Managed and Unmanaged Pinus sylvestris Forest Stands Harbour Similar Diversity and Composition of the Phyllosphere and Soil Fungi

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
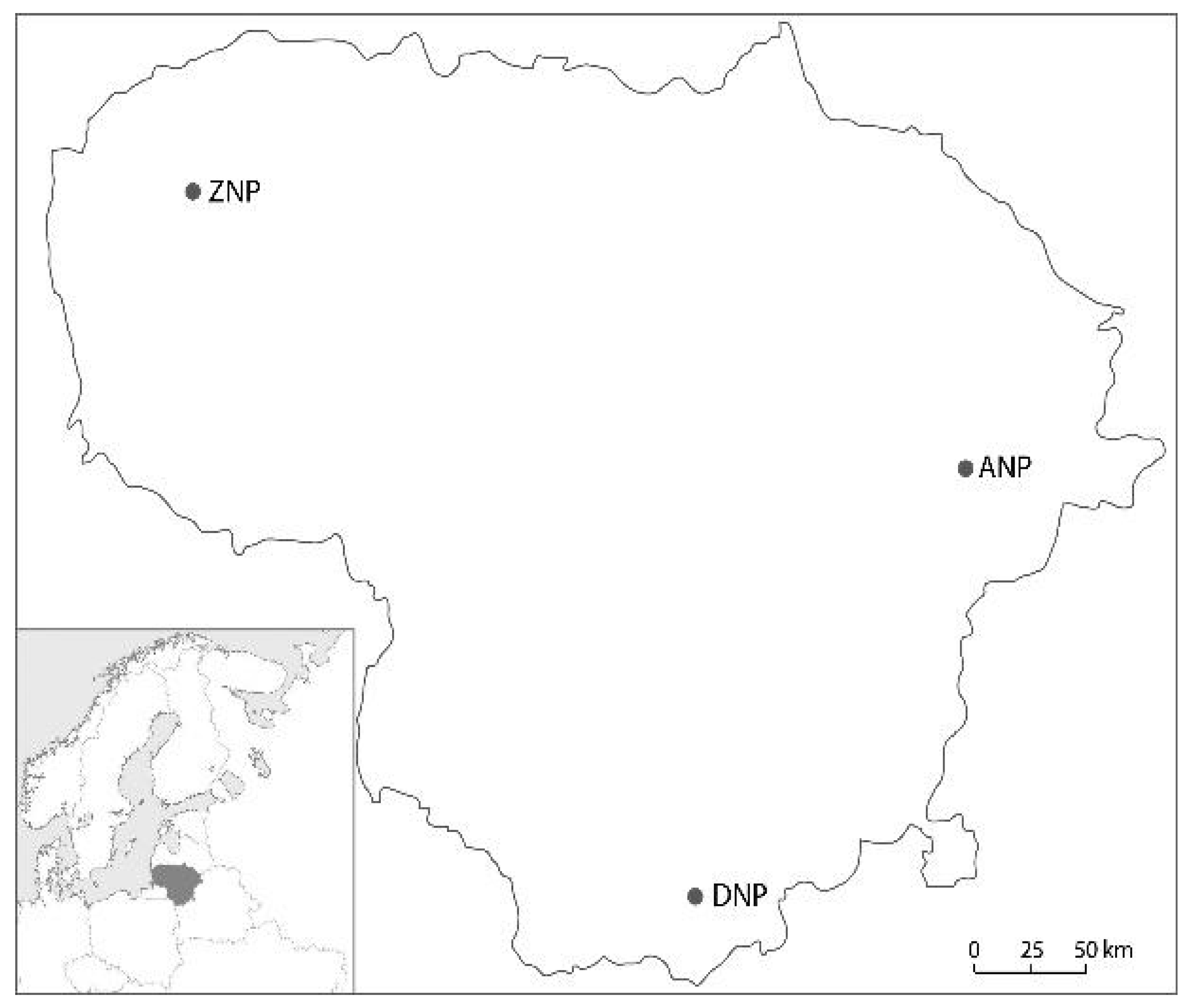
2.1. Study Sites and Sampling
2.2. DNA Isolation and Sequencing
2.3. Bioinformatics
2.4. Statistical Analyses
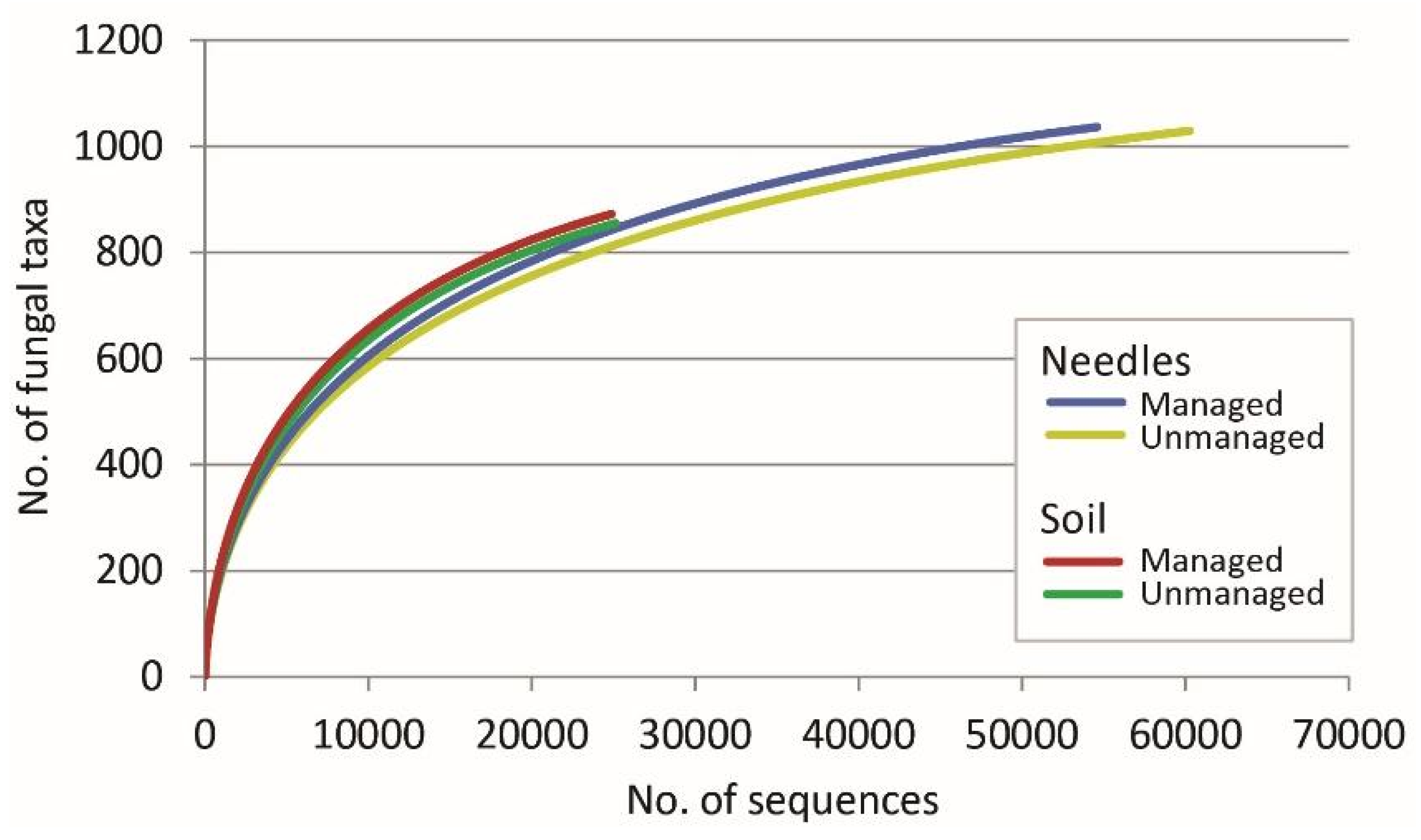
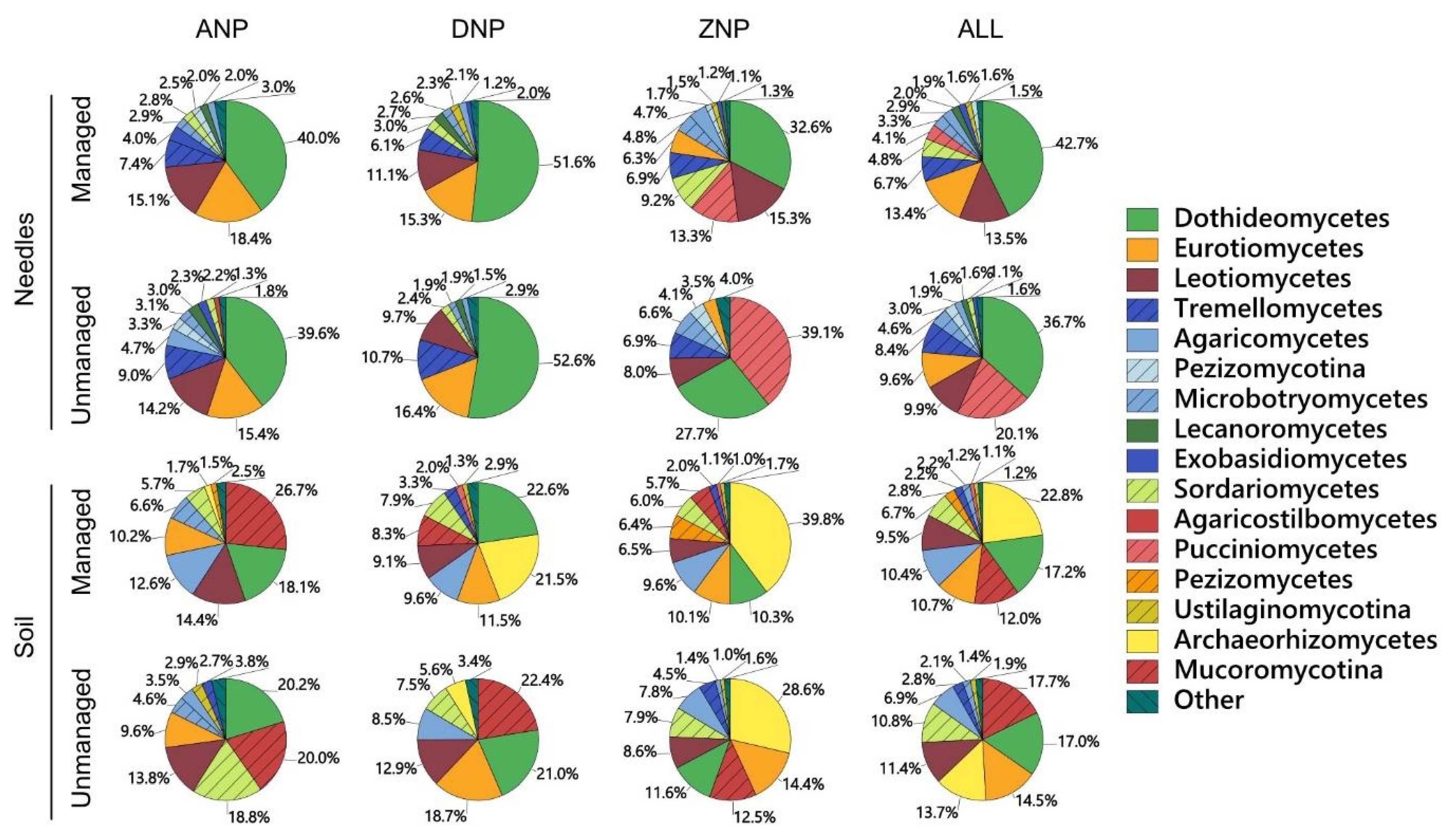
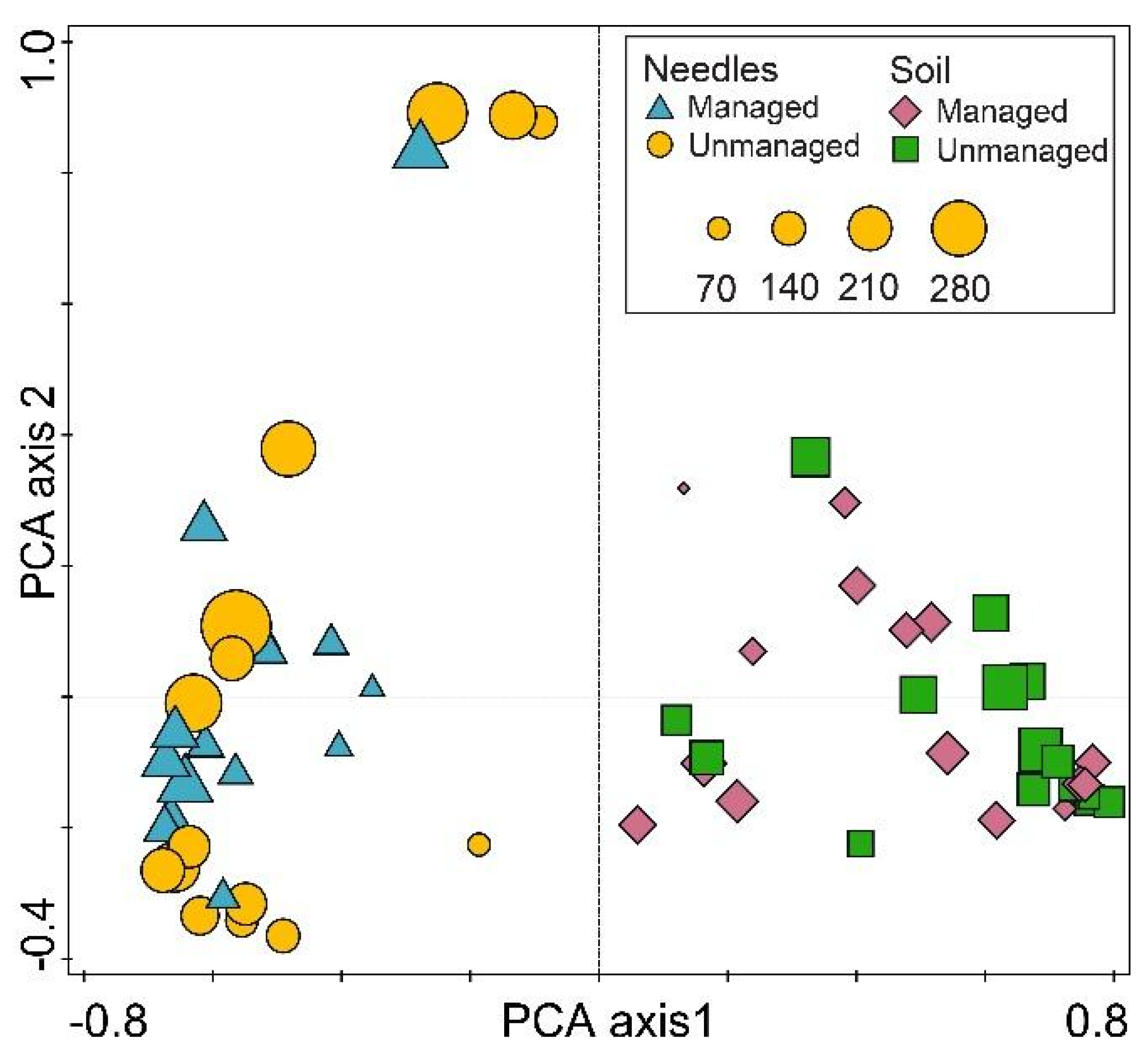
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gustafsson, L.; Baker, S.C.; Bauhus, J.; Beese, W.J.; Brodie, A.; Kouki, J.; Lindenmayer, D.B.; Lõhmus, A.; Pastur, G.M.; Messier, C.; et al. Retention forestry to maintain multifunctional forests: A world perspective. BioScience 2012, 62, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Kuuluvainen, T. Forest management and biodiversity conservation based on natural ecosystem dynamics in northern Europe: the complexity challenge. Ambio 2009, 38, 309–315. [Google Scholar] [CrossRef]
- Chaudhary, A.; Burivalova, Z.; Koh, L.P.; Hellweg, S. Impact of forest management on species richness: global meta-analysis and economic trade-offs. Sci. Rep. 2016, 6, 23954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horák, J.; Materna, J.; Halda, J.P.; Mladenović, S.; Bogusch, P.; Pech, P. Biodiversity in remnants of natural mountain forests under conservation-oriented management. Sci. Rep. 2019, 9, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paillet, Y.; Bergès, L.; Hjälén, J.; Ódor, P.; Avon, C.; Bernhardt-Römermann, M.; Bijlsma, R.-J.; De Bruyn, L.; Fuhr, M.; Grandin, U.; et al. Biodiversity differences between managed and unmanaged forests: meta-analysis of species richness in Europe. Conserv. Biol. 2010, 24, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Elek, Z.; Kovács, B.; Aszalós, R.; Boros, G.; Samu, F.; Tinya, F.; Ódor, P. Taxon-specific responses to different forestry treatments in a temperate forest. Sci. Rep. 2018, 8, 16990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pioli, S.; Antonucci, S.; Giovannelli, A.; Traversi, M.L.; Borruso, L.; Bani, A.; Brusetti, L.; Tognetti, R. Community fingerprinting reveals increasing wood-inhabiting fungal diversity in unmanaged Mediterranean forests. For. Ecol. Manag. 2018, 408, 202–210. [Google Scholar] [CrossRef]
- Lombardi, F.; Lasserre, B.; Chirici, G.; Tognetti, R.; Marchetti, M. Deadwood occurrence and forest structure as indicators of old-growth forest conditions in Mediterranean mountainous ecosystems. Écoscience 2012, 19, 344–355. [Google Scholar] [CrossRef]
- Kuuluvainen, T. Disturbance dynamics in boreal forests: defining the ecological basis of restoration and management of biodiversity. Silva Fenn. 2002, 36, 5–11. [Google Scholar] [CrossRef]
- Lange, M.; Türke, M.; Pašalić, E.; Boch, S.; Hessenmöller, D.; Müller, J.; Prati, D.; Socher, S.A.; Fischer, M.; Weisser, W.W.; et al. Effects of forest management on ground-dwelling beetles (Coleoptera; Carabidae, Staphylinidae) in Central Europe are mainly mediated by changes in forest structure. For. Ecol. Manag. 2014, 329, 166–176. [Google Scholar] [CrossRef]
- Villa-Castillo, J. Ground beetle (Coleoptera: Carabidae) species assemblage as an indicator of forest condition in northern Arizona ponderosa pine forests. Environ. Entomol. 2002, 31, 242–252. [Google Scholar] [CrossRef]
- Allegro, G.; Sciaky, R. Assessing the potential role of ground beetles (Coleoptera, Carabidae) as bioindicators in poplar stands, with a newly proposed ecological index (FAI). For. Ecol. Manag. 2003, 175, 275–284. [Google Scholar] [CrossRef]
- Fuller, R.J.; Oliver, T.H.; Leather, S.R. Forest management effects on carabid beetle communities in coniferous and broadleaved forests: implications for conservation. Insect. Conserv. Divers. 2008, 1, 242–252. [Google Scholar] [CrossRef]
- Koivula, M. Useful model organisms, indicators, or both? Ground beetles (Coleoptera, Carabidae) reflecting environmental conditions. ZooKeys 2011, 100, 287–317. [Google Scholar] [CrossRef] [Green Version]
- Gernandt, D.S.; López, G.G.; García, S.O.; Liston, A. Phylogeny and classification of Pinus. Taxon 2005, 54, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Ganopoulos, I.; Aravanopoulos, F.; Madesis, P.; Pasentsis, K.; Bosmali, I.; Ouzounis, C.; Tsaftaris, A. Taxonomic identification of mediterranean pines and their hybrids based on the high resolution melting (HRM) and trnL approaches: from cytoplasmic inheritance to timber tracing. PLoS ONE 2013, 8, e60945. [Google Scholar] [CrossRef] [Green Version]
- Millberg, H.; Boberg, J.; Stenlid, J. Changes in fungal community of Scots pine (Pinus sylvestris) needles along a latitudinal gradient in Sweden. Fungal Ecol. 2015, 17, 126–139. [Google Scholar] [CrossRef]
- Koukol, O. Linking Biodiversity and Ecology of Fungi from Pine and Spruce Needles; Charles University in Prague: Prague, Czech Republic, 2016. [Google Scholar]
- Felton, A.; Petersson, L.; Nilsson, O.; Witzell, J.; Cleary, M.; Felton, A.M.; Björkman, C.; Sang, Å.O.; Jonsell, M.; Holmström, E.; et al. The tree species matters: biodiversity and ecosystem service implications of replacing Scots pine production stands with Norway spruce. Ambio 2019, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Lust, N.; Muys, B.; Nachtergale, L. Increase of biodiversity in homogeneous Scots pine stands by an ecologically diversified management. Biodivers. Conserv. 1998, 7, 249–260. [Google Scholar] [CrossRef]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 486. [Google Scholar] [CrossRef] [Green Version]
- Millberg, H. Foliar fungi of Scots pine (Pinus sylvestris). Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2015. [Google Scholar]
- Goldmann, K.; Schoning, I.; Buscot, F.; Wubet, T. Forest management type influences diversity and community composition of soil fungi across temperate forest ecosystems. Front. Microbiol. 2015, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterkenburg, E.; Bahr, A.; Brandström Durling, M.; Clemmensen, K.E.; Lindahl, B.D. Changes in fungal communities along a boreal forest soil fertility gradient. New Phytol. 2015, 207, 1145–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: London, UK, 1997; p. 605. [Google Scholar]
- Nguyen, D.; Boberg, J.; Ihrmark, K.; Stenstrom, E.; Stenlid, J. Do foliar fungal communities of Norway spruce shift along a tree species diversity gradient in mature European forests? Fungal Ecol. 2016, 23, 97–108. [Google Scholar] [CrossRef] [Green Version]
- Gömöryová, E.; Ujházy, K.; Martinák, M.; Gömöry, D. Soil microbial community response to variation in vegetation and abiotic environment in a temperate old-growth forest. Appl. Soil Ecol. 2013, 68, 10–19. [Google Scholar] [CrossRef]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 28, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menkis, A.; Burokienė, D.; Gaitnieks, T.; Uotila, A.; Johannesson, H.; Rosling, A.; Finlay, R.D.; Stenlid, J.; Vasaitis, R. Occurrence and impact of the root-rot biocontrol agent Phlebiopsis gigantea on soil fungal communities in Picea abies forests of northern Europe. FEMS Microbiol. Ecol. 2012, 81, 438–445. [Google Scholar] [CrossRef] [Green Version]
- Menkis, A.; Marčiulynas, A.; Gedminas, A.; Lynikienė, J.; Povilaitienė, A. High-throughput sequencing reveals drastic changes in fungal communities in the phyllosphere of Norway spruce (Picea abies) following invasion of the spruce bud scale (Physokermes piceae). Microb. Ecol. 2015, 70, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, B.D.; Nilsson, R.H.; Tedersoo, L.; Abarenkov, K.; Carlsen, T.; Kjøller, R.; Kõljalg, U.; Pennanen, T.; Rosendahl, S.; Stenlid, J.; et al. Fungal community analysis by high-throughput sequencing of amplified markers--a user’s guide. New Phytol. 2013, 199, 288–299. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Tooming-Klunderud, A.; Anslan, S. PacBio metabarcoding of Fungi and other eukaryotes: errors, biases and perspectives. New Phytol. 2018, 217, 1370–1385. [Google Scholar] [CrossRef] [Green Version]
- Lithuanian Statistical Yearbook of Forestry. Available online: http://www.amvmt.lt/images/veikla/stat/miskustatistika/2016/02%20Misku%20ukio%20statistika%202016_m.pdf (accessed on 17 December 2019).
- Vaičys, M. Miško dirvožemių klasifikacija. In Lietuvos Dirvožemiai [Forest Site Types. Lithuanian Soils]; Mokslas Publishers: Vilnius, Lithuania, 2001; pp. 1040–1043. (In Lithuanian) [Google Scholar]
- Karazija, S. Lietuvos Mišku Tipai [Lithuanian Forest Typology]; Mokslas Publishers: Vilnius, Lithuania, 1988. (In Lithuanian) [Google Scholar]
- Menkis, A.; Burokienė, D.; Stenlid, J.; Stenström, E. High-throughput sequencing shows high fungal diversity and community segregation in the rhizospheres of container-grown conifer seedlings. Forests 2016, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Ihrmark, K.; Bodeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2 region - evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; p. 192. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- ter Braak, C.J.F.; Smilauer, P. Canoco Reference Manual and User’s Guide to Canoco for Windows: Software for Canonical Community Ordination, Version 4; Microcomputer Power: Ithaca, NY, USA, 1998; p. 351. [Google Scholar]
- Parlade, J.; Queralt, M.; Pera, J.; Bonet, J.A.; Castano, C.; Martinez-Pena, F.; Pinol, J.; Senar, M.A.; De Miguel, A.M. Temporal dynamics of soil fungal communities after partial and total clear-cutting in a managed Pinus sylvestris stand. For. Ecol. Manag. 2019, 449, 10. [Google Scholar] [CrossRef]
- Lilja, S.; Kuuluvainen, T. Structure of old Pinus sylvestris dominated forest stands along a geographic and human impact gradient in mid-boreal Fennoscandia. Silva Fenn. 2005, 39, 407–428. [Google Scholar] [CrossRef] [Green Version]
- Aime, M.C.; Matheny, P.B.; Henk, D.A.; Frieders, E.M.; Nilsson, R.H.; Piepenbring, M.; McLaughlin, D.J.; Szabo, L.J.; Begerow, D.; Sampaio, J.P.; et al. An overview of the higher level classification of Pucciniomycotina based on combined analyses of nuclear large and small subunit rDNA sequences. Mycologia 2006, 98, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Menkis, A.; Urbina, H.; James, T.Y.; Rosling, A. Archaeorhizomyces borealis sp. nov. and a sequence-based classification of related soil fungal species. Fungal Biol. 2014, 118, 943–955. [Google Scholar] [CrossRef]
- Rosling, A.; Cox, F.; Cruz-Martinez, K.; Ihrmark, K.; Grelet, G.A.; Lindahl, B.D.; Menkis, A.; James, T.Y. Archaeorhizomycetes: unearthing an ancient class of ubiquitous soil fungi. Science 2011, 333, 876–879. [Google Scholar] [CrossRef]
- Helfer, S. Coleosporium in Europe. Mycotaxon 2013, 124, 87–99. [Google Scholar] [CrossRef]
- Karadžić, D.; Milijašević, T. тhe most important parasitic and saprophytic fungi in Austrian pine and Scots pine plantations in Serbia. Bull. Fac. For. 2008, 97, 147–170. [Google Scholar] [CrossRef]
- Pirttila, A.M.; Pospiech, H.; Laukkanen, H.; Myllyla, R.; Hohtola, A. Two endophytic fungi in different tissues of scots pine buds (Pinus sylvestris L.). Microb. Ecol. 2003, 45, 53–62. [Google Scholar] [CrossRef]
- Talgo, V.; Chastagner, G.; Thomsen, I.M.; Cech, T.; Riley, K.; Lange, K.; Klemsdal, S.S.; Stensvand, A. Sydowia polyspora associated with current season needle necrosis (CSNN) on true fir (Abies spp.). Fungal Biol. 2010, 114, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Davydenko, K.; Vasaitis, R.; Menkis, A. Fungi associated with Ips acuminatus (Coleoptera: Curculionidae) in Ukraine with a special emphasis on pathogenicity of ophiostomatoid species. Eur. J. Entomol. 2017, 114, 77–85. [Google Scholar] [CrossRef]
- Davydenko, K.; Vasaitis, R.; Meshkova, V.; Menkis, A. Fungi associated with the red-haired bark beetle, Hylurgus ligniperda (Coleoptera: Curculionidae) in the forest-steppe zone in eastern Ukraine. Eur. J. Entomol. 2014, 111, 561–565. [Google Scholar] [CrossRef] [Green Version]
- Munoz-Adalia, E.J.; Sanz-Ros, A.V.; Flores-Pacheco, J.A.; Hantula, J.; Diez, J.J.; Vainio, E.J.; Fernandez, M. Sydowia polyspora dominates fungal communities carried by two Tomicus species in pine plantations threatened by Fusarium circinatum. Forests 2017, 8, 127. [Google Scholar] [CrossRef]
- Rosling, A.; Timling, I.; Taylor, D.L. Archaeorhizomycetes: patterns of distribution and abundance in soil. In Genomics of Soil- and Plant-Associated Fungi; Horwitz, B.A., Mukherjee, P.K., Mukherhee, M., Kubicek, C.P., Eds.; Springer Science & Business Media: Berlin, Germany, 2013; pp. 333–349. [Google Scholar]
- Baar, J.; Horton, T.R.; Kretzer, A.M.; Bruns, T.D. Mycorrhizal colonization of Pinus muricata from resistant propagules after a stand-replacing wildfire. New Phytol. 1999, 143, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Menkis, A.; Uotila, A.; Arhipova, N.; Vasaitis, R. Effects of stump and slash removal on growth and mycorrhization of Picea abies seedlings outplanted on a forest clear-cut. Mycorrhiza 2010, 20, 505–509. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenlid, J.; Finlay, R. Fungal communities in mycorrhizal roots of conifer seedlings in forest nurseries under different cultivation systems, assessed by morphotyping, direct sequencing and mycelial isolation. Mycorrhiza 2005, 16, 33–41. [Google Scholar] [CrossRef]
- Romon, P.; Hatting, H.; Goldarazena, A.; Iturrondobeitia, J.C. Variation in virulence of Beauveria bassiana and B. pseudobassiana to the pine weevil Pissodes nemorensis in relation to mycelium characteristics and virulence genes. Fungal Biol. 2017, 121, 189197. [Google Scholar] [CrossRef]
- Sánchez-Peña, S.R.; Lara, J.S.-J.; Medina, R.F. Occurrence of entomopathogenic fungi from agricultural and natural ecosystems in Saltillo, México, and their virulence towards thrips and whiteflies. J. Insect. Sci. 2011, 11, 1–10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand | Position | Age (y) | Mean Height (m) | Mean Diameter (cm) | Stocking Level | Forest Site Type * | Forest Vegetation Type ** | Tree Species Composition *** |
|---|---|---|---|---|---|---|---|---|
| Aukštaitija National Park (ANP) | ||||||||
| Managed | 55°13′26″ N, 25°50′49″ E | 37 | 14.8 | 18 | 0.7 | Nbl | vm | 60% P, 30% S, 10% B |
| 55°13′17″ N, 25°50′51″ E | 37 | 14.8 | 18 | 0.7 | Nbl | vm | 60% P, 30% S, 10% B | |
| 55°13′12″ N, 25°51′26″ E | 37 | 16 | 18 | 0.8 | Nbl | vm | 70% P, 40% S, 20% B | |
| 55°12′51″ N, 25°52′14″ E | 34 | 17.6 | 18 | 0.8 | Nbl | vm | 60% P, 20% S, 20% B | |
| 55°12′44″ N, 25°52′26″ E | 107 | 27.6 | 37 | 0.6 | Nbl | vm | 100% P | |
| Unmanaged | 55°13′12″ N, 25°51′08″ E | 27 | 18 | 19 | 0.6 | Nbl | vm | 70% P, 40% B, 20%S |
| 55°13′11″ N, 25°51′41″ E | 67 | 25.6 | 28 | 0.8 | Nbl | vm | 70% P, 30% S | |
| 55°12′55″ N, 25°51′59″ E | 77 | 25.2 | 28 | 0.7 | Nbl | vm | 80% P, 20% B | |
| 55°12′51″ N, 25°52′07″ E | 67 | 25.2 | 28 | 0.7 | Nbl | vm | 90% P, 10% B | |
| 55°12′50″ N, 25°52′22″ E | 34 | 15.2 | 16 | 0.8 | Nbl | vm | 50% P, 30% B, 20% S | |
| Dzūkija National Park (DNP) | ||||||||
| Managed | 54°04′36″ N, 24°19′16″ E | 106 | 24.5 | 37 | 0.7 | Nal | cl | 100% P |
| 54°04′11″ N, 24°19′34″ E | 56 | 19.5 | 20 | 0.9 | Nal | cl | 80% P, 20% B | |
| 54°04′10″ N, 24°20′30″ E | 96 | 25.6 | 35 | 0.7 | Nbl | vm | 100% P | |
| 54°04′08″ N, 24°20′56″ E | 111 | 25.4 | 35 | 0.7 | Nal | cl | 100% P | |
| 54°03′53″ N, 24°21′45″ E | 116 | 24.4 | 37 | 0.6 | Nal | cl | 100% P | |
| Unmanaged | 54°05′29″ N, 24°18′51″ E | 61 | 23.5 | 26 | 0.7 | Nal | cl | 100% P |
| 54°05′23″ N, 24°18′53″ E | 101 | 26.6 | 33 | 0.6 | Nal | cl | 100% P | |
| 54°05′11″ N, 24°18′58″ E | 36 | 11.8 | 14 | 0.9 | Nal | cl | 80% P, 20% B | |
| 54°04′58″ N, 24°18′58″ E | 51 | 19.7 | 20 | 0.8 | Nal | cl | 80% P, 20% B | |
| 54°04′47″ N, 24°19′04″ E | 81 | 24.9 | 32 | 0.8 | Nal | cl | 100% P | |
| Žemaitija National Park (ZNP) | ||||||||
| Managed | 56°02′09″ N, 21°54′45″ E | 23 | 8.8 | 9 | 0.7 | Ncl | ox | 50% P, 30% S, 20% B |
| 56°02′01″ N, 21°55′04″ E | 29 | 14 | 13 | 0.6 | Nbl | vm | 80% P, 20% S | |
| 56°02′11″ N, 21°55′23″ E | 57 | 23.5 | 25 | 0.9 | Ncl | ox | 100% P | |
| 56°02′35″ N, 21°56′29″ E | 67 | 27.3 | 29 | 0.7 | Ncl | ox | 40% P, 40% S, 20% B | |
| 56°02′42″ N, 21°57′00″ E | 32 | 18.9 | 19 | 0.8 | Ncl | ox | 50% P, 30% S, 20% B | |
| Unmanaged | 56°01′22″ N, 21°54′44″ E | 67 | 27.3 | 29 | 0.6 | Ncl | ox | 60% P, 30% S, 10% B |
| 56°01′35″ N, 21°54′20″ E | 52 | 20.6 | 25 | 0.6 | Ncl | ox | 40% P, 40% S, 20% B | |
| 56°01′24″ N, 21°54′07″ E | 52 | 19.6 | 21 | 0.7 | Ncl | ox | 60% P, 40% S | |
| 56°00′58″ N, 21°52′54″ E | 72 | 26.3 | 31 | 0.8 | Ncl | ox | 90% P, 10% B | |
| 56°00′48″ N, 21°52′18″ E | 132 | 27.1 | 36 | 1.0 | Pbn | csps | 40% P, 40%, 20% B | |
| Stand | Needles | Soil | ||||||
|---|---|---|---|---|---|---|---|---|
| No. of Needle Samples/Amplicon Pools | No. of Sequences | No. of Fungal Taxa | Shannon Diversity Index | No. of Soil Samples/Pools | No. of Sequences | No. of Fungal Taxa | Shannon Diversity Index | |
| Aukštaitija National Park (ANP) | ||||||||
| Managed | 5/1 | 1708 | 202 | 4.2 | 3/1 | 486 | 77 | 3.2 |
| 5/1 | 3289 | 284 | 3.9 | 3/1 | 1875 | 155 | 3.5 | |
| 5/1 | 1668 | 183 | 3.8 | 3/1 | 1269 | 160 | 4.0 | |
| 5/1 | 4385 | 318 | 4.3 | 3/1 | 2 | 2 | 0.7 | |
| 5/1 | 3496 | 286 | 3.8 | 3/1 | 2637 | 144 | 3.5 | |
| All Managed | 25/5 | 14,546 | 598 | 15/5 | 6269 | 312 | ||
| Unmanaged | 5/1 | 5819 | 378 | 4.4 | 3/1 | 2866 | 200 | 3.7 |
| 5/1 | 3434 | 196 | 3.0 | 3/1 | 430 | 114 | 3.9 | |
| 5/1 | 1697 | 170 | 3.5 | 3/1 | 718 | 140 | 3.9 | |
| 5/1 | 209 | 71 | 3.4 | 3/1 | 921 | 60 | 2.6 | |
| 5/1 | 4106 | 292 | 4.1 | 3/1 | 1919 | 133 | 3.4 | |
| All Unmanaged | 25/5 | 15,265 | 595 | 15/5 | 6854 | 399 | ||
| All ANP | 50/10 | 29,811 | 799 | 30/10 | 13,123 | 525 | ||
| Dzūkija National Park (DNP) | ||||||||
| Managed | 5/1 | 5811 | 240 | 3.3 | 3/1 | 1138 | 199 | 4.1 |
| 5/1 | 4798 | 276 | 3.6 | 3/1 | 2636 | 158 | 3.7 | |
| 5/1 | 8802 | 341 | 3.6 | 3/1 | 4104 | 199 | 2.9 | |
| 5/1 | 2460 | 174 | 3.3 | 3/1 | 1464 | 216 | 4.3 | |
| 5/1 | 1536 | 134 | 3.2 | 3/1 | 681 | 165 | 4.3 | |
| All Managed | 25/5 | 23,407 | 553 | 15/5 | 10,023 | 519 | ||
| Unmanaged | 5/1 | 1388 | 135 | 3.6 | 3/1 | 306 | 90 | 3.7 |
| 5/1 | 2963 | 194 | 2.9 | 3/1 | 2176 | 119 | 2.9 | |
| 5/1 | 3043 | 207 | 3.4 | 3/1 | 1672 | 118 | 3.1 | |
| 5/1 | 1638 | 141 | 3.3 | 3/1 | 1841 | 131 | 3.4 | |
| 5/1 | 5301 | 247 | 3.5 | 3/1 | 2048 | 99 | 2.9 | |
| All Unmanaged | 25/5 | 14,333 | 434 | 15/5 | 8043 | 308 | ||
| All DNP | 50/10 | 37,740 | 662 | 30/10 | 18,066 | 616 | ||
| Žemaitija National Park (ZNP) | ||||||||
| Managed | 5/1 | 4977 | 259 | 3.6 | 3/1 | 1783 | 174 | 3.8 |
| 5/1 | 988 | 183 | 4.2 | 3/1 | 4366 | 151 | 2.3 | |
| 5/1 | 666 | 106 | 3.4 | 3/1 | 1256 | 120 | 2.3 | |
| 5/1 | 6269 | 339 | 3.7 | 3/1 | 210 | 98 | 4.2 | |
| 5/1 | 3731 | 267 | 3.7 | 3/1 | 966 | 161 | 4.2 | |
| All Managed | 25/5 | 16,631 | 638 | 15/5 | 8581 | 450 | ||
| Unmanaged | 5/1 | 4058 | 278 | 2.8 | 3/1 | 2425 | 151 | 2.5 |
| 5/1 | 5214 | 313 | 4.1 | 3/1 | 1050 | 173 | 4.2 | |
| 5/1 | 5751 | 135 | 1.5 | 3/1 | 887 | 159 | 4.2 | |
| 5/1 | 7409 | 208 | 2.9 | 3/1 | 2,952 | 212 | 3.9 | |
| 5/1 | 8240 | 229 | 2.9 | 3/1 | 2,934 | 151 | 2.9 | |
| All Unmanaged | 25/5 | 30,672 | 648 | 15/5 | 10,248 | 517 | ||
| All ZNP | 50/10 | 47,303 | 870 | 30/10 | 18,829 | 741 | ||
| All | 150/30 | 114,854 | 1261 | 90/30 | 50,018 | 1186 | ||
| Taxon | Phylum | Class | GenBank Reference | Similarity, % * | Managed, % | Unmanaged, % | All, % |
|---|---|---|---|---|---|---|---|
| Coleosporium campanulae | Basidiomycota | Pucciniomycetes | KY810468 | 322/322 (100%) | 4.1 | 20.0 | 12.4 |
| Unidentified sp. 3980_1 | Ascomycota | Dothideomycetes | KP897304 | 244/244 (100%) | 14.2 | 10.8 | 12.4 |
| Unidentified sp. 3980_4 | Ascomycota | Dothideomycetes | KP891553 | 259/259 (100%) | 3.8 | 4.5 | 4.1 |
| Sydowia polyspora | Ascomycota | Dothideomycetes | MG888613 | 256/256 (100%) | 3.1 | 3.0 | 3.1 |
| Unidentified sp. 3980_13 | Ascomycota | Eurotiomycetes | MG827663 | 262/262 (100%) | 3.8 | 2.1 | 2.9 |
| Unidentified sp. 3980_3 | Basidiomycota | Tremellomycetes | KU687386 | 302/307 (98%) | 3.1 | 2.5 | 2.8 |
| Phaeococcomyces eucalypti | Ascomycota | Eurotiomycetes | NR_120226 | 246/248 (99%) | 2.6 | 2.0 | 2.3 |
| Unidentified sp. 3980_10 | Ascomycota | Dothideomycetes | MG827778 | 258/258 (100%) | 2.8 | 1.8 | 2.3 |
| Microsphaeropsis olivacea | Ascomycota | Dothideomycetes | MH871969 | 249/249 (100%) | 2.1 | 1.9 | 2.0 |
| Epithamnolia xanthoriae | Ascomycota | Leotiomycetes | KY814539 | 234/238 (98%) | 2.4 | 1.6 | 2.0 |
| Cladosporium cladosporioides | Ascomycota | Dothideomycetes | MH042811 | 243/243 (100%) | 2.1 | 1.5 | 1.8 |
| Unidentified sp. 3980_33 | Ascomycota | Leotiomycetes | KP897394 | 223/258 (86%) | 2.0 | 1.1 | 1.6 |
| Unidentified sp. 3980_37 | Ascomycota | Dothideomycetes | KP897394 | 236/259 (91%) | 2.5 | 0.6 | 1.5 |
| Curvibasidium cygneicollum | Basidiomycota | Microbotryomycetes | KY102972 | 310/310 (100%) | 1.1 | 1.7 | 1.4 |
| Unidentified sp. 3980_30 | Ascomycota | Leotiomycetes | KY742593 | 242/242 (100%) | 1.9 | 0.9 | 1.4 |
| Vishniacozyma victoriae | Basidiomycota | Tremellomycetes | LC085209 | 234/234 (100%) | 0.6 | 2.0 | 1.3 |
| Unidentified sp. 3980_25 | Ascomycota | Eurotiomycetes | KP891398 | 255/255 (100%) | 1.6 | 1.0 | 1.3 |
| Seimatosporium lichenicola | Ascomycota | Sordariomycetes | JF320818 | 247/248 (99%) | 2.4 | 0.2 | 1.2 |
| Heterotruncatella spartii | Ascomycota | Sordariomycetes | MK012418 | 245/245 (100%) | 1.4 | 0.9 | 1.1 |
| Lophodermium pinastri | Ascomycota | Leotiomycetes | MH856647 | 239/239 (100%) | 1.6 | 0.6 | 1.1 |
| Total of 20 taxa | 59.3 | 60.6 | 60.0 |
| Taxon | Phylum | Class | GenBank Reference | Similarity, % * | Managed, % | Unmanaged, % | All, % |
|---|---|---|---|---|---|---|---|
| Unidentified sp. 3980_21 | Ascomycota | Archaeorhizomycetes | KC965182 | 219/219 (100%) | 12.1 | 5.2 | 8.6 |
| Umbelopsis nana | Mucoromycota | Mucoromycotina | MH857049 | 293/293 (100%) | 7.4 | 8.9 | 8.2 |
| Archaeorhizomyces sp. 3980_5 | Ascomycota | Archaeorhizomycetes | MH248043 | 207/207 (100%) | 10.1 | 6.1 | 8.1 |
| Penicillium spinulosum | Ascomycota | Eurotiomycetes | MK131675 | 251/251 (100%) | 3.8 | 8.7 | 6.3 |
| Oidiodendron chlamydosporicum | Ascomycota | Dothideomycetes | MG597466 | 235/235 (100%) | 4.4 | 6.0 | 5.2 |
| Oidiodendron echinulatum | Ascomycota | Dothideomycetes | MG597467 | 236/236 (100%) | 2.0 | 2.8 | 2.4 |
| Sagenomella verticillata | Ascomycota | Eurotiomycetes | MH860215 | 263/263 (100%) | 2.8 | 1.7 | 2.3 |
| Unidentified sp. 3980_28 | Mucoromycota | Mucoromycotina | HQ022209 | 299/299 (100%) | 0.9 | 3.6 | 2.2 |
| Tolypocladium geodes | Ascomycota | Sordariomycetes | MH864065 | 248/248 (100%) | 0.5 | 3.6 | 2.0 |
| Pseudogymnoascus roseus | Ascomycota | Dothideomycetes | MH865208 | 241/241 (100%) | 1.8 | 1.5 | 1.7 |
| Meliniomyces bicolor | Ascomycota | Leotiomycetes | MG597461 | 237/238 (99%) | 1.7 | 1.5 | 1.6 |
| Cladosporium cladosporioides | Ascomycota | Dothideomycetes | MH042811 | 243/243 (100%) | 1.5 | 1.5 | 1.5 |
| Wilcoxina rehmii | Ascomycota | Pezizomycetes | MF926519 | 253/254 (99%) | 2.5 | 0.03 | 1.2 |
| Unidentified sp. 3980_60 | Basidiomycota | Microbotryomycetes | HQ021811 | 320/320 (100%) | 1.4 | 0.7 | 1.1 |
| Unidentified sp. 3980_64 | Ascomycota | Sordariomycetes | KJ826970 | 317/322 (98%) | 1.2 | 0.9 | 1.0 |
| Malassezia restricta | Basidiomycota | Ustilaginomycotina | CP030254 | 368/369 (99%) | 0.9 | 1.1 | 1.0 |
| Pseudeurotium sp. 3980_68 | Ascomycota | Dothideomycetes | MF692976 | 229/241 (95%) | 1.8 | 0.2 | 1.0 |
| Hyphodiscus sp. 3980_71 | Ascomycota | Leotiomycetes | NR_155151 | 235/243 (97%) | 0.2 | 1.7 | 0.9 |
| Aspergillus cervinus | Ascomycota | Eurotiomycetes | MH865525 | 262/262 (100%) | 0.8 | 1.0 | 0.9 |
| Beauveria pseudobassiana | Ascomycota | Sordariomycetes | MF872419 | 255/255 (100%) | 0.2 | 1.4 | 0.8 |
| Total of 20 taxa | 58.2 | 58.0 | 58.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynikienė, J.; Marčiulynienė, D.; Marčiulynas, A.; Gedminas, A.; Vaičiukynė, M.; Menkis, A. Managed and Unmanaged Pinus sylvestris Forest Stands Harbour Similar Diversity and Composition of the Phyllosphere and Soil Fungi. Microorganisms 2020, 8, 259. https://doi.org/10.3390/microorganisms8020259
Lynikienė J, Marčiulynienė D, Marčiulynas A, Gedminas A, Vaičiukynė M, Menkis A. Managed and Unmanaged Pinus sylvestris Forest Stands Harbour Similar Diversity and Composition of the Phyllosphere and Soil Fungi. Microorganisms. 2020; 8(2):259. https://doi.org/10.3390/microorganisms8020259
Chicago/Turabian StyleLynikienė, Jūratė, Diana Marčiulynienė, Adas Marčiulynas, Artūras Gedminas, Miglė Vaičiukynė, and Audrius Menkis. 2020. "Managed and Unmanaged Pinus sylvestris Forest Stands Harbour Similar Diversity and Composition of the Phyllosphere and Soil Fungi" Microorganisms 8, no. 2: 259. https://doi.org/10.3390/microorganisms8020259