Google Scholar] [CrossRef]
- Varela, M.F.; Griffith, J.K. Nucleotide and deduced protein sequences of the class D tetracycline resistance determinant: Relationship to other antimicrobial transport proteins. Antimicrob. Agents Chemother. 1993, 37, 1253–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaffe, D.; Radestock, S.; Shuster, Y.; Forrest, L.R.; Schuldiner, S. Identification of molecular hinge points mediating alternating access in the vesicular monoamine transporter VMAT2. Proc. Natl. Acad. Sci. USA 2013, 110, E1332–E1341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginn, S.L.; Brown, M.H.; Skurray, R.A. The TetA(K) tetracycline/H+ antiporter from Staphylococcus aureus: Mutagenesis and functional analysis of motif C. J. Bacteriol. 2000, 182, 1492–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, S.; Iwaki, S.; Kimura-Someya, T.; Yamaguchi, A. Cysteine-scanning mutagenesis around transmembrane segment VI of Tn10-encoded metal-tetracycline/H+ antiporter. FEBS Lett. 1999, 461, 315–318. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Krulwich, T.A. Site-directed mutagenesis studies of selected motif and charged residues and of cysteines of the multifunctional tetracycline efflux protein Tet(L). J. Bacteriol. 2002, 184, 1796–1800. [Google Scholar] [CrossRef] [Green Version]
- De Jesus, M.; Jin, J.; Guffanti, A.A.; Krulwich, T.A. Importance of the GP dipeptide of the antiporter motif and other membrane-embedded proline and glycine residues in tetracycline efflux protein Tet(L). Biochemistry 2005, 44, 12896–12904. [Google Scholar] [CrossRef] [Green Version]
- Saraceni-Richards, C.A.; Levy, S.B. Second-site suppressor mutations of inactivating substitutions at Gly247 of the tetracycline efflux protein, Tet(B). J. Bacteriol. 2000, 182, 6514–6516. [Google Scholar] [CrossRef] [Green Version]
- Saraceni-Richards, C.A.; Levy, S.B. Evidence for interactions between helices 5 and 8 and a role for the interdomain loop in tetracycline resistance mediated by hybrid Tet proteins. J. Biol. Chem. 2000, 275, 6101–6106. [Google Scholar] [CrossRef] [Green Version]
- Hassan, K.A.; Galea, M.; Wu, J.; Mitchell, B.A.; Skurray, R.A.; Brown, M.H. Functional effects of intramembranous proline substitutions in the staphylococcal multidrug transporter QacA. FEMS Microbiol. Lett. 2006, 263, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Pasrija, R.; Banerjee, D.; Prasad, R. Structure and function analysis of CaMdr1p, a major facilitator superfamily antifungal efflux transporter protein of Candida albicans: Identification of amino acid residues critical for drug/H+ transport. Eukaryot. Cell. 2007, 6, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Parsons, S.M. Conformational propensities of peptides mimicking transmembrane helix 5 and motif C in wild-type and mutant vesicular acetylcholine transporters. ACS Chem. Neurosci. 2010, 1, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Lekshmi, M.; Ammini, P.; Adjei, J.; Sanford, L.M.; Shrestha, U.; Kumar, S.; Varela, M.F. Modulation of antimicrobial efflux pumps of the major facilitator superfamily in Staphylococcus aureus. AIMS Microbiol. 2017, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, U.; Lekshmi, M.; Kumar, S.; Adjei, J.; Jones, K.M.; Hernandez, A.J.; Sanford, L.M.; Varela, M.F. Bioactive agents as modulators of multidrug efflux pumps from the major facilitator superfamily in key bacterial pathogens. Curr. Trends Microbiol. 2018, 12, 15–37. [Google Scholar]
- Rao, M.; Padyana, S.; Dipin, K.; Kumar, S.; Nayak, B.; Varela, M.F. Antimicrobial compounds of plant origin as efflux pump inhibitors: New avenues for controlling multidrug resistant pathogens. J. Antimicrob. Agents 2018, 4, 1000159. [Google Scholar] [CrossRef]
- Bhardwaj, A.K.; Mohanty, P. Bacterial efflux pumps involved in multidrug resistance and their inhibitors: Rejuvinating the antimicrobial chemotherapy. Recent. Pat. Anti Infect. Drug Discov. 2012, 7, 73–89. [Google Scholar] [CrossRef]
- Cheesman, M.J.; Ilanko, A.; Blonk, B.; Cock, I.E. Developing new antimicrobial therapies: Are synergistic combinations of plant extracts/compounds with conventional antibiotics the solution? Pharmacogn. Rev. 2017, 11, 57–72. [Google Scholar]
- Barrero, M.A.; Pietralonga, P.A.; Schwarz, D.G.; Silva, A., Jr.; Paula, S.O.; Moreira, M.A. Effect of the inhibitors phenylalanine arginyl ß-naphthylamide (PAßN) and 1-(1-naphthylmethyl)-piperazine (NMP) on expression of genes in multidrug efflux systems of Escherichia coli isolates from bovine mastitis. Res. Vet. Sci. 2014, 97, 176–181. [Google Scholar] [CrossRef]
- Bruns, M.M.; Kakarla, P.; Floyd, J.T.; Mukherjee, M.M.; Ponce, R.C.; Garcia, J.A.; Ranaweera, I.; Sanford, L.M.; Hernandez, A.J.; Willmon, T.M.; et al. Modulation of the multidrug efflux pump EmrD-3 from Vibrio cholerae by Allium sativum extract and the bioactive agent allyl sulfide plus synergistic enhancement of antimicrobial susceptibility by A sativum extract. Arch. Microbiol. 2017, 199, 1103–1112. [Google Scholar] [CrossRef]
- Putman, M.; Koole, L.A.; van Veen, H.W.; Konings, W.N. The secondary multidrug transporter LmrP contains multiple drug interaction sites. Biochemistry 1999, 38, 13900–13905. [Google Scholar] [CrossRef]
- Dymek, A.; Armada, A.; Handzlik, J.; Viveiros, M.; Spengler, G.; Molnar, J.; Kiec-Kononowicz, K.; Amaral, L. The activity of 16 new hydantoin compounds on the intrinsic and overexpressed efflux pump system of Staphylococcus aureus. In Vivo 2012, 26, 223–229. [Google Scholar]
- Wang, S.Y.; Sun, Z.L.; Liu, T.; Gibbons, S.; Zhang, W.J.; Qing, M. Flavonoids from Sophora moorcroftiana and their synergistic antibacterial effects on MRSA. Phytother. Res. 2014, 28, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Heng, J.; Gao, Y.; Wang, X. Crystal structures of MdfA complexed with acetylcholine and inhibitor reserpine. Biophys. Rep. 2016, 2, 78–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakarla, P.; Floyd, J.; Mukherjee, M.; Devireddy, A.R.; Inupakutika, M.A.; Ranweera, I.; Kc, R.; Shrestha, U.; Cheeti, U.R.; Willmon, T.M.; et al. Inhibition of the multidrug efflux pump LmrS from Staphylococcus aureus by cumin spice Cuminum cyminum. Arch. Microbiol. 2017, 199, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Gracio, M.A.; Gracio, A.J.; Viveiros, M.; Amaral, L. Since phenothiazines alter antibiotic susceptibility of microorganisms by inhibiting efflux pumps, are these agents useful for evaluating similar pumps in phenothiazine-sensitive parasites? Int. J. Antimicrob. Agents 2003, 22, 347–351. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Lorenz, P.; Tawara, J.N.; Zenewicz, L.A.; Lewis, K. Synergy in a medicinal plant: Antimicrobial action of berberine potentiated by 5’-methoxyhydnocarpin, a multidrug pump inhibitor. Proc. Natl. Acad. Sci. USA 2000, 97, 1433–1437. [Google Scholar] [CrossRef] [Green Version]
- Kaatz, G.W.; Seo, S.M. Inducible NorA-mediated multidrug resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1995, 39, 2650–2655. [Google Scholar] [CrossRef] [Green Version]
- Aeschlimann, J.R.; Dresser, L.D.; Kaatz, G.W.; Rybak, M.J. Effects of NorA inhibitors on in vitro antibacterial activities and postantibiotic effects of levofloxacin, ciprofloxacin, and norfloxacin in genetically related strains of Staphylococcus aureus. Antimicrob. Agents Chemother. 1999, 43, 335–340. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, S.; Oluwatuyi, M.; Kaatz, G.W. A novel inhibitor of multidrug efflux pumps in Staphylococcus aureus. J. Antimicrob. Chemother. 2003, 51, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Leitner, I.; Nemeth, J.; Feurstein, T.; Abrahim, A.; Matzneller, P.; Lagler, H.; Erker, T.; Langer, O.; Zeitlinger, M. The third-generation P-glycoprotein inhibitor tariquidar may overcome bacterial multidrug resistance by increasing intracellular drug concentration. J. Antimicrob. Chemother. 2011, 66, 834–839. [Google Scholar] [CrossRef] [Green Version]
- Ng, E.Y.; Trucksis, M.; Hooper, D.C. Quinolone resistance mediated by norA: Physiologic characterization and relationship to flqB, a quinolone resistance locus on the Staphylococcus aureus chromosome. Antimicrob. Agents Chemother. 1994, 38, 1345–1355. [Google Scholar] [CrossRef] [Green Version]
- Michalet, S.; Cartier, G.; David, B.; Mariotte, A.M.; Dijoux-franca, M.G.; Kaatz, G.W.; Stavri, M.; Gibbons, S. N-Caffeoylphenalkylamide derivatives as bacterial efflux pump inhibitors. Bioorg. Med. Chem. Lett. 2007, 17, 1755–1758. [Google Scholar] [CrossRef]
- Couto, I.; Costa, S.S.; Viveiros, M.; Martins, M.; Amaral, L. Efflux-mediated response of Staphylococcus aureus exposed to ethidium bromide. J. Antimicrob. Chemother 2008, 62, 504–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchi, E.; Furi, L.; Arioli, S.; Morrissey, I.; Di Lorenzo, V.; Mora, D.; Giovannetti, L.; Oggioni, M.R.; Viti, C. Novel insight into antimicrobial resistance and sensitivity phenotypes associated to qac and norA genotypes in Staphylococcus aureus. Microbiol. Res. 2015, 170, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Kaatz, G.W.; Moudgal, V.V.; Seo, S.M.; Hansen, J.B.; Kristiansen, J.E. Phenylpiperidine selective serotonin reuptake inhibitors interfere with multidrug efflux pump activity in Staphylococcus aureus. Int. J. Antimicrob. Agents 2003, 22, 254–261. [Google Scholar] [CrossRef]
- Holler, J.G.; Christensen, S.B.; Slotved, H.C.; Rasmussen, H.B.; Guzman, A.; Olsen, C.E.; Petersen, B.; Molgaard, P. Novel inhibitory activity of the Staphylococcus aureus NorA efflux pump by a kaempferol rhamnoside isolated from Persea lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [Green Version]
- Holler, J.G.; Slotved, H.C.; Molgaard, P.; Olsen, C.E.; Christensen, S.B. Chalcone inhibitors of the NorA efflux pump in Staphylococcus aureus whole cells and enriched everted membrane vesicles. Bioorg. Med. Chem. 2012, 20, 4514–4521. [Google Scholar] [CrossRef]
- Sabatini, S.; Gosetto, F.; Serritella, S.; Manfroni, G.; Tabarrini, O.; Iraci, N.; Brincat, J.P.; Carosati, E.; Villarini, M.; Kaatz, G.W.; et al. Pyrazolo [4,3-c][1,2] benzothiazines 5,5-dioxide: A promising new class of Staphylococcus aureus NorA efflux pump inhibitors. J. Med. Chem. 2012, 55, 3568–3572. [Google Scholar] [CrossRef]
- Roy, S.K.; Kumari, N.; Pahwa, S.; Agrahari, U.C.; Bhutani, K.K.; Jachak, S.M.; Nandanwar, H. NorA efflux pump inhibitory activity of coumarins from Mesua ferrea. Fitoterapia 2013, 90, 140–150. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, Y.; Wang, Y.; Lu, M.; He, J.; Liu, J.; Chen, Q.; Zhang, X.; Zhou, F.; Wang, G.; et al. Non-antibiotic agent ginsenoside 20(S)-Rh2 enhanced the antibacterial effects of ciprofloxacin in vitro and in vivo as a potential NorA inhibitor. Eur. J. Pharmacol. 2014, 740, 277–284. [Google Scholar] [CrossRef]
- Fontaine, F.; Hequet, A.; Voisin-Chiret, A.S.; Bouillon, A.; Lesnard, A.; Cresteil, T.; Jolivalt, C.; Rault, S. First identification of boronic species as novel potential inhibitors of the Staphylococcus aureus NorA efflux pump. J. Med. Chem. 2014, 57, 2536–2548. [Google Scholar] [CrossRef]
- Wang, D.; Xie, K.; Zou, D.; Meng, M.; Xie, M. Inhibitory effects of silybin on the efflux pump of methicillin resistant Staphylococcus aureus. Mol. Med. Rep. 2018, 18, 827–833. [Google Scholar] [PubMed]
- Sharma, A.; Gupta, V.K.; Pathania, R. Efflux pump inhibitors for bacterial pathogens: From bench to bedside. Indian J. Med. Res. 2019, 149, 129–145. [Google Scholar] [PubMed]
- Coelho, M.L.; Ferreira, J.H.; de Siqueira Junior, J.P.; Kaatz, G.W.; Barreto, H.M.; de Carvalho Melo Cavalcante, A.A. Inhibition of the NorA multi-drug transporter by oxygenated monoterpenes. Microb. Pathog. 2016, 99, 173–177. [Google Scholar] [CrossRef]
- Costa, L.M.; de Macedo, E.V.; Oliveira, F.A.; Ferreira, J.H.; Gutierrez, S.J.; Pelaez, W.J.; Lima, F.C.; de Siqueira Junior, J.P.; Coutinho, H.D.; Kaatz, G.W.; et al. Inhibition of the NorA efflux pump of Staphylococcus aureus by synthetic riparins. J. Appl. Microbiol. 2016, 121, 1312–1322. [Google Scholar] [CrossRef]
- Martins, A.; Vasas, A.; Viveiros, M.; Molnar, J.; Hohmann, J.; Amaral, L. Antibacterial properties of compounds isolated from Carpobrotus edulis. Int. J. Antimicrob. Agents 2011, 37, 438–444. [Google Scholar] [CrossRef] [Green Version]
- De Sousa Andrade, L.M.; de Oliveira, A.B.M.; Leal, A.; de Alcantara Oliveira, F.A.; Portela, A.L.; de Sousa Lima Neto, J.; de Siqueira-Junior, J.P.; Kaatz, G.W.; da Rocha, C.Q.; Barreto, H.M. Antimicrobial activity and inhibition of the NorA efflux pump of Staphylococcus aureus by extract and isolated compounds from Arrabidaea brachypoda. Microb. Pathog. 2019, 140, 103935. [Google Scholar] [CrossRef]
- McMurry, L.M.; Cullinane, J.C.; Petrucci, R.E., Jr.; Levy, S.B. Active uptake of tetracycline by membrane vesicles from susceptible Escherichia coli. Antimicrob. Agents Chemother. 1981, 20, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Varela, M.F. Biochemistry of bacterial multidrug efflux pumps. Int. J. Mol. Sci. 2012, 13, 4484–4495. [Google Scholar] [CrossRef]
- Ughachukwu, P.; Unekwe, P. Efflux pump-mediated resistance in chemotherapy. Ann. Med. Health Sci. Res. 2012, 2, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Piddock, L.J. Mechanisms of fluoroquinolone resistance: An update 1994–1998. Drugs 1999, 58 (Suppl. 2), 11–18. [Google Scholar] [CrossRef]
- Hegeto, L.A.; Caleffi-Ferracioli, K.R.; Perez de Souza, J.; Almeida, A.L.; Nakamura de Vasconcelos, S.S.; Barros, I.L.E.; Canezin, P.H.; Campanerut-Sa, P.A.Z.; Scodro, R.B.L.; Siqueira, V.L.D.; et al. Promising antituberculosis activity of piperine combined with antimicrobials: A systematic review. Microb. Drug Resist. 2019, 25, 120–126. [Google Scholar] [PubMed]
- Lamut, A.; Peterlin Masic, L.; Kikelj, D.; Tomasic, T. Efflux pump inhibitors of clinically relevant multidrug resistant bacteria. Med. Res. Rev. 2019, 39, 2460–2504. [Google Scholar] [PubMed]
- Handzlik, J.; Matys, A.; Kiec-Kononowicz, K. Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of Gram-positive bacteria S. aureus. Antibiotics 2013, 2, 28–45. [Google Scholar] [CrossRef] [Green Version]
- Saidijam, M.; Benedetti, G.; Ren, Q.; Xu, Z.; Hoyle, C.J.; Palmer, S.L.; Ward, A.; Bettaney, K.E.; Szakonyi, G.; Meuller, J.; et al. Microbial drug efflux proteins of the major facilitator superfamily. Curr. Drug Targets 2006, 7, 793–811. [Google Scholar] [CrossRef]
- Lewis, K. In search of natural substrates and inhibitors of MDR pumps. J. Mol. Microbiol. Biotechnol. 2001, 3, 247–254. [Google Scholar]
- Floyd, J.L.; Smith, K.P.; Kumar, S.H.; Floyd, J.T.; Varela, M.F. LmrS is a multidrug efflux pump of the major facilitator superfamily from Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 5406–5412. [Google Scholar] [CrossRef] [Green Version]
- Hassanzadeh, S.; Ganjloo, S.; Pourmand, M.R.; Mashhadi, R.; Ghazvini, K. Epidemiology of efflux pumps genes mediating resistance among Staphylococcus aureus: A systematic review. Microb. Pathog. 2019, 139, 103850. [Google Scholar] [CrossRef]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef]
- Jang, S. Multidrug efflux pumps in Staphylococcus aureus and their clinical implications. J. Microbiol. 2016, 54, 1–8. [Google Scholar] [CrossRef]
- Heng, J.; Zhao, Y.; Liu, M.; Liu, Y.; Fan, J.; Wang, X.; Zhang, X.C. Substrate-bound structure of the E. coli multidrug resistance transporter MdfA. Cell Res 2015, 25, 1060–1073. [Google Scholar]
- Nagarathinam, K.; Nakada-Nakura, Y.; Parthier, C.; Terada, T.; Juge, N.; Jaenecke, F.; Liu, K.; Hotta, Y.; Miyaji, T.; Omote, H.; et al. Outward open conformation of a Major Facilitator Superfamily multidrug/H+ antiporter provides insights into switching mechanism. Nat. Commun. 2018, 9, 4005. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [Green Version]
- Beceiro, A.; Tomas, M.; Bou, G. Antimicrobial resistance and virulence: A successful or deleterious association in the bacterial world? Clin. Microbiol. Rev. 2013, 26, 185–230. [Google Scholar] [CrossRef] [Green Version]
- Croucher, N.J.; Hanage, W.P.; Harris, S.R.; McGee, L.; van der Linden, M.; de Lencastre, H.; Sa-Leao, R.; Song, J.H.; Ko, K.S.; Beall, B.; et al. Variable recombination dynamics during the emergence, transmission and disarming of a multidrug-resistant pneumococcal clone. BMC Biol. 2014, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, M.; Kakarla, P.; Kumar, S.; Gonzalez, E.; Floyd, J.T.; Inupakutika, M.; Devireddy, A.R.; Tirrell, S.R.; Bruns, M.; He, G.; et al. Comparative genome analysis of non-toxigenic non-O1 versus toxigenic O1 Vibrio cholerae. Genom. Discov. 2014, 2, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Lindquist, I.E.; Sundararajan, A.; Rajanna, C.; Floyd, J.T.; Smith, K.P.; Andersen, J.L.; He, G.; Ayers, R.M.; Johnson, J.A.; et al. Genome Sequence of Non-O1 Vibrio cholerae PS15. Genome Announc. 2013, 1, e00227-12. [Google Scholar] [CrossRef] [Green Version]
- Heidelberg, J.F.; Eisen, J.A.; Nelson, W.C.; Clayton, R.A.; Gwinn, M.L.; Dodson, R.J.; Haft, D.H.; Hickey, E.K.; Peterson, J.D.; Umayam, L.; et al. DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature 2000, 406, 477–483. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Efflux Pump Targeted | Modulators | References |
|---|---|---|
| EmrB from Escherichia coli | Phenylalanine arginyl β-naphthylamide (PAβN) and 1-(1-naphthyl methyl)-piperazine (NMP) | [127] |
| EmrD-3 from Vibrio cholerae | Garlic, allyl sulfide | [128] |
| LmrP from Lactococcus lactis | Verapamil and quinine Nicardipine and vinblastine Tetraphenyl phosphonium | [129] |
| QacA from Staphylococcus aureus | Hydantoin, silybin | [130,131] |
| MdfA from Escherichia coli | Reserpine | [132] |
| QacB from Staphylococcus aureus | Silybin | [131] |
| LmrS from Staphylococcus aureus | Cumin seed oil, cumin aldehyde, reserpine | [133] |
| NorA from Staphylococcus aureus | 3-aryl piperidines | [134] |
| Berberine | [135] | |
| Reserpine | [136] | |
| Omeprazole, lansoprazole | [137] | |
| GG918, tariquidar (primary active transport inhibitors) | [138,139] | |
| Verapamil, ciprofloxacin, ofloxacin | [140] | |
| 5,9′dimethyl-deca-2,4,8-trienoic acid, 9-formyl-5-methyl-deca-2,4,8-trienoic acid | [141] | |
| Chlorpromazine, thioridazine, and prochlorperazine | [142,143,144] | |
| Kaempferol rhamnoside | [145] | |
| Chalones | [146] | |
| COX-2 inhibitor analog, 3-(4-chlorophenyl)-1-(4-nitrophenyl)-1,4-dihydropyrazolo[4,3-c] [1,2] benzothiazine 5,5-dioxide | [147] | |
| Coumarin | [148] | |
| Genistein (flavonoid compound) | [131] | |
| Ginsenoside 20(S)-Rh2 | [149] | |
| Boronic acid molecules, 6-(3-phenylpropoxy) pyridine-3-boronic acid and 6-(4-phenylbutoxy) pyridine-3- boronic acid | [150] | |
| Silybin | [151] | |
| 5′-methoxy-hydnocarpin, pheophorbide A, 5′-MHC, curcumin, kaempferol, silibinin, isoflavone, orizabins, capsaicin, tannic acid, | [152] | |
| nerol, dimethyl octanol, estragole | [153] | |
| Riparin B | [154] | |
| Olaanolic acid, ulvaol | [155] | |
| Brachydins: BR-A, BR-B | [156] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, S.; Lekshmi, M.; Parvathi, A.; Ojha, M.; Wenzel, N.; Varela, M.F. Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps. Microorganisms 2020, 8, 266. https://doi.org/10.3390/microorganisms8020266
Kumar S, Lekshmi M, Parvathi A, Ojha M, Wenzel N, Varela MF. Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps. Microorganisms. 2020; 8(2):266. https://doi.org/10.3390/microorganisms8020266
Chicago/Turabian StyleKumar, Sanath, Manjusha Lekshmi, Ammini Parvathi, Manisha Ojha, Nicholas Wenzel, and Manuel F. Varela. 2020. "Functional and Structural Roles of the Major Facilitator Superfamily Bacterial Multidrug Efflux Pumps" Microorganisms 8, no. 2: 266. https://doi.org/10.3390/microorganisms8020266